广西乡土用材树种米老排资源分布
2015-04-29刘光金张培贾宏炎蔡道雄农志劳庆祥
刘光金 张培 贾宏炎 蔡道雄 农志 劳庆祥
摘要[目的]保护和利用优异米老排种质资源。[方法]开展米老排种质资源分布调查,收集82株优势木,并对其种实特性进行了分析。[结果]米老排分布区域狭窄,零星分布于龙州、靖西、德保、那坡、东兴、上思、容县、凭祥8个县,集中分布在21°32′~23°11′N,105°51′~110°34′E,海拔179~1 202 m之间的低中山区。采种优势木胸径与树高之间呈现极显著的正相关性,相关系数为0.526,S型曲线拟合度最高,相关系数为0.749。米老排结实量大,果实为蒴果,10月中旬至11月上旬是种子成熟期和收集期。[结论]该研究为进一步筛选和鉴定优良种源提供基础资料。
关键词 米老排;资源分布;种实特性
中图分类号 S722 文献标识码 A 文章编号 0517-6611(2015)11-159-02
米老排(Mytilaria laosensis Lecomte)是金缕梅科(Hamamelidaceae)壳菜果属(Mytilaria)常绿阔叶树种,主要分布于我国广东、广西、云南3省(区)及东南亚的越南、老挝等地[1]。其干形通直,材质优良,适于制作家具、胶合板、纸浆等,是珍贵优良的工业用材树种[2]。其树脂道细胞分泌物及种仁富含脂类物质,是重要的生物质油脂树种[3]。米老排根深叶茂,水土保持与培肥等生态防护功能优异,是生物防火林、水源涵养和土壤改良优选树种[4-8]。叶子肥大嫩绿,营养成分含量高,也是一种农家饲料树种[9]。自20世纪80年代,我国开展了米老排的引种栽培、育苗造林、生长特性等多方面研究[10-14],取得了可喜成果。中国林业科学院热带林业实验中心主持的“米老排中间试验与组装配套技术的研究”获1989年度林业部科技进步三等奖。而种质资源收集与保育研究缺乏,造成了当前米老排良种选育及推广栽培迟滞。种质资源保育和优质良种选育已经成为当前米老排林木遗传改良首要任务,为此该研究开展了米老排资源分布调查与种质保育,为下一步筛选和鉴定出优良种源和选育优良品种提供基础研究资料。
1 材料与方法
1.1 种群选择
2012年10~11月上旬对广西壮族自治区米老排分布区进行了全面的实地调查。米老排天然居群零星分布于广西南部、西南部7个县和广西凭祥市中国林业科学院热带林业实验中心人工林,共计选择8个参试种源。
1.2 采种优势木性状测定
采种优势木应间隔距离50 m,避免花粉相互传播,并测量树高、胸径等生长指标和记录采种地经纬度、海拔等地理位置信息。
1.3 统计分析方法
利用软件SPSS17.0进行米老排8个种源共82株优势木胸径、树高之间的相关性分析和回归分析。
2 结果与分析
2.1 资源分布
米老排分布区调查结果(表1)表明:米老排天然种质资源稀少,零星分布于广西壮族自治区龙州、靖西、德保、那坡、东兴、上思、容县7个县,集中分布在21°32′~23°11′N,105°51′~110°34′E,海拔179~1 202 m之间的低中山区。作为珍优阔叶用材树种培育与经营样板基地,广西凭祥中国林业科学院热带林业实验中心米老排资源丰富,人工造林面积500 hm.2,可作为米老排种质资源保存和用材林培育基地。
2.2 树高与胸径的相关性
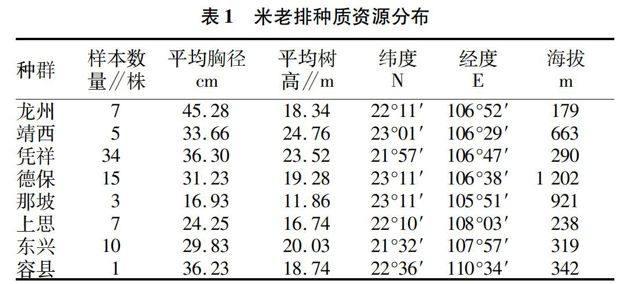
树高和胸径是米老排良种选育重要的数量评价指标。不同种群之间,胸径和树高差异显著。龙州米老排平均胸径最大;靖西、凭祥、德保、容县米老排次之,那坡平均胸径最小;平均树高以靖西、凭祥、东兴种群较好,超过20 m,龙州、德保、容县次之,那坡平均树高最小。对米老排胸径和树高进行相关性分析结果表明:8个种源82株优势木树高与胸径显著正相关(r=0.526,P=0.000<0.01,图1),说明米老排优势木树高随胸径的增加而增大。树高、胸径2因素数学模型拟合结果(表2)表明:8种数学模型拟合相关性达到显著水平,相关系数R.2在0.526~0.749间波动,相关系数最大的是S型曲线方程,最小的是线性方程,说明S型曲线拟合度最高。
