中国荷斯坦育肥公犊肉储藏过程中脂肪氧化对肌红蛋白稳定性的影响
2015-05-08吴成帆于春起余群力李洪波
吴成帆,韩 玲,*,陈 骋,于春起,余群力,李洪波
(1.甘肃农业大学食品科学与工程学院,甘肃兰州 730070;2.河北福成五丰食品股份有限公司,河北三河 065200;3.甘肃陇原中天生物工程股份有限公司,甘肃陇西 748100)
中国荷斯坦育肥公犊肉储藏过程中脂肪氧化对肌红蛋白稳定性的影响
吴成帆1,韩 玲1,*,陈 骋1,于春起2,余群力1,李洪波3
(1.甘肃农业大学食品科学与工程学院,甘肃兰州 730070;2.河北福成五丰食品股份有限公司,河北三河 065200;3.甘肃陇原中天生物工程股份有限公司,甘肃陇西 748100)
本文通过参考国内外研究方法,测定中国荷斯坦育肥公犊和普通黄牛肉脂肪酸组成及其在有氧贮藏过程中脂肪和肌红蛋白的氧化程度,研究其脂肪氧化和肌红蛋白氧化之间的关系。结果表明,育肥公犊的饱和脂肪酸含量极显著高于黄牛(p<0.01),多不饱和脂肪酸和单不饱和脂肪酸含量都极显著低于黄牛(p<0.01)。在贮藏期间,两种牛都发生脂质氧化,且育肥公犊的氧化程度极显著低于黄牛(p<0.01)。在有氧贮藏条件下,两种牛的肌红蛋白氧化程度增加。育肥公犊的高铁肌红蛋白含量与脂质氧化程度呈线性正相关,(r=0.945,p<0.01),氧合肌红蛋白含量与脂质氧化程度呈线性负相关,(r=-0.906,p<0.01)。黄牛的高铁肌红蛋白含量与脂质氧化程度呈线性正相关,(r=0.972,p<0.01),氧合肌红蛋白含量与脂质氧化程度呈线性负相关,(r=-0.851,p<0.05)。通过研究表明中国荷斯坦育肥公犊肉脂质氧化与肉色稳定性相关。
背最长肌,脂肪氧化,高铁肌红蛋白,氧合肌红蛋白
肉色是评价牛肉品质的一项主要标准,鲜肉的货架期较短是肉类企业亟待克服的问题[1]。消费者大多都通过肉色来决定是否购买,良好、正常的肉色可以刺激消费者的购买欲,给商家带来利益,而一旦肉色劣变则会很难被卖出,给商家带来经济损失[2]。
肉色主要是由肉中的两种色素物质肌红蛋白和血红蛋白决定的,在放血充分的条件下肉色主要是由肌红蛋白的化学性质、数量和分布决定的[3]。肌红蛋白本身是紫红色,它的铁离子是二价的,且第六位缺乏配体键结合,当二价铁离子与氧气结合后,肌红蛋白成为氧合肌红蛋白,呈鲜红色,铁离子仍是二价的[4-5]。继续氧化一段时间后二价的铁离子会变成三价的,生成高铁肌红蛋白,呈褐色[6]。
研究发现脂类氧化产生的各种小分子次级代谢产物,比如醛会催化肌红蛋白氧化,生成高铁肌红蛋白,4-羟基壬烯醛与肌红蛋白共价结合,使肌红蛋白更易被氧化成高铁肌红蛋白从而失去运输氧气的能力[7-8]。在肌肉微粒体实验中,不饱和脂肪酸的含量越高,肌红蛋白氧化速率越快[9-10]。饲喂动物含有丰富的不饱和脂肪酸的饲料,其肌红蛋白氧化速率也显著加快。因此,体内脂肪酸成分的差异及脂类氧化的程度可影响肌红蛋白氧化[11-12]。
但已有研究的不足表现在对于肉制品中脂肪氧化促进色素氧化的机理目前还没有系统的研究报道,当前研究尚不能提供良好的方法来控制肉色的氧化。中国荷斯坦育肥公犊是一种优良的牛肉品种,对其研究还不足,因此,本研究以中国荷斯坦育肥公犊的背最长肌作为研究对象,普通黄牛的背最长肌作为对照,参考国内外的研究方法,测定其脂肪酸组成及其在有氧贮藏过程中脂肪和肌红蛋白的氧化程度,研究其脂肪氧化和肌红蛋白氧化之间的关系,为进一步探索牛肉脂肪氧化对肉色稳定性的影响提供科学依据。
1 材料与方法
1.1 材料与仪器
盐酸;乙醇 成都市科龙化工试剂厂、乙醚 杭州新方五交化有限公司;石油醚 天津市津东天正精细化学试剂厂;氢氧化钠 天津市凯通化学试剂有限公司;甲醇 南京化学试剂有限公司;正庚烷 深圳市科天化玻仪器有限公司;无水硫酸钠 天津市光复科技发展有限公司;三氯乙酸 天津市光复精细化工研究所;硫代巴比妥酸 上海科丰化学试剂有限公司;氯仿 上海研臣实业有限公司;1,1,3,3,-四乙氧基丙烷 上海研臣实业有限公司;磷酸二氢钠 天津市光复精细化工研究所;磷酸氢二钠 天津市光复科技发展有限公司。
