NO在GA3抑制离体蒜薹细胞内含物转运过程中的作用
2015-04-24屠荫华骆桥妹张丽华牛军鹏段眉会
屠荫华,惠 伟,骆桥妹,张丽华,牛军鹏,段眉会
(1.陕西师范大学生命科学学院,陕西西安710062;2.岐山县食品药品监督管理局,陕西宝鸡722400;3.周至县农业机械化学校,陕西西安710400)
NO在GA3抑制离体蒜薹细胞内含物转运过程中的作用
屠荫华1,2,惠 伟1,*,骆桥妹1,张丽华1,牛军鹏1,段眉会3
(1.陕西师范大学生命科学学院,陕西西安710062;2.岐山县食品药品监督管理局,陕西宝鸡722400;3.周至县农业机械化学校,陕西西安710400)
蒜薹细胞内营养物质再转运严重影响其商品价值和食用价值。本文研究了一氧化氮(NO)参与赤霉素(GA3)延缓蒜薹衰老过程中细胞内含物转运的生理机制,为解决此类园艺产品的贮藏保鲜提供实验依据。实验用GA3、SNP、GA3+HB、GA3+L-NAME、GA3+Na2WO4、SNP+PP333分别处理离体蒜薹基部3 h,干燥后于4℃下贮藏,定期测定薹苞直径、蒜薹薹苞失水率和内源NO等指标。结果表明:GA3和NO都可抑制薹苞直径的增大和离体蒜薹的大量失水;清除NO或抑制NO合成的任一途径,都会削弱GA3对细胞内含物再分配的抑制作用,由此得出,GA3延缓离体蒜薹物质的再转运可能通过NO起作用。
蒜薹,NO,GA3,保鲜,物质再转运
园艺产品在收获后会出现许多外观变化,如绿色果褪绿黄化,黄瓜梗端果肉组织萎缩发糠,花端部分发育膨大,瓜条呈棒槌型等,这些现象是由果实内物质发生转移和再分配造成的。蒜薹(大蒜的花薹):包括花茎(薹茎)和总苞(薹苞)两部分。薹苞是大蒜花茎顶端的总苞,内含发育不全的花序[1]。蒜薹采后变黄,基部薹茎纤维化,薹苞膨大开裂并长出气生鳞茎,以致食用品质降低。研究表明蒜薹含有多种营养成分,具有很高的食用价值,其所含营养成分高于蒜头[2]。此外,蒜薹中含大蒜素和大蒜新素,具有较强的杀菌作用[3],李时珍在《本草纲目》中就已详述了其药用价值[4-5]。但是蒜薹在采收后含水量往往很高,在贮藏期间极易老化,较难保鲜,若不经处理,在常温下保存7~9 d[6]就会丧失商品价值。五十年代中期,我国植物细胞生物学家与生理学家以葱、蒜类植物为材料对细胞衰退过程中原生质的胞间运动进行了系统广泛的探讨[7-9]。还有人以蒜薹为材料研究了细胞内含物的转运问题,认为在贮藏过程中,细胞内含物由衰退薹茎向薹苞转移并被再利用,致使顶端气生珠蒜生长[10]。蔡可等的研究表明,GA3处理蒜薹基部可提高薹茎活力,明显防止薹茎衰老和阻抑细胞内含物向珠蒜转移,从而起到防衰的作用[11]。经前人研究,GA3对延缓离体蒜薹衰老的调节作用可能与物质再分配有密切的关系[12]。
