医学分子生物学进展诠释着病毒的起源和进化
2015-04-17杨漫宇匡林林MAJianjieISAACSWilliams徐学红
周 鑫, 杨漫宇, 匡林林, MA Jianjie, ISAACS Williams, 徐学红
(1. 陕西师范大学 生命科学学院,陕西 西安 710062;2. Ohio State University Medical Center Columbus OH, USA;3. Hopkins University School of Medicine Baltimore MD, USA)

·实验技术·
医学分子生物学进展诠释着病毒的起源和进化
周 鑫1, 杨漫宇1, 匡林林1, MA Jianjie2, ISAACS Williams3, 徐学红1
(1. 陕西师范大学 生命科学学院,陕西 西安 710062;2. Ohio State University Medical Center Columbus OH, USA;3. Hopkins University School of Medicine Baltimore MD, USA)

在人类依然抗争着禽流感病毒、艾滋病病毒以及SARS病毒等对生命威胁和享受现代医学及分子生物学的最新研究成果的同时,思考病毒的起源和进化的分子机理是痛定思痛且必须关心审视的医学科学问题。病毒是介于生命与非生命间的非细胞生命体,关于它的起源及进化有3种观点:①自起源理论,病毒先于细胞产生于生命前地球,然后进化成原核细胞及真核细胞;②平行进化理论,病毒和细胞是在生命前环境条件下同时平行起源而成,进化过程中形成寄生于细胞的关系;③细胞源退化理论或逆向理论,病毒是细胞形成之后,向外“抛出”的大分子或其复合体,来源于并寄生于细胞。近些年的分子生物学的一些进展揭示:理解并接受病毒起源的多元性,认同病毒在生命历史起源进化的多机制特征,是回答自然赋予的这一生物医学课题命题的最合理答案。
病毒起源; 逆转录; 朊病毒; 起源与进化多元性
0 引 言
病毒是目前地球上所发现的最小生命单位,是一种最简单的非细胞结构生命形式,其生活和繁殖必须借助于其他生命个体的细胞结构生命个体[1]。从多细胞高等动物植物到仅有一个细胞的动植物和微生物,从结构和功能高度分化的真核细胞到进化地位公认低于真核细胞、结构和功能相对简单的原核细胞,都可以是其赖以生存的宿主。病毒在离开其宿主时,不能表现出生命的最主要特征——繁殖,即使是在充满构建生命大分子材料的环境中,细胞结构生命体足以生存的情况下,没有其所需的特定宿主,病毒依然不能繁殖,这显然是一种非生命状态;非生命状态的病毒,只有在进入细胞后,或者说接触到宿主细胞时,才表现生命状态,进入细胞的遗传物质利用宿主的遗传复制装置和蛋白质合成装置以及相关的分子调控系统,制造起自身繁殖所需的组件蛋白质,和能使自身特性稳定遗传于子代病毒的基因组核酸[1]。显然,病毒这种生命形式是一种介于生命状态和非生命状态间的准生命形式。