美国安捷伦7890A 型气相色谱仪 上海科晓科学仪器有限公司;TGL-24MC台式高速冷冻离心机 长沙湘仪离心机仪器有限公司;SP-756P型紫外可见分光光度计 上海光谱仪器有限公司;、JY92-ⅡDN超声波细胞粉碎机 宁波新芝生物科技股份有限公司。
1.2 实验方法
1.2.1 样品采集 在河北福成五丰食品股份有限公司分别采集5头12月龄的生长发育良好的中国荷斯坦育肥公犊和相同条件下当地普通黄牛的背最长肌,除去结缔组织,垂直肌纤维方向切分成50g肉块,装入聚乙烯收缩薄膜托盘,储存在(4±0.5)℃避光的冰箱中,分别在宰后1、2、3、4、5、6、7d进行相应指标的测定。
1.2.2 脂肪酸测定
1.2.2.1 脂肪的提取 准确称取育肥公犊和黄牛背最长肌各10g,在研钵内研细,加10mL HCl和10mL蒸馏水,置于80℃水浴中加热至完全溶解,然后加20mL 乙醇冷却。在具塞试管中加入25mL乙醚浸提摇匀1min,然后放气,用石油醚-乙醚(各取10mL)冲洗塞子及壁口附着的脂肪,静置15min后用10mL乙醚浸提,上清液水浴蒸干后置于烘箱干燥。
1.2.2.2 脂肪酸甲酯化 脂肪酸甲酯化:参照GB/T 17376-2008。将0.5mL 氢氧化钠-甲醇标准液和40mL 甲醇加入圆底烧瓶,在(60~70)℃下回流10min,待冷却后加20mL正庚烷洗涤,转入分液漏斗,并加40mL蒸馏水震荡、静置分层,上清液pH为7.0时,用适量无水硫酸钠过滤后用有机相过滤膜过滤,密封避光冷藏。甲酯化的脂肪酸待测样品用美国安捷伦7890A型气相色谱仪测定[13-14]。
1.2.3 脂类氧化测定 参考Sinnhuber的方法[15]:准确称取肉样2g,置于250mL具塞三角瓶内,加入10mL 7.5%的三氯乙酸溶液,振摇30min,用双层滤纸过滤,重复用双层滤纸过滤一次,准确移取上述滤液5mL置于25mL比色管内,加入5mL TBA(0.02mol/L),混匀,加塞,置于90℃水浴锅内,保温40min,取出冷却1h,移入小试管内离心5min(1600r/min),上清液倒入25mL比色管内,加入5mL氯仿,摇匀,静置,分层,吸出上清液分别在532nm和600nm波长处比色(同时做空白实验),记录吸光值。按照标准曲线将TBA值换算成丙二醛含量。
标准曲线的制备:分别取1,1,3,3-四乙氧基丙烷(TEP)应用液(相当于10μg丙二醛/mL)0.0、0.2、0.4、0.6、0.8、1.0mL,置于25mL闭塞管内,加水至总体积5mL,加入5mL TBA溶液,与样品做同样处理,最后比色,以OD值为横坐标,以TEP溶液浓度为纵坐标做曲线。
1.2.4 肌红蛋白氧化状态测定 参考Krzywicki的方法[16],取肉样20g,加入20mL浓度为0.04 mol/L、pH6.8的磷酸钠缓冲液,室温下匀浆25s,10800r/min。置匀浆液于冰浴中放置1h,后于1000×g、10~15℃下离心30min。上清经滤纸过滤,用同样的缓冲液补足至25mL,测定其在525、545、565、572nm处的吸光度值。
不同氧化状态肌红蛋白相对含量的计算方法:
总肌红蛋白(TMb)含量(mg·g-1)=-0.166A572+0.086A565+0.088A545+0.099A525
氧合肌红蛋白(OMb)百分含量(%)=(0.882R1-1.267R2+0.809R3-0.361)×100
高铁肌红蛋白(MMb)百分含量(%)=(-2.514R1+0.777R2+0.800R3+1.098)×100