NO是植物体内重要的信号分子,有关其在果蔬采后保鲜方面的研究是近几年的焦点。李琴[13]以SNP为蒜薹NO供体,发现它能够明显地延缓蒜薹失重率的增加,抑制蒜薹薹苞膨大和可溶性固形物含量的变化,有效减慢叶绿素降减的速率,保持其固有品质。有较多的证据表明,NO及其衍生物(N2O)通过延缓植物组织衰老进程、抑制乙烯合成和作用来提高果蔬贮藏过程中抵御逆境的能力,从而延长货架期,并改善果蔬采后贮藏的品质[14]。NO可在植物中合成,并可能作为一种植物生长调节物质而作用于植物的成熟和衰老[15]。研究发现,浓度为0.1 mmol/L的外源NO处理能够抑制蒜薹薹苞的膨大,降低失水率,延缓蒜薹的衰老,并可抑制基部内含物向顶端转运,即其对蒜薹采后保鲜具有显著的效果[16]。GA3抑制营养物质转移过程是否与NO有关,NO是否参与其中,有关此方面的研究未见报道。因此,本文旨在研究GA3延缓蒜薹营养物质转移过程中NO的作用及其生理机制,为解决此类园艺产品的贮藏保鲜问题提供科学依据。
1 材料与方法
1.1 材料与仪器
实验所用的材料 为无机械损伤、生长均匀的苍蒜蒜薹,采自陕西兴平蒜薹基地;NO供体硝普钠(SNP)、二氨基荧光素二乙酸酯(DAF-2DA,NO的荧光探针)、牛血红蛋白(HB)、NG-硝基-L-精氨酸-甲酯(L-NAME,NO合酶抑制剂)、二甲基亚砜(DMSO)Sigma公司;其他试剂 均为分析纯;10 mmol/L DAF-2DA 将DAF-2DA溶解于DMSO溶液配成10 mmol/L的浓度,分成小包装冷冻保存、10 mmol/L Tris、5 mmol/L KCl,pH7.2、10 μmol/L DAF-2DA在500 μL的Tris-KCl缓冲液中,加入0.5 μL的DAF-2DA母液混匀。
TCS-SP2型激光共聚焦扫描显微镜 Leica Lasertechnik Gmbh,Germany。
1.2 实验方法
1.2.1 实验处理 本实验以蒸馏水浸泡为对照,共7个处理(具体见表1)。各处理用50枝蒜薹,将薹茎基部10 cm以下部分浸入溶液中3 h,取出后晾干,置于4℃培养箱中暗贮藏四周。每个处理重复三次。
1.2.2 生理指标的测定 薹苞直径用游标卡尺测量;失水率用称量法测定,每周一次。以上各指标均重复测量3次,取其平均值。
薹苞增加率(%)=(终薹苞直径-初始薹苞直径)/初始薹苞直径×100
失水率(%)=(初始质量-终质量)/初始质量×100