关于病毒的起源及进化地位,目前可归纳简述为:① 自起源理论,该理论认为病毒先于细胞产生于生命前(pre-biotic)地球,然后进化成原核细胞及真核细胞;② 平行进化理论,该学说认为病毒和细胞是在生命前环境条件下同时平行起源而成,进化过程中形成病毒寄生于细胞的关系;③ 细胞源退化理论或逆向理论,依据相关证据该理论认为,病毒最初是细胞中的组份,进化过程中逐渐丢失掉寄生生活中非必需的能够独立生活的基因,最初是由于某种原因,细胞向外“抛出”的大分子、大分子复合体或“小细胞”,来源于并寄生于细胞。这3种假定起源程式,最终形成现在存在的病毒的完全寄生方式。虽然,部分生物学工作者偏向于第3种观点,但从近些年的现代分子生物学和医学分子生物学的一些重大进展来看,特别是瑞典皇家科学院将诺贝尔生理学及医学奖授予对现代医学影响经久不息的[2-5]“朊病毒(prion)”的发现者,加州大学医学院的Stanley B Prusiner;时隔几年后,又将该奖项授予了拯救千万人生命[6-8]的导致人类宫颈癌的“疱疹病毒(human papilloma viruses)”的发现者和导致人类免疫性缺陷疾病的逆转录RNA病毒“人类免疫缺陷病毒HIV(human immunodeficiency virus)”的发现者,德国癌症研究中心的Harald zur Hausen和法国巴斯德研究所的Fran?oise Barré-Sinoussi及世界艾滋病研究防护基金会的Luc Montagnier[9-11],次年又授予霍普金斯大学医学院的Carol W. Greider等,以表彰其发现揭示癌症及衰老奥秘[12-14]的具逆转录特性的端粒酶;这都使广大关注这一重大自然命题的哲学及自然科学工作者认识到,摒弃另外两种观点是不科学的。在已经或依然抗争着禽流感病毒、艾滋病病毒以及SARS病毒等的过程中,享受着现代医学及分子生物学的最新研究成果的同时,也思考着病毒起源与进化的基础科学问题,而只有理解并接受病毒起源的多元性,才应是回答自然赋予的这一神秘命题的最有说服力的答案。
1 病毒的类型、生命的特征及起源的相关理论
病毒不仅生命形式低级,而且维持其低等生命形式的组份,比较于细胞结构生命也显得极为简单。通常所见的病毒,氨基酸单位重复构成位于病毒表面的蛋白质规则多面体形状,遗传物质核酸包裹于其中,部分病毒表面还包有主要由双亲性脂类构成的囊膜。数种蛋白质、一条核酸即构成了这一生命形式,极为简单。更有甚者,仅有一条RNA单链的类病毒和仅有一条肽链的朊病毒,也像其他常见病毒一样具有感染性,这两种病毒即是到目前为止发现的世界上最简单的生命。历史上,疯牛病在西方人日常生活上留下了可怕阴影,这种导致他们主要食品来源危机的朊病毒性家畜疾病,即是这种最简单生命形式寄生于高等脊椎动物,极端复杂且高度分化的真核细胞所致,疯牛病朊病毒寄生的结果是直接导致畜牛大脑神经元细胞死亡,从而在脑组织形成“空洞”。同时朊病毒也会导致人类疾病,如克雅二氏病,根据脑部受破坏区域不同症状也不同,致死率极高[15]。极强的致死性,使这种最简单生命形式的病毒的种类十分稀少。
自Miescher从外伤病人绷带上脓细胞的细胞核中分离出富磷有机化合物——核素(即DNA)后,Avery以其经典的肺炎球菌转化实验精巧地证明转化因子是DNA以来,科学家们已公认核酸是一切生物遗传的物质基础。虽然依据病毒的大小,形态结构和易感染宿主等性质,有详尽的分类系统,但通常可根据核酸的种类将病毒分为两大类,即DNA病毒(包括双链DNA病毒和单链DNA病毒)和RNA病毒(包括双链RNA病毒和单链RNA病毒)。前者可以转录翻译成蛋白,后者可以反转录成DNA最后进入经典的中心法则[16-17],逆转录RNA病毒的发现,如AIDS病毒HIV和其在动物体及人类中的传播方式,如它仅仅在大猩猩这一特有的动物体内能像在人类中发病一样以及在恒河猴体内能以潜伏无症状的表象传播以外,在其他动物体内均不能存活以及分子生物学上各类机理的发现[18-21]表明:这一独特的病毒与宿主的关系,使得病毒在进化上的地位更加扑朔迷离,再加上以上朊病毒,这种引起广大科技工作者广泛重视的简单病毒形式。这种从生物大分子本质上的分类,为我们探讨研究病毒的起源和进化地位提供了极具生物学本质特性的有效依据和途径。