式中:R1、R2、R3分别为A572/A525、A565/A525、A545/A525。
1.3 统计分析
分析结果采用Microsoft Excel进行处理,并用SPSS19.0软件对两种牛高铁肌红蛋白和氧合肌红蛋白氧化程度与脂质氧化程度的关系进行相关性分析。
2 结果与分析
2.1 脂肪酸组成
由表1 可以看出,育肥公犊肉含10种饱和脂肪酸(SFA),黄牛肉含6种,饱和脂肪酸含量育肥公犊肉(52.54%)极显著高于黄牛肉(48.53%)(p<0.01);育肥公犊肉不饱和脂肪酸总量(35.93%)极显著低于黄牛(42.25%)(p<0.01)。其中,育肥公犊肉含3种单不饱和脂肪酸(MUFU),黄牛肉含2种,单不饱和脂肪酸含量育肥公犊肉(30.67%)极显著低于黄牛肉(35.50%)(p<0.01);育肥公犊肉含5种多不饱和脂肪酸(PUFA),黄牛肉含3种,多不饱和脂肪酸含量育肥公犊肉(5.26%)极显著低于黄牛肉(6.75%)(p<0.01)。

表1 中国荷斯坦育肥公犊和黄牛的脂肪酸形式和组成
注:SFA:饱和脂肪酸;MUFA:单不饱和脂肪酸;PUFA:多不饱和脂肪酸;N:未检测出;表中同行间*代表差异显著(p<0.05),**代表差异极显著(p<0.01)。
2.2 肌肉脂质氧化
由TBA标准曲线得计算公式:TBA(mg/kg)=13.51(A532-A600)-0.061
式中:A532表示532nm波长处的吸光度值;A600表示600nm波长处的吸光度值。
由图1可以看出,在宰后第1d黄牛的脂质氧化程度与育肥公犊相似,随着储藏时间的延长,黄牛和育肥公犊的脂质氧化程度都增加,但黄牛的脂质氧化速率高于育肥公犊,在第7d黄牛的脂质氧化程度明显的高于育肥公犊。贮藏过程中,黄牛脂肪氧化程度极显著高于育肥公犊(p<0.01)。

图1 中国荷斯坦育肥公犊和黄牛在贮藏期间脂肪氧化Fig.1 Time course of lipid oxidation in fattened Chinese Holstein male calves and Yellow Cattle muscle
霍晓娜等[17]研究发现,不饱和脂肪酸含量高的猪腿肉肌间组织中的脂类氧化速率显著高于不饱和脂肪酸含量低的猪腿肉皮下组织,可能是由两者不饱和脂肪酸尤其是多不饱和脂肪酸含量差异所致。不饱和脂肪酸含量和不饱和程度越高,脂类氧化越易进行。这与本实验研究结果一致。
2.3 肌红蛋白氧化状态
从图2可以看出,在宰后第1d黄牛背最长肌中高铁肌红蛋白含量与育肥公犊差异不明显,并且在贮藏的前3d两种牛的高铁肌红蛋白含量变化都不明显,但从第3d开始,两种牛背最长肌中高铁肌红蛋白含量都显著升高,黄牛的高铁肌红蛋白含量上升趋势明显快于育肥公犊肉,在第7d,黄牛背最长肌中高铁肌红蛋白含量达到51.02%,高于育肥公犊,贮藏过程中,黄牛高铁肌红蛋白含量极显著高于育肥公犊(p<0.01)。
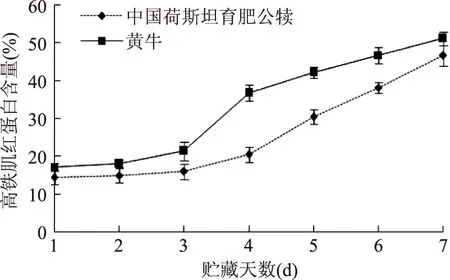
图2 中国荷斯坦育肥公犊和黄牛在贮藏期间高铁肌红蛋白含量变化Fig.2 Time course of metmyoglobin oxidation in fattened Chinese Holstein male calves and Yellow cattle muscle
高铁肌红蛋白是牛肉肌纤维中肌红蛋白在低氧的情况下Fe2+转变为Fe3+所形成,高铁肌红蛋白含量的增加也是牛肉氧化程度加深的表现。在有氧状态下造成鲜肉腐败的主要微生物是好氧的假单胞菌,当其在对数生长期时,好氧量增大,从而使肉表面局部氧分压降低,促进棕褐色的高铁肌红蛋白形成[18]。
孙烨琨[8]和Hutchins[19]的研究报告表明无论是在牦牛还是黄牛中,其肌红蛋白和脂类氧化的趋势相近,呈显著正相关,这与本实验的研究结果一致。Lee等[20]报道称脂肪酸氧化的次级代谢产物,尤其是不饱和产物可能会使得氧合肌红蛋白血红素基团暴露而更易氧化,产生的自由基加剧高铁肌红蛋白的积累。