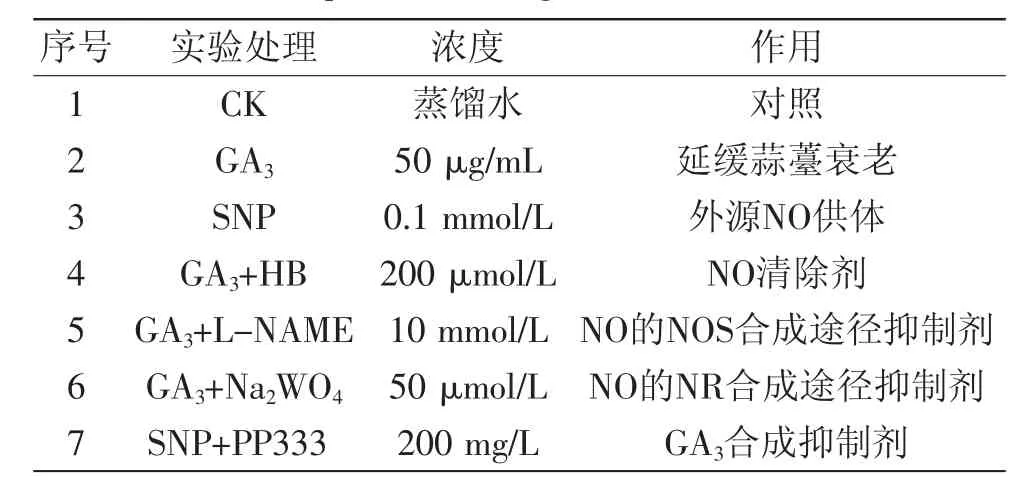
表1 实验试剂及处理浓度表Table 1 Experimental reagents and concentrations
1.2.3 NO荧光探针的负载 预先以1%(v/v)的DMSO为溶剂将NO荧光探针DAF-2DA配制成10 mol/L母液,于-4℃下冷冻保存。每次在基部1 cm处取样,徒手横截面切片。装载时将各处理切样放入Tris-KCl缓冲液中,加入DAF-2DA母液混匀,DAF-2DA的最终浓度为10 μmol/L,混匀后置于27℃下避光孵育2 h。
1.2.4 NO的激光扫描共聚焦显微检测 将负载DAF-2DA的切样用新鲜Tris-KCl缓冲液漂洗6次,以洗去细胞表层多余的探针,然后置于加入少许Tris-KCl缓冲液的载玻片上,立即在激光扫描共聚焦显微镜下观察各处理组材料荧光强度的变化并获取图像。激光扫描共聚焦显微镜的工作条件为:激发波长(Ex)=488 nm,发射波长(Em)=505~530 nm,功率(Power)=3%,变焦(Zoom)=4,帧(Frame)=512×512,中速扫描。在相同的设置下获得荧光图像,用共聚焦显微系统配套软件(Leica Image Software)和Photoshop 7.0图像软件分析处理。每个处理至少重复5次,当结果一致时,在10倍镜下取图,取其中1/4截面进行比较分析。荧光强度按表2计算。

表2 蒜薹组织内部结构荧光分级表Table 2 The level of fluorescence in the internal structure of garlicscape tissue
1.2.5 数据处理方法 采用SPSS数据统计软件进行多重分析比较。
2 结果与分析
2.1 不同处理对离体蒜薹薹苞直径的影响
通过测定贮藏4周后不同处理的薹苞的增加率可知,GA3和SNP处理过的薹苞直径增加率显著低于对照;GA3+HB处理组与GA3处理组相比,薹苞直径增加率大于GA3处理组,但不显著,说明清除内源NO后,GA3对蒜薹薹苞直径增加率的抑制作用有所降低;GA3+L-NAME和GA3+Na2WO4处理组与GA3处理组的薹苞直径增加率没有显著差异;SNP+PP333处理组和CK组比较,薹苞直径增加率抑制效果显著,说明外源GA3能够抑制薹苞的膨大;SNP处理组和SNP+PP333处理组比较,SNP抑制薹苞直径增加率的作用有所减弱,结果如图1所示。
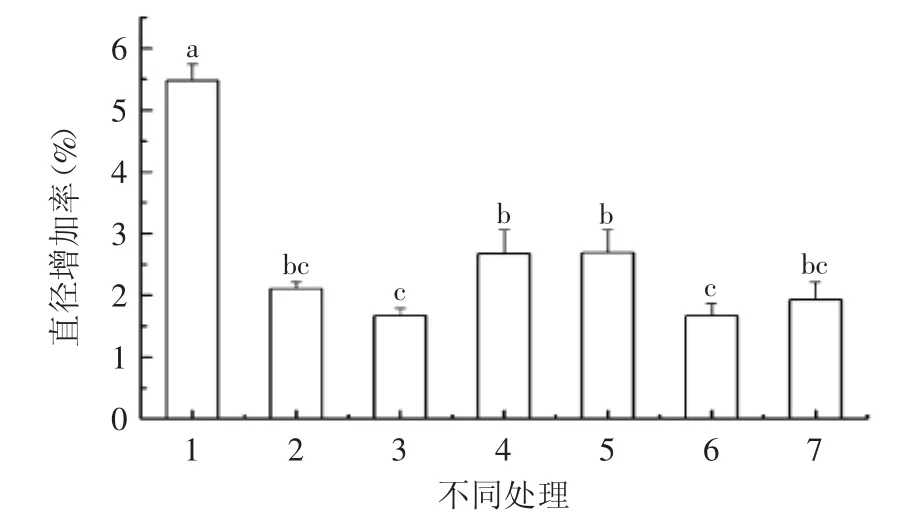