上述这种介于生命与非生命,既依赖于细胞才能表现出生命特征,又较细胞明显简单的准生命形式,其在地球生物物种中的地位以及起源,自病毒发现之日,就引起自然科学家的极大兴趣。自1898年Beijerinck首次证实细菌过滤性烟草花叶体,实际上是另一种生命形式,并命名为病毒以来,已有一个多世纪。百年多的努力,人类依然尚未弄清病毒的起源与进化地位,在地球生物的进化树上,我们仍未为病毒这一生命形式找到其确切的位置[16]。
因为病毒这一生命形式具有的独特生物学特征,它的起源与进化必然与细胞(包括原核细胞和真核细胞)有着内在的联系。目前,病毒与细胞在起源上的相关性,有以下3种理论[17]:
随着现代生物学和医学进入分子研究水平,病毒起源的这3种假说也正在接受着检验。当然,检验的结果并不一定就是某两个观点的错误,某一观点的正确,两种或三种以及加之其他观点的产生相互补也是可能的。诚然,病毒进化和起源的多元性更应引起自然科学工作者以及生物医学工作者的重视。
2 现代医学分子生物学进展与病毒起源理论
2.1 自起源理论:病毒起源于生物大分子前体、进化地位先于细胞
现代医学生物学的发展已经深入到了揭示生命本质的分子水平的阶段,其中也包括对地球生命起源的探索。特别是近二三十年来,医学分子生物学的研究成果成批涌现,分子生物学技术的不断突破,人类对生命的认识日新月异,一些老的理论由于新知识的注入,而重现活力。其中,病毒起源与进化的理论也因此而渐现明了。自起源理论认为在远古地球,小分子肽及寡聚核苷酸等生物分子前体形成“亚生命单位”,这些“亚生命单位”即为或即将发展为亚病毒/病毒,然后与其他分子,如脂类分子等自组装/相结合,如包被于脂类分子形成细胞的雏形,这种细胞雏形具有现代细胞的基本结构,脂类分子将小分子肽及寡聚核苷酸与外界“恶劣”环境隔绝的同时,也为它们进一步聚合、特化、偶联以及协同形成生物大分子提供了不可缺少的环境,最后形成原核细胞然后进化成真核细胞。在该理论中,显然亚病毒/病毒在进化地位上先于亚细胞/细胞。
《RNA世界(RNA World)》的讨论专集提出“RNA世界”这一概念,即是在产生生命之前的时期,生物大分子是以RNA占统治地位的。此概念的提出是基于美国科罗拉多大学Cech等的工作,他们的研究表明,RNA分子具有自身拼接的功能,在一定的条件下,RNA分子也可以具有使单核苷酸聚合成寡聚核苷酸(即RNA)的功能,这一结果大大扩展了人类对生物酶的认识,酶不仅是蛋白质的概念得到了重要扩充,即RNA也是一种酶(核糖核酸质酶),RNA可以自身催化自己的拼接,可以复制自己[22]。Ekland等在世界著名科学杂志《Nature》上报导RNA可以以自身为模板化三磷酸核苷聚合,其正确配对率较错配率高1 000倍[23]。这些实验表明,RNA极可能在没有蛋白质或者肽出现的生命前时期的条件下,独立“生存”并将其“遗传信息”延续给子一代RNA分子。