2.3 径级分布规律
从米老排的径级结构来看(图2),米老排径级分布范围较广,平均胸径为33.4 cm,最大胸径为75.7 cm,最小胸径11.9 cm。米老排的径级呈近似正态分布:在10~80 cm径级范围内呈连续分布,10~40 cm径级范围内随着径级的增大个体数逐渐增多,径级为40 cm个体数最多,之后随径级的增大个体数逐渐减少,50 cm径级后的个体数大幅减少,各径级仅为1株。径阶结构中,径级<20 cm的个体数占优势木总数的10.345%。20~40 cm的个体数占优势木总数的73.563%。>40 cm的个体数占优势木总数的16.092%。米老排建群种以大径材为主,小径材少,表明该树种群落优势种群缺乏可供更新的幼苗、幼树,种群呈现衰退趋势。
2.4 树高分布规律
从米老排的树高分布规律来看(图3),米老排树高分布范围较广,平均树高为20.8 m,最高为32.8 m,最小为8.4 m。米老排的树高分布呈正态分布:在5~35 m范围内呈连续分布,5~20 m范围内随着树高的增加个体数逐渐增多,之后随树高的增加个体数逐渐减少。5~10 m范围个体数占优势木总数的3.448%,10~15 m的个体数占优势木总数的5.747%,15~20m个体数占优势木总数的41.379%,20~25 m的个体数占优势木总数的33.330%,25~30 m的个体数占优势木总数的11.494%,30~35 m的个体数占优势木总数的4.598%。
2.5 种实特性
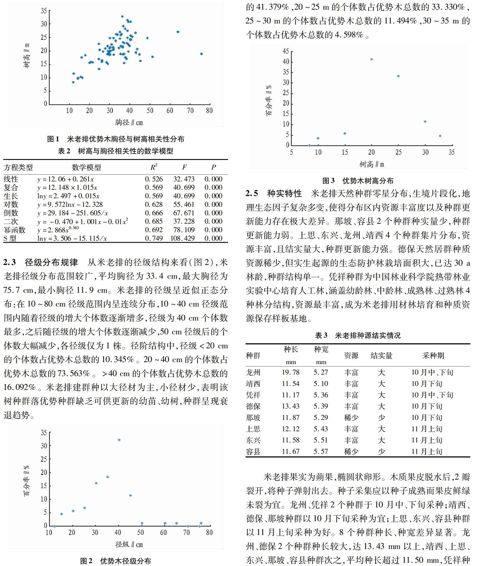
米老排天然种群零星分布,生境片段化,地理生态因子复杂多变,使得分布区内资源丰富度以及种群更新能力存在极大差异。那坡、容县2个种群种实量少,种群更新能力弱。上思、东兴、龙州、靖西4个种群集片分布,资源丰富,且结实量大,种群更新能力强。德保天然居群种质资源稀少,但实生起源的生态防护林栽培面积大,已达30 a林龄,种群结构单一。凭祥种群为中国林业科学院热带林业实验中心培育人工林,涵盖幼龄林、中龄林、成熟林、过熟林4种林分结构,资源最丰富,成为米老排用材林培育和种质资源保存样板基地。
米老排果实为蒴果,椭圆状卵形。木质果皮脱水后,2瓣裂开,将种子弹射出去。种子采集应以种子成熟而果皮鲜绿未裂为宜。龙州、凭祥2个种群于10月中、下旬采种;靖西、德保、那坡种群以10月下旬采种为宜;上思、东兴、容县种群以11月上旬采种为好。8个种群种长、种宽差异显著。龙州、德保2个种群种长较大,达13.43 mm以上,靖西、上思、东兴、那坡、容县种群次之,平均种长超过11.50 mm,凭祥种群种长最小,平均仅为11.17 mm。容县种群种宽最大,为5.57 mm,其次为东兴,种宽为5.51 mm,德保、凭祥、那坡、龙州4个种群种宽分别为5.39、5.36、5.29、5.27 mm,靖西种群种宽最小,仅为5.10 mm。
3 结论与讨论
3.1 米老排分布特点
米老排是我国热带南亚热带速生丰产乡土用材树种,自然分布于20°30′~23°50′N,105°45′~112°00′E之间的广东南部、西部,广西西南部和云南南部。广西是米老排种质资源分布的中心,集中分布在22°11′~23°11′N,105°51′~110°34′E,海拔179~1 200 m之间的低中山区。境内生境片段化严重,地理生态因子复杂多变,使得米老排资源丰富度以及种群演化存在差异。靖西、德保、那坡3个种群地理位置相近,但由于地理隔离,种群之间缺乏基因交流。东兴、上思2个种群呈连续分布,花粉漂移引发基因迁移或重组的可能较大。龙州、容县2个种群相对独立,与其他种群缺乏基因交流。