图1 不同处理对薹苞直径增加率的影响Fig.1 Effect of different treatments on the increasing rate of the diameter in bulbet
2.2 不同处理对离体蒜薹薹苞失水率的影响
通过测定处理后贮藏4周的蒜薹失水率可知,CK组的蒜薹失水率最高,为9.6%;GA3处理组的失水率最低,为2.3%;SNP处理组的失水率为5.7%。该结果表明GA3和SNP处理均可抑制蒜薹水分的散失。GA3处理组和GA3+HB处理组比较时发现,GA3+HB处理组蒜薹失水率显著增高,也即:当加入HB后,GA3抑制蒜薹失水的作用显著降低;GA3处理组与GA3+L-NAME相比,GA3+L-NAME处理组抑制蒜薹失水率的作用显著升高;而GA3处理组与GA3+Na2WO4处理组比较时,效果显著;加入GA3合成抑制剂PP333后和CK相比,发现其对蒜薹失水率有十分显著的影响;SNP+PP333处理组与SNP处理组比较,SNP+PP333处理组对蒜薹失水率的抑制作用更为显著。

图2 不同处理对蒜薹失水率的影响Fig.2 Effect of different treatments on water loss rate in the garlic scape
2.3 自然状态下蒜薹的组织形态结构
在光镜下,苔茎横切面的蜡质层呈波浪状,厚度为2.5 μm,角质层为5 μm。表皮细胞为一层排列紧密的长砖形细胞。表皮组织内侧为皮层组织,它主要由薄壁细胞组成,近表皮的几层薄壁细胞小且具有叶绿体,内层的薄壁细胞大。细胞之间有间隙,皮层组织的最内层薄壁细胞之间比较规则地排列着一轮形状小而密集的周缘维管束。中央由大量薄壁细胞组成,细胞直径为50~70 μm,胞间有大量的细胞间隙,在中央薄壁细胞间散生大量比周缘维管束粗大的中央维管束,如图3所示。
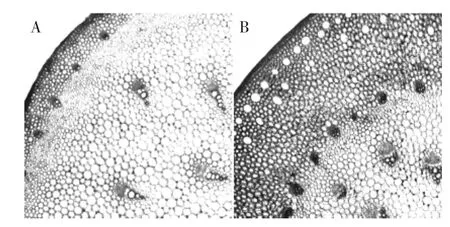
图3 蒜薹组织横断面形态结构部分图(×10)Fig.3 The cross section structure of garlic scape(×10)
2.4 不同处理对离体蒜薹组织NO含量和分布的影响
2.4.1 GA3处理对蒜苔内部NO的水平及分布状况的影响 以CK荧光强度为标准(中)将荧光按强度划分为无、弱、强和最强4个等级(见表2)。用NO专一荧光探针DAF-2DA负载蒜薹不同部位的组织,48 h内直接观察内源NO含量的变化。由图4(a,b)可以看出,CK组基部组织中NO主要集中在中央维管束基部组织中,基部组织中荧光强度为弱,顶部组织中NO主要集中在周缘维管束和中央维管束中,且荧光强度等级也为弱。