然后RNA分子再与其平行发生的多肽相结合,形成一种RNA-蛋白质复合体,当其陷入生命前期的“囊泡”[16]后即形成了原始的细胞。设想,在生命前时期,具逆转录酶性质的蛋白酶将RNA逆转录成DNA,最后形成了具稳定遗传特性的原始原核细胞,然后通过其相应的进化途径最终发展出真核细胞[24-25]。实验表明,这一设想的病毒进化成细胞的途径,在现代细胞中仍有着明显的痕迹。如,1989年Varmus报导的细菌的逆转录现象[26],以及近些年来的分子生物学研究热点——真核细胞染色体稳定遗传的必需组分之一,端粒酶,即是蛋白质与RNA的复合体,其蛋白质组分能以此RNA为模板反转录成DNA[12],这一2009年获得诺贝尔生理及医学奖的工作在生物医学领域造成了极大的影响[25],同时也表明细胞内本身就具有反转录功能,它向胞外的“抛出”形成现在的逆转录病毒也是可能的。
自起源理论认为,在当今出现的病毒种类上仍然保持着进化的痕迹,如仅由RNA分子构成的类病毒不被一些病毒学权威分类归为病毒类,而属亚病毒类。又如人类丁型肝炎病毒具有与类病毒相雷同的RNA基因组,不能合成蛋白质衣壳,靠利用乙型肝炎病毒衣壳而生存,这些被认为是“卫星病毒”的亚病毒物质进化地位明显低于病毒的进化中间体。虽然自起源理论是几十年前普遍公认而随后又极有可能被自然科学工作者抛弃的观点,但现代生物学的发展,特别是近些年来医学分子生物学的重大进展无疑为这种行将过时的观点注入了新的血液,使它重新成为了一种不可轻视的理论。
2.2 平行进化理论:病毒进化平行于细胞,然后建立与细胞的依赖关系
平行进化理论与自起源理论都认为病毒起源独立于细胞起源,不同的是后一学说认为,如前所述在远古地球上病毒先于细胞出现于地球,并为细胞进化的先期生命形式。而平行进化理论认为,远古生命有几种不同的形式,生命前体分子以两条相对独立的进化途径分别形成病毒和细胞;现在病毒依赖于细胞生命的格局是进化后期,病毒为更有效地应对环境而进化成的,是适应进化的渐变的结果进而在我们面前呈现出复杂的不同的病毒与细胞间的关系。
Piccirilli在《Nature》杂志上发表评述指出,在生命前期的地球环境中,首先产生能自我复制的前RNA(pre-RNA),有RNA进化压力的迫使而导致的DNA/蛋白质世界(DNA/Protein world)出现,直至现今复杂的包罗万象的生物世界[27]。同期《Nature》上,Bohler及其同事报道了一种结构上既不同于RNA,又不同于蛋白质的大分子肽核酸PNA(peptide nucleic acid)[28],PNA以多聚甘氨酸为骨架(这一结构通常为蛋白质的基本结构),以亚甲基与碱基(通常为核酸的主要组成部分)相键合, 两条结构互补的链形成双股结构。PNA不仅外观上与双股核酸相似,其复制行为上也与其相同,依照Waston-Crick互补复制原则而自我复制。而且PNA在结构组成上以及动态化学行为上均介于蛋白质和核酸之间。Piccirilli和Bohler及其同事均认为,PNA是生命前地球环境中出现的早于Pre-RNA的物质,极有可能是RNA的前体[27-28]。