从图3可以看出,在贮藏第1d育肥公犊的氧合肌红蛋白含量略高于黄牛,贮藏前3d两种牛的氧合肌红蛋白含量都略有上升,这可能是在最初有氧储藏条件下高铁肌红蛋白与氧气结合,生成了氧合肌红蛋白,导致其含量增加。从第3d到第7d两种牛的氧合肌红蛋白含量都显著下降,这可能是因为随着贮藏时间的延长,高铁肌红蛋白生成量增加,导致氧合肌红蛋白含量减少。黄牛的下降速率高于育肥公犊,在第7d育肥公犊的氧合肌红蛋白含量显著高于黄牛。贮藏过程中,黄牛氧合肌红蛋白含量极显著低于育肥公犊(p<0.01)。

图3 中国荷斯坦育肥公犊和黄牛在贮藏期间氧合肌红蛋白含量变化Fig.3 Time course of oxymyoglobin oxidation in fattened Chinese Holstein male calves and Yellow cattle muscle
孙京新等[21]报道称脂肪酸氧化的不饱和产物可能会导致氧合肌红蛋白构象的改变,使得氧合肌红蛋白血红素基团暴露而氧化,从而使得氧合肌红蛋白含量不断减少,这与高铁肌红蛋白变化趋势相互补。
2.4 肌肉脂肪氧化与肌红蛋白稳定性的关系
从表2中可以看出,两种牛的高铁肌红蛋白含量与脂质氧化程度(TBArs)呈极显著正相关。经过对高铁肌红蛋白含量和TBArs相关性的分析可以得出,随着脂质氧化程度的上升,高铁肌红蛋白含量增加,这就使得肉的色泽不断向褐色变化,影响其销售。育肥公犊的氧合肌红蛋白含量与脂质氧化程度(TBArs)呈极显著负相关,黄牛的氧合肌红蛋白含量与脂质氧化程度(TBArs)呈显著负相关。随着脂质氧化程度的不断上升,氧合肌红蛋白不断被氧化成高铁肌红蛋白,含量减少,使得肉色由鲜红色变为褐色。

表2 肌肉脂肪氧化与肌红蛋白含量相关性分析
注:表中同行间*代表差异显著(p<0.05),**代表差异极显著(p<0.01)。
脂肪氧化过程中产生的自由基可破坏肉中的色素,使肉变色而牛肉变色后产生的Fe3+又是脂肪氧化的催化剂[22]。脂肪氧化和牛肉的褪色之间存在着密切的关系,脂肪氧化与肉色紧密相关,脂肪氧化产生的自由基促进高铁肌红蛋白的积累,游离脂肪酸及肌肉磷脂会加速氧合肌红蛋白的自动氧化,脂肪氧化越剧烈,高铁肌红蛋白的积累越多,致使牛肉的颜色逐渐转变为褐色[23]。
3 结论
3.1 育肥公犊的饱和脂肪酸含量极显著高于黄牛(p<0.01),单不饱和脂肪酸和多不饱和脂肪酸含量都极显著低于黄牛(p<0.01)。
3.2 随着贮藏时间的延长,育肥公犊和普通黄牛都发生脂质氧化,育肥公犊的脂质氧化程度极显著低于黄牛(p<0.01)。
3.3 在贮藏期间,高铁肌红蛋白含量的变化趋势与脂质氧化变化趋势一致,氧合肌红蛋白含量的变化趋势与脂质氧化变化趋势相互补。
3.4 在有氧贮藏条件下,中国荷斯坦育肥公犊肉高铁肌红蛋白含量与脂质氧化程度呈正相关,且差异极显著(p<0.01),氧合肌红蛋白含量与脂质氧化程度呈负相关,且差异极显著(p<0.01),黄牛的高铁肌红蛋白含量与脂质氧化程度呈正相关,且差异极显著(p<0.01),氧合肌红蛋白含量与脂质氧化程度呈负相关,且差异显著(p<0.05)。
[1]Jun Lee B,G Hendricks D,P Cornforth D. A comparison of carnosine and ascorbic acid on color and lipid stability in a ground beef pattie model system[J]. Meat Science,1999,51(3):245-253.