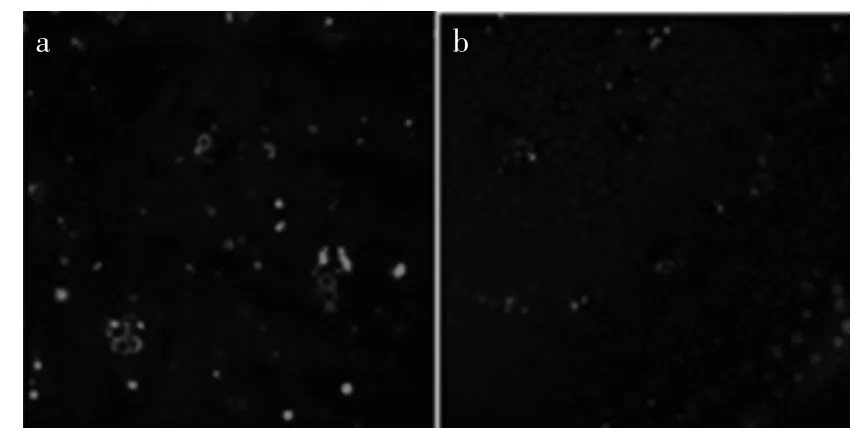
图4 对照组蒜薹内部NO的水平及分布Fig.4 Distributed condition and level of NO in control garlic scape
由图5(a,b)可以看出,GA3处理组基部组织中NO除在微管组织中没有分布外,在其他组织中都均匀分布,荧光强度等级为强,顶部组织中NO的分布情况与基部组织中相同,但NO含量很高,荧光等级为最强。说明外源GA3引起了内源NO的积累,尤其在生长旺盛的顶端组织细胞中其含量更高。
2.4.2 SNP处理对蒜薹内部NO水平及分布的影响SNP处理组的基部组织中NO均有分布,且皮层组织中NO的含量高、荧光为强,维管束中也含有较高水平的NO,强度为强;顶部组织中的分布及强度同基部组织,中央维管束细胞中的含量略高于其他组织细胞中的含量。如图6(a,b)所示。NO的分布和水平说明了物质再转运的程度,外源GA3引起内源NO的积累,可能通过控制NO水平的变化而抑制蒜薹衰老过程中细胞内含物从基部向顶部的转运。
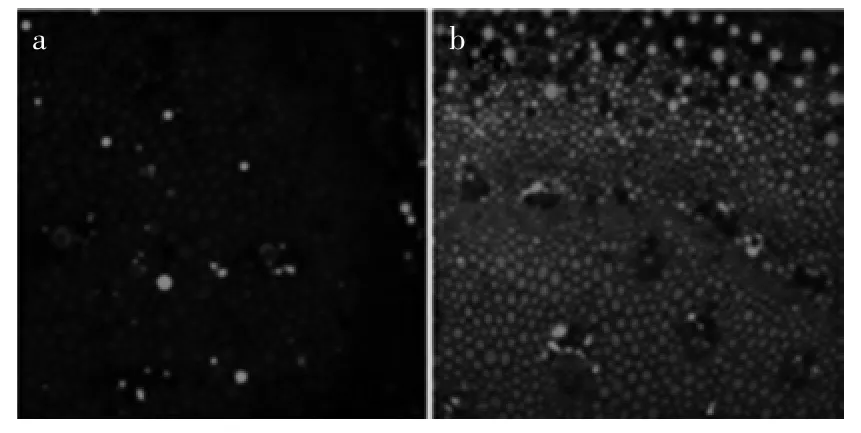
图5 GA3处理组蒜薹内部NO的水平及分布Fig.5 Distributed condition and level of NO in GA3treatments
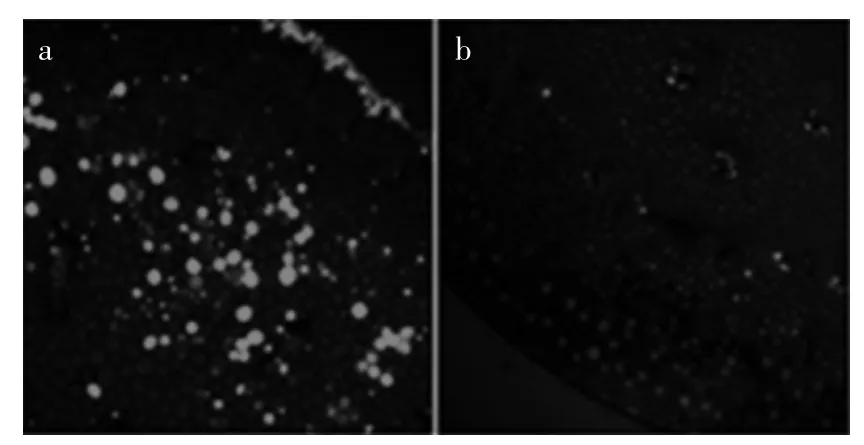
图6 SNP处理组蒜薹内部NO的水平及分布Fig.6 Distributed condition and level of NO in SNP treatments
2.4.3 GA3复合HB、L-NAME、Na2WO4处理对蒜薹内部NO水平及分布的影响 如图7(a,b)所示,GA3复合HB处理组与GA3处理组相比,整个蒜薹组织细胞中NO的分布和水平均发生了变化,基部和顶部组织细胞中的NO消失,荧光强度等级均为无。

图7 GA3+HB处理蒜薹内部NO的水平及分布Fig.7 Distributed condition and level of NO in GA3+HB complex treatments
GA3+L-NAME复合处理与GA3处理组相比,NO的分布和含量发生了显著变化,基部组织细胞中无NO荧光,顶部组织细胞中荧光强度有所减弱,且荧光都集中在中央维管束细胞中,周围薄壁细胞中未见荧光分布,见图8(a,b)。