随后Lee及其同事在《Nature》上报导了肽(蛋白质结构的亚单位)可以由比其更小的肽复制而成的实验结果[29]。虽然肽复制的机理尚待研究,但由于这一现象完全违背Watson-Crickz中心法则,暗示自然界还存在另一种不同于经典的碱基配对的复制体系,当时引起了生物学界的强烈反响。《Nature》杂志编辑部委托著名的生物学家Kauffman在同期刊物上发表对这一实验结果的评议[30],他认为这一现象的发现,在RNA世界尚有许多问题仍待解决的时候,提出我们生命前时期还可能存在另一个(或取代前一个)生物起源发生的途径。显然,肽可以进行自身复制就如同RNA可以自身复制一样,在生物起源及进化上具有同等重要的意义。
综合以上分析,Piccirilli提出的地球上生命进化起源的可能过程[27]可进一步充实如下:
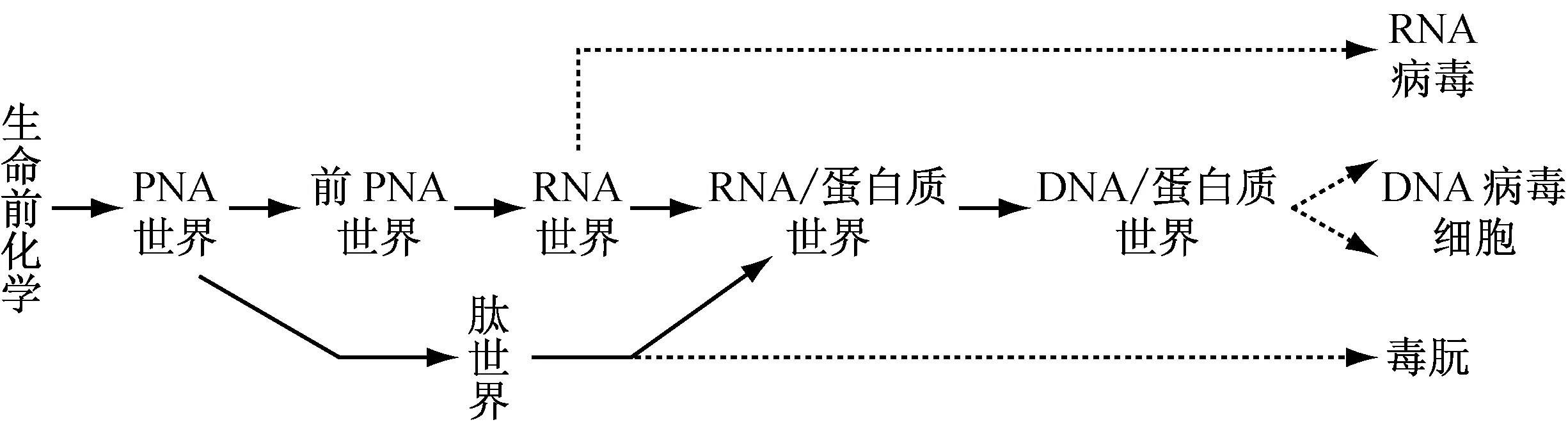
*注:---作者补充的各类病毒起源的可能途径
根据以上讨论,细胞由DNA/蛋白质世界逐渐演化而成的同时,与之平行,RNA世界可能演变出RNA病毒(如类病毒),DNA病毒可由DNA/蛋白质世界演化而成,而肽世界则可能进化产生朊病毒,如图中虚线所示。这一理论在当今生物世界有着很有趣的代表性生物,如朊病毒最早发现于绵阳瘙痒症,后又发现也会导致人类库鲁和雅克综合症。这种朊病毒为氨基酸形成的类似肽结构,它自身可以复制。正常生物中有与其一级序列相同的蛋白和编码这些蛋白的基因,但它们的高级结构却决然不同。朊病毒的侵入会使内源的肽结构发生变化,将后者转化成朊病毒[2, 3, 5]。从这些最新的分子生物学发现和相关疾病机理的研究来看,病毒与细胞平行进化,然后形成与后者的依赖性寄生的进化观点也是非常合理的。
2.3 细胞源退化理论或逆向理论:起源于细胞产生于细胞的病毒进化起源理论
病毒起源于细胞的观点是当今生物学界占主导作用的理论[17],有大量的实验支持这一观点,这里不作赘述。这一理论认为,病毒是细胞在其进化过程中“抛出”的一个细胞组分,由于它来源于细胞,所以与细胞有着不可割舍的密切关系,又由于它虽然源于细胞却又只是细胞的一个组分,所以在细胞之外不能独立繁殖生存,它的生命特征必须依赖于细胞来表现。然而,这些细胞的组分为何会被细胞抛出?这一过程是细胞的主动过程还是被动行为?是一些细胞死亡留下的活性大分子,待时机成熟时再回到细胞?这是一个适应进化的过程呢还是选择过程?许多问题当前分子生物学的最新进展尚不能解释,它们的分子机制是目前该理论尚未解决的问题。
从现代细胞分子生物学的研究结果来看,当今细胞中仍然存在与病毒结构功能接近的类似结构组分。例如,RNA病毒基因组通常具有逆转录酶基因,而正常生长的原核细胞中也存在逆转录酶,这说明其基因组中也包括有相应的逆转录酶基因片段;另外,近些年来的研究结果表明[25],真核细胞遗传物质要稳定流向子一代细胞,染色体上必须的DNA片段之一即是每条染色体线性脱氧核糖核酸的端部必须存在端粒序列,而这一特定序列是不直接受上代DNA模板控制的,它是由一种被称为端粒酶的来复制完成的。如前所述,端粒酶是由一个起模板作用的RNA片段和一个起逆转录作用的蛋白酶构成,后者以前者为模板逆转录成DNA,并连接于真核细胞每一条染色体DNA双链的末端。显然,端粒酶的RNA片段和蛋白部分在真核细胞的基因组中都应具有相对应的基因。生物学家们通常认为,这些组分形成的复合体的“抛出”,即是最早的病毒。