[2]Cornforth D. Color-its basis and importance[M]. Quality attributes and their measurement in meat,poultry and fish products. Springer US,1994:34-78.
[3]Faustman C,Cassens R G. The biochemical basis for discoloration in fresh meat:A review[J]. Journal of Muscle Foods,1990,1(3):217-243.
[4]Clydesdale F M. Colorimetry-methodology and applications[J]. Critical Reviews in Food Science and Nutrition,1978,10(3):243-302.
[5]阚建全,食品化学[M].北京:中国农业大学出版社,第2版,2008:300-303.
[6]唐孟甜,谢丽,戴瑞彤. 脂肪氧化产物丙二醛对牛肉色泽稳定性的影响[J]. 农产品加工学刊,2011,08:4-8
[7]Faustman C,Liebler D C,McClure T D,et al. α,β-Unsaturated aldehydes accelerate oxymyoglobin oxidation[J]. Journal of agricultural and food chemistry,1999,47(8):3140-3144.
[8]孙烨琨,王淼,杨虎,等. 牦牛肉脂肪酸组成与肌红蛋白氧化的关系[J]. 食品科学,2010,31(21):62-64.
[9]Alderton A L,Faustman C,Liebler D C,et al. Induction of redox instability of bovine myoglobin by adduction with 4-hydroxy-2-nonenal[J]. Biochemistry,2003,42(15):4398-4405.
[10]Yin M,Faustman C. The influence of microsomal and cytosolic components on the oxidation of myoglobin and lipidinvitro[J]. Food Chemistry,1994,51(2):159-164.
[11]Nute G R,Richardson R I,Wood J D,et al. Effect of dietary oil source on the flavour and the colour and lipid stability of lamb meat[J]. Meat science,2007,77(4):547-555.
[12]Martin D,Ruiz J,Kivikari R,et al. Partial replacement of pork fat by conjugated linoleic acid and/or olive oil in liver patés:Effect on physicochemical characteristics and oxidative stability[J]. Meat science,2008,80(2):496-504.
[13]Wood JD,Enser M,Fisher AV,et al. Fat deposition,fatty acid composition and meat quality:a review[J]. Meat Sci,2008,78(4):343-358.
[14]李儒仁,余群力,韩玲,等.日本和牛与秦川牛杂交牛肝脏营养特性分析[J].营养学报,2013,35(5):502-504.