图8 GA3+LNAME处理组蒜薹内部NO的水平及分布Fig.8 Distributed condition and level of NO in GA3+L-NAME complex treatments
GA3+Na2WO4复合处理组与GA3处理组相比,基部组织细胞中NO的分布和强度没有显著变化,但顶部组织细胞中NO有所变化,薄壁组织细胞中无NO,中央维管束细胞中有极弱的NO分布,见图9(a,b)。
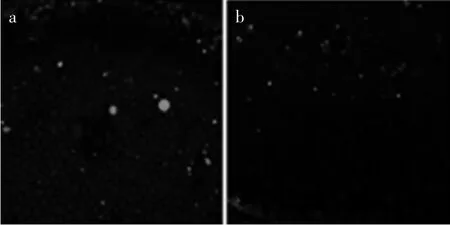
图9 GA3+Na2WO4处理组蒜薹内部NO的水平及分布Fig.9 Distributed condition and level of NO in GA3+Na2WO4complex treatments
2.4.4 SNP复合PP333处理对蒜薹内部NO水平及分布的影响 由图10(a,b)可知,与CK组和SNP处理组相比,SNP+PP333处理组整个组织中NO的分布均发生了显著变化,其荧光水平增强,基部组织细胞中除维管束细胞NO消失,NO均匀的分布在薄壁组织细胞中,且顶部组织细胞中都分布有高水平的NO,尤其在皮层组织细胞中其含量最高,周缘维管束细胞中也含有较高水平的NO。外源NO引起的内源NO的变化小于降低内源GA3水平所引起的内源NO的变化幅度,因此可知:降低内源GA3的水平,可以引起内源NO的积累。
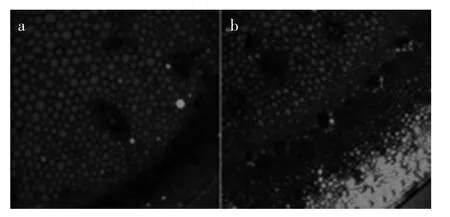
图10 不同处理的蒜薹内部NO的水平及分布Fig.10 Distributed condition and level of NO in SNP+PP333 complex treatments
3 讨论
NO在植物体内主要经一氧化氮合酶(NOS)和硝酸还原酶(NR)催化而形成,且NO有双重作用,高浓度的NO会抑制生长,甚至会毒害植物,低浓度的NO可以促进植物生长,可以延缓离体果蔬衰老[17]。NO的含量与植物的生理状态有关,生长旺盛的部位NO含量较高[18]。有人[19-20]已证明,用一定浓度的GA3处理蒜薹的基部,可以有效抑制细胞内含物再分配的过程,抑制蒜薹衰老并阻抑细胞内含物向珠蒜转移,从而起到延缓衰老的作用。本实验用50 μg/mL的GA3处理蒜薹基部,有效的抑制了蒜薹薹苞的膨大,降低了蒜薹的失水率,有效抑制细胞内含物再转运的过程,并且蒜薹组织中NO含量水平较高,这与前人研究结论相同;GA3处理引起内源NO分布和水平的变化:对照蒜薹组织中的NO主要分布在微管组织中,但经GA3处理后,NO的含量和分布发生了变化,并且GA3处理引发了蒜薹组织中NO水平的升高,尤其是顶部组织中NO的积累。