显然,病毒起源观点三的关键是“抛出”的合理性。那么,在现代生物中是否有这种现象呢?答案是肯定的。从现在我们认识到的生命过程来看,多种情况下细胞会抛出其某些组分。某些动物种类在进化过程中,细胞中染色体数目有一个片段的流失;转座因子(mobile element)现象的发现就是一个有力的证据,真核细胞中有一些基因在一定情况下,会从线性DNA链的位置通过RNA的介导逆转录至另一位置。以上两种过程中,如遇到该细胞偶然死亡,细胞的破裂则会导致DNA或RNA组分的外泄,即“抛出”。细胞凋亡(apoptosis)的正式发现使得“死亡与生命生存在生物世界同等重要”的观点在医学分子生物学界被广泛接受,这一现象的发现者及主要贡献者S Brenner,H R Horvitz 和J E Sulston于2002年10月7日共享诺贝尔生理和医学奖[31]。这一死亡形式是真核细胞死亡的常见形式,是细胞的一种正常的死亡方式。所说的正常是指它是生物体发生发育生存繁衍所必需,是一种受细胞自身自主控制的死亡,是一种主动的死亡过程。细胞在启动其程控死亡后,细胞核内DNA链会被随机、非特异性地切割成小的片段。如细胞在正常死亡过程中,遇外界原因而突然瓦解,这些片段则被抛出细胞之外,显然,细胞中的任何一个片段或碎片都有这种机会。在现今生物界中,质粒经常能游离于微生物细胞之外,又能进入相应的细胞之中,已被视为某些病毒起源的可能模式[16]。综合以上分析,上述过程的发生,可能会导致各类病毒的起源,抛出的DNA片段(含蛋白质)可能会形成DNA病毒;抛出一个具逆转录功能的RNA(也可能含有蛋白质),则可能形成RNA病毒;抛出可自我复制的肽,这可能形成朊病毒。
3 病毒起源、进化理论观点的多元性
许多医学分子生物学近期的研究进展都表明,像物种进化一样,地球上病毒的进化仍然在永不停息地进行着,感冒病毒的变异进化就是一个典型的例证。流感病毒如禽流感H7N9以及其他同类病毒有一个共同的特点就是基因的变异性极强[32-34],表现在病毒表面蛋白上即是结构不断变化,导致每次流行病域的流感感冒病毒对人类都有很大的影响,预制的疫苗都不能起到相应的作用。这样的病毒特性有不少仍然对人类有着重大的影响,又例如先天性免疫缺陷疾病艾滋病病毒HIV,从发现它的存在到现在,已有30年的历史,疫苗的研制工作也几乎有相同的时间,虽然病毒作用机理上的研究结果不断有突破性的进展[10-11],但是仍然没有通常意义上的疫苗研制成功。发生在这些常见的或罕见的病毒身上的作为进化的基本动力的变异表明病毒中确实存在现代分子生物学已有原则不能解释的“不明机理”,而且我们认为这种特异性不明机理明显表示各种病毒对细胞的依赖性的发生是迥然而异的,预示着在变异特性进化的不停息的同时,各种病毒进化理论都是可能存在的。
2007年,科罗拉多大学和米兰大学的研究者发现,小到几对核苷酸的DNA 寡聚物就可以在体外自组装形成液晶状态,这是现今为止我们发现的最早的可能的生命前体状态,无需蛋白、RNA或任何其他可能的生命大分子,依靠自身的自由能形成有序的类似前期生命的形式[35],这一发现似乎更支持DNA是最早形成的生命起源的假想,但它是否会先形成DNA病毒显然是不确定的。
病毒是如何起源的,它是在地球生命前时期先于细胞产生,作为简单的细胞前生命体,然后发展进化成?还是在漫长的化学进化过程中与细胞平行进化而同步产生,而后再与细胞形成初步的寄生关系,物竞天择,最后成为完全的寄生关系?或者是它本身就是细胞的某一或某几个大分子组份,因某种因素暂时性地以非生命的状态独立于细胞之外,再次与细胞接触时,又能重返细胞形成寄生的生命状态,从而周而复始地进行着自己的准生命过程?生物学家及自然科学工作者们至今尚不能确定其确切途径。