3.2 种群结构
长期以来我国南方省份工业用材原料多取自松、杉、桉,乡土用材树种资源保护及利用不足,导致米老排种质资源破坏严重,种群分布和数量日益减少,亟待种质资源保育和适应对策研究。对不同居群米老排采种优势木径级结构研究表明,不同居群采种优势木大径木、老龄株数多,说明大龄材流失严重;幼龄株数少,缺乏小径材、幼龄树,说明幼龄种质损坏严重,种群更新能力弱[15-16]。
3.3 种实特性
种子表型性状受遗传因素和环境2个因素互相影响,体现在种长、种宽等数量评价指标差异,研究其种子表型性状是揭示天然居群遗传变异及其格局的有效途径[17-18]。米老排分布呈零星状态,地理分布跨度大,从西部地区一直到东南沿海,分布区内的温度、湿度、无霜期和降水量等环境因子差异显著。生境片段化引起生殖隔离以及分布区内选择压力和基因流动差异,使得米老排的表型变异较大。对8个居群采种优势木种子表型性状差异研究表明:种长、种宽等表型性状种间差异显著。受物候规律影响,各居群种实成熟期也存在显著差异,龙州、凭祥种群应于10月中旬收集,靖西、德保、那坡种群在10月下旬收集,东兴、上思、容县种群在11月上旬收集。居群内采种优势木存在大小年现象,其影响机理尚需进一步研究。
参考文献
[1]祁承经.树木学[M].北京:中国林业出版社,2003:282.
[2] 梁善庆,罗建举.人工林米老排木材解剖性质及其变异性研究[J].北京林业大学学报,2007,29(3):142-148.
[3] 黄正暾,王顺峰,姜仪民,等.米老排的研究进展及其开发利用前景[J].广西农业科学,2009,40(9):1220-1223.
[4] 卢立华,贾宏炎,何日明,等.南亚热带6种人工林凋落物的初步研究[J].林业科学研究,2008,21(3):346-352.
[5] 王光玉.杉木混交林水源涵养和土壤性质研究[J].林业科学,2003,39(S1):15-20.
[6] 李夷荔,林文莲.杉木米老排混交林水源涵养功能的研究[J].福建水土保持,2001,13(4):43-46.
[7] 林德喜,韩金发,肖正秋,等.米老排对土壤理化性质的改良[J].福建林学院学报,2000,20(1):62-65.
[8] 田晓瑞,舒立福,乔启宇,等.南方林区防火树种的筛选研究[J].北京林业大学学报,2001,23(5):43-47.
[9] 景跃波,杨德军,马赛宇,等.热带速生树种米老排的育苗与造林[J].林业实用技术,2008(1):21-23.
[10] 李良昌,廖翠兰,黄德林,等.米老排在乐昌林场的引种及生长表现[J].广东林业科技,2003,19(4):66-68.
[11] 刘济祥,何伟民.米老排引种试验及繁育技术研究[J].江西林业科技,2006(5):17-19.
[12] 吴光枝,张万幸,温恒辉,等.米老排容器育苗试验[J].中国林业,2008(10):57.
[13] 李炎香,谭天泳,黄镜光,等.米老排造林密度试验初报[J].林业科学研究,1988(2):206-212.
[14] 郭文福,蔡道雄,贾宏炎,等.米老排人工林生长规律的研究[J].林业科学研究,2006,19(5):585-589.
[15] 邵方丽,余新晓,吴海龙,等.冀北土石山区天然次生林山杨、白桦种群的空间分布格局[J].林业科学,2012,48(6):12-17.
[16] 韩照祥,山仑.栓皮栎种群变异与适应性对策研究[J].林业科学,2005,41(6):16-22.
[17] 曾杰,郑海水,甘四明,等.广西西南桦天然居群的表型变异[J].林业科学,2005,3(2):59-65.
[18] 李文英,顾万春.蒙古栎天然群体表型多样性研究[J].林业科学,2005,41(1):49-56.
责任编辑 朱淼 责任校对 李岩