加NO清除剂或抑制剂时,GA3的这种效应减弱,说明GA3延缓蒜薹衰老起到保鲜作用的效果是通过NO实现。而在SNP+PP333处理中,NO的分布变化不显著,但NO的含量显著提高,由此可知,降低内源GA3的水平,也可影响NO的水平。或许物质再转运过程中,不同阶段二者相互影响,究竟二者在物质再转运过程中通过何种方式传递信号,还有待进一步研究。
4 结论
本文首次对NO在GA3延缓蒜薹营养物质转移过程中的作用及其生理机制开展了研究,通过测量薹苞膨大率、蒜薹失水率和内源NO含量,得出如下结论:a.GA3和NO均有抑制物质再转运的作用;b.GA3延缓蒜薹营养物质转移中的作用是通过调控NO得以实现的;c.NO的分布与转运程度有关,在生长旺盛的部位,NO均匀的分布于组织细胞中,而衰老部位的NO主要分布在维管束细胞中;d.外源NO延缓物质再转运的作用GA3参与其中。二者相互影响的研究结果为进一步研究此类园艺产品有关贮藏保鲜的问题打下了良好基础。
[1]张华云,王善广,郭颖,等.蒜薹形态特征对其耐藏性的影响[J].保鲜与加工,2001,1(5):15-17.
[2]王修俊.蒜薹采收后生理变化和贮藏保鲜的研究[J].贵州工业大学学报,2000,29(5):28-31.
[3]陈萍,朱连平.气调贮藏对蒜薹品质的影响[J].现代化农业,2005(5):38-39.
[4]王善广.蒜薹、蒜头及洋葱贮运保鲜实用技术[M].北京:中国农业科学技术出版社,2004.
[5]王杰.大蒜和洋葱的风味化学[J].食品科学,1987,8(2):41-43.
[6]杨艳荣.蒜薹的贮藏保鲜技术[J].农村实用科技信息,2008(4):25.
[7]娄成后,邵莉楣,段静霞.高等植物衰老叶片中原生质的撤退现象以及原生质运动在有机物运输中可能具有的作用[J].植物学报,1973,15(2):204-216.
[8]张蜀秋,张培元,娄成后.蒜薹贮藏过程中某些生理变化和物质再分配[J].北京农业大学学报,1993,19(3):105-106.
[9]王毅,娄成后,杨世杰.离体蒜薹贮存中薄壁细胞超微结构的变化[J].植物学报,1994,36(3):165-169.
[10]黄承祥,花宝光,王学成,等.贮藏蒜薹中细胞内含物由衰退茎组织向顶端珠蒜的分配[J].北京农业大学学报,1982,8(2):1-14.
[11]蔡可,娄成后.贮藏过程中蒜薹细胞内含物再分配的激素控制[J].植物生理学报,1985,11(4):403-408.