自然界中,生物的起源和进化是一个极其漫长而复杂的过程,从达尔文经典的物种起源理论,到现代分子生物学不断发展的进化学说,无一不体现这一特性。显然,病毒的起源和进化也不例外,并且,依以上的分析和讨论,更充分地表现出进化的多样性。因而,病毒的起源及进化极有可能存在不同的途径,DNA病毒、RNA病毒以及朊病毒分别会依照其自身的轨迹发祥发展至今。虽然,病毒进化的结果具很强的统一性,如其繁殖特性表现出对真核及原核细胞的完全依赖性,和脱离细胞后完全无生命性质的特点,它们的进化轨道并不重要,其进化途径很可能是具多元性。起源理论观点三虽然看起来比较完善,大量的事实都表明它对解释DNA病毒和某些RNA病毒的起源是比较合理的,有较为充分的研究结果支持,但对解释某些特殊的病毒如仅有一个RNA分子的类病毒,仅有一个肽分子的朊病毒,则显示出明显的不足。生物学家们尚未确定细胞中存在能自我复制且能催化蛋白质合成的RNA分子,也尚未有确切的证据表明在细胞中,肽能行使自我复制的功能。如细胞中不具这种功能的组份,则不可能向外“抛出”它们,以形成现今的类病毒和朊病毒。这里,病毒起源观点一和观点二则显示出其优势。在生命前时期地球的某些环境可能造就能自我复制的前肽物质世界,也可产生能独立复制、自我拼接的前RNA分子,独立于细胞之外,可能产生现今类病毒和朊病毒的前身,然后与细胞建立寄生关系,以发展成类病毒和朊病毒。看来,病毒起源观点一和观点二并不能简单地被否定,相反,它们很可能是观点三所必需的补充。虽然,朊病毒的发现者S B Prusiner获得了诺贝尔生理学及医学奖,但由于这一发现的意义重大[3-5],所以有关朊病毒的争议尚未结束。当然,正如发现蛋白质具有自我复制现象的分子生物学家Lee所说的那样,即只有在发现蛋白质能以更小的肽段完成复制之后,才能进一步完善Watson-Crick遗传物质复制中心原则,同时使病毒进化、起源理论的多元性有更充分的实验证据。病毒起源这一自然留给我们的神秘命题尚未解决,而至今地球上新的病毒却在不断被发现。是否是生命前物理自组装进化原理任然在地球上进行尚不可断言[36],但用我们人类对生命本质不断深入的理解和认识来看,生命的发生与延续在某种意义上看是与细胞也是与病毒共同进化存在发展的过程,例如,整十年前的SARS病毒牺牲掉了众多人类生命的同时,我们战胜了病毒[37-39],但可以预料的是它们并不会从地球上彻底消失,而极有可能进入潜伏,待条件适宜时再以变异的形式重新再现。
4 结 语
不同种类的病毒在特性上有着十分重大的差异,它们在细胞中进行其生命过程的具体路线和涉及的代谢途径也差异颇大。由此推论,仅以一种病毒起源理论解释所有病毒在地球上的产生,明显有很大的局限性;而以多种起源理论相互补充,来看待病毒的起源进化问题才是科学的思维途径。科学的范式是多元化的范式,并不能以某一范式而统一。病毒起源理论的多元化,是我们早日确定各类病毒在地球生物进化树上的确切位置、完成大自然赋予的这一神秘问题的最合理答案。
[1] Susanne Modrow, Dietrich Falke, Hermann Schatzl,etal. Molecular Virology[M]. New York: Springer-Verlag Berlin and Heidelberg GmbH & Co K, 2013.