[15]Sinnhuber R O,Yu T C. The 2-thiobarbituric acid reaction,an objective measure of the oxidative deterioration occurring in fats and oils[J]. 油化学,1977,26(5):259-267.
[16]Krzywicki,K. The determinate of haem pigments in meat. Meat Science 1982,7:29-36.
[17]霍晓娜,李兴民,刘毅,等. 猪腿肉脂肪酸组成及脂肪氧化的研究[J]. 食品科学,2006,27(1):101-104.
[18]Hoffman L C,Ferreria A V. pH decline of the M. longissimus thoracis of night-cropped Grey Duiker(Sylvicapra grimmia)[J]. South African Journal of Animal Science,2000,30(1):16-17.
[19]Hutchins B K,Liu T H P,Watts B M. Effect of additives and refrigeration on reducing activity,metmyoglobin and malonaldehyde of raw ground beef[J]. Journal of Food Science,1967,32(2):214-217.
[20]Lee S,Phillips A L,Liebler D C,et al. Porcine oxymyoglobin and lipid oxidationinvitro[J]. Meat Science,2003,63(2):241-247.
[21]孙京新,周光宏,徐幸莲. 猪肉微粒体脂肪酸氧化产物对氧合肌红蛋白氧化的影响[J]. 南京农业大学学报,2004,27(1):101-104.
[22]Gebert S,Eichenberger B,Pfirter H P,et al. Influence of different dietary vitamin C levels on vitamin E and C content and oxidative stability in various tissues and stored m. longissimus dorsi of growing pigs[J]. Meat science,2006,73(2):362-367.
[23]梁成云,唐丹,展凤军,等. 不同年龄延黄牛肉在贮藏期间肉色与氧化稳定性的研究[J]. 食品科技,2009(7):99-102.
Effect of lipid oxidation on the stability of myoglobin during storage of fattened Chinese Holstein male calves meat
WU Cheng-fan1,HAN Ling1,*,CHEN Cheng1,YU Chun-qi2,YU Qun-li1,LI Hong-bo3
(1.Food Science and Engineering Department of Gansu Agricultural University,Lanzhou 730070,China;2.Hebei Fortune Ng Fung Food Co.,Ltd.,Sanhe 065200,China;3.Gansu Longyuan Zhongtian Biological Engineering Co.,Ltd.,Longxi 748100,China)
In this paper,the fatty acids composition of the fattened Chinese Holstein male calves and yellow cattle were investigated. Then,the relationship between lipid oxidation and myoglobin oxidation were studied during the aerobic storage by using the domestic and foreign research methods. The result showed that the contents of saturated fatty acids(SFA)of Holstein calves was extremely significant higher than that of yellow cattle(p<0.01),the content of polyunsaturated fatty acids(PUFA)and monounsaturated fatty acids(MSFA)were extremely significant lower(p<0.01). Lipid oxidation and myoglobin oxidation happened in both of them under the aerobic storage conditions,the oxidation degree of Holstein calves was extremely significant lower than that of yellow cattle(p<0.01). For Holstein calves,Metmyoglobin content was positively related to lipid oxidation(r=0.945,p<0.01),Oxymyoglobin content was negatively related to lipid oxidation(r=-0.906,p<0.01).For yellow cattle,Metmyoglobin content was positively related to lipid oxidation(r=0.972,p<0.01),Oxymyoglobin content was negatively related to lipid oxidation(r=-0.851,p<0.05). In a conclusion,the lipid oxidation of fattened Chinese Holstein male calves meat was associated with color stability.
longissimus muscle;lipid oxidation;metmyoglobin;oxymyoglobin
2014-05-15
吴成帆(1989-),女,硕士研究生,研究方向:食品科学。
*通讯作者:韩玲(1963-),女,博士,教授,主要从事畜产品加工及贮藏研究。
国家现代农业(肉牛牦牛)产业技术体系资助(CARS-38);国家自然科学基金(31260380);国家公益性行业(农业)科研专项经费资助项目(201203009)。
TS252.1
A
1002-0306(2015)07-0323-05
10.13386/j.issn1002-0306.2015.07.059