[12]张映璜,方建雄,张炎.离体蒜薹衰老过程中乙烯的产生和纤维素酶活力的变化[J].植物生理学报,1990,16(2):206-208.
[13]李琴.NO对蒜薹的保鲜作用研究[J].天津农业科学,2010,16(6):57-60.
[14]Leshem Y Y,WiLLs R B H.Harnessing senescence delaying gases nitric oxide and nitrous oxide:a novel approach to postharvest control of fresh horticultural produce[J].Biological Plantarum,1998,41(1):1-10.
[15]Leshem Y Y,Haramaty E.Plant aging:the emission of NO and ethylene and the effects of NO-releasing compounds on growth of pea(Pisμm sativμm)foLiage[J].Plant Physiol,1996,148:258-263.
[16]屠荫华,惠伟,李彩香,等.硝普钠(SNP)对蒜薹保鲜效果的影响[J].食品工业科技,2008(10):541-547.
[17]Wendehenne D,Pugin A,Klessig D F.Nitric oxide:comparative synthesis and signaling in animal and plant cells[J]. Trends Plant Science,2001,6(4):177-183.
[18]Kelm M,Dahmann R,Wink D.The nitric oxide/superoxide assay.Insights into the biological chemistry of the NO/O2interaction [J].Biol Chem,1997,272(15):9922-9932.
[19]ZHAO Lin-chuan,HAN Juan-juan.Effects of GA on Metabolism of Hydrogen Peroxide during Redistribution of Cell Contents in Excised garlicScape(Alliμm Sativμml)[J]. DevelopentaL&Reproductive Biology,2000,9(2):43-52.
[20]屠荫华,惠伟,冯德霞,等.NO对GA3延缓蒜薹衰老的影响[J].陕西师范大学学报:自然科学版,2010,38(1):82-88.
Effect of NO on the process of GA3inhibiting cell inclusions transport in the excised garlic scape
TU Yin-hua1,2,HUI Wei1,*,LUO Qiao-mei1,ZHANG Li-hua1,NIU Jun-peng1,DUAN Mei-hui3
(1.Shaanxi Normal University,College of Life Sciences,Xi’an 710062,China;2.Qishan County Food and Drug Administration,Baoji 722400,China;3.Zhouzhi County Agricultual Mechanization School,Xi’an 710400,China)
The redistribution of nutrients in the cells seriously affected the commodity value and edible value of garlics cape.This paper studied the physiological mechanism that NO participates in the process of GA3delaying garlics cape senescence via the cell inclusions transport,in order to provide experimental basis for the storage and refreshing of such kind of horticultural products.The basal part of garlic scape were treated with GA3,SNP,GA3+L-NAME,GA3+Na2WO4,GA3+HB or SNP+PP333 for three hours,respectively.Dried in the air and then stored at 4℃.Regularly measured diameter,water loss rate and level of endogenous NO in the garlic scape bulbet.The results showed that the increase of diameter and the loss of water in the garlic bulbet were inhibited by both GA3and NO treatments.All treatments that eliminating NO or preventing any synthesis pathway of NO reduced the effect of GA3in inhibiting the cell content redistribution in the garlicscape of Allium Sativum.It was concluded that NO might play a important role in the process of GA3delaying material transfer in garlic scape.
garlicscape;NO;GA3;refreshing;material transfer
TS201.1
A
1002-0306(2015)22-0325-05
10.13386/j.issn1002-0306.2015.22.058
2014-12-23
屠荫华(1980-),女,硕士研究生,工程师,主要从事果蔬采后贮藏和食品安全检验检测研究,E-mail:tuyinhua@126.com。
*通讯作者:惠伟(1964-),女,硕士,主要从事植物衰老及果蔬采后贮藏方面的研究,E-mail:huihui@snnu.edu.cn。
国家农业科技成果转化项目(2013GB236000662);陕西省农业科技创新项目(2012NKC01-22)。