[2] Roy C, Gagné V, Fernandes M J,etal.High affinity capture and concentration of quinacrine in polymorphonuclear neutrophils via vacuolar ATPase-mediated ion trapping: comparison with other peripheral blood leukocytes and implications for the distribution of cationic drugs[J].Toxicol Appl Pharmacol,2013,270(2):77-86.
[3] Godsave S F, Wille H, Pierson J,etal. Plasma membrane invaginations containing clusters of full-length PrPSc are an early form of prion-associated neuropathology in vivo[J]. Neurobiol Aging, 2013, 34(6):1621-1631.
[4] Prusiner S B. Biology and genetics of prions causing neurodegeneration [J]. Annu Rev Genet, 2013,47:601-623.
[5] Watts J C, Giles K, Patel S,etal. Evidence That Bank Vole PrP Is a Universal Acceptor for Prions [J]. PLoS Pathog,2014,10(4):e1003990.
[6] Brundtland G H, Glinka E, zur Hausen H,etal. Open letter: let us treat patients in Syria[J]. Lancet, 2013,382(9897):1019-1020.
[7] zur Hausen H. Q & A: On the case. Interview by Michelle Grayson [J].Nature,2012, 488(7413):S16.
[8] Zur Hausen H. HPV vaccines: what remains to be done? Interview by Lauren Constable[J].Expert Rev Vaccines,2011,10(11):1505-1507.
[9] Ojesina A I, Lichtenstein L, Freeman S S,etal. Landscape of genomic alterations in cervical carcinomas [J].Nature,2014, 506(7488):371-375.
[10] Gallo R, Montagnier L. HIV's leading men [J].IAVI Rep, 2011,15(3):13-14.
[11] Andy Coghlan, “Scorn over claim of teleported DNA”[J]. New Scientist,2011, 2795:8-9
[12] Greider C W.Molecular biology.Wnt regulates TERT——putting the horse before the cart[J].Science,2012, 336(6088):1519-1520.
[13] Bender H S, Murchison E P, Pickett H A,etal. Extreme telomere length dimorphism in the Tasmanian devil and related marsupials suggests parental control of telomere length [J]. PLoS One,2012, 7(9):e46195.
[14] Tom H I, Greider C W. A sequence-dependent exonuclease activity from Tetrahymena thermophila [J]. BMC Biochem,2010,11:45.
[15] Wade Jr L G. Organic Chemistry[M].8th ed. London: Pearson PLC. 2013: 1194-1195.
[16] Alberts B. Molecular Biology of the Cell [M].5th ed. New York: Garland Science Publishing, Inc, 2008.
[17] 翟中和,王喜忠,丁明孝. 细胞生物学[M]. 4版. 北京:高等教育出版社,2011:23-29.
[18] Pépin J. The Origins of AIDS [M].Cambridge: Cambridge University Press,2011.
[19] Plantier J C. A new human immunodeficiency virus derived from gorillas [J].Nature Medicine, 2009, 15(8): 871-872.
[20] De Sousa J D. High GUD Incidence in the early 20th century created a particularly permissive Time Window for the Origin and Initial Spread of Epidemic HIV Strains [J]. PLoS ONE, 2010, 5(4): e9936.
[21] Levavasseur E, Privat N, Martin JC,etal. Molecular modeling of prion transmission to humans[J]. Viruses. 2014,6(10):3766-3777.
[22] Attwater J, Wochner A, Holliger P. In-ice evolution of RNA polymerase ribozyme activity [J]. Nature Chem,2013,5(12):1011-1018.
[23] Ekland E H. RNA-catalysed RNA polymerization using nucleoside triphosphates [J].Nature, 1996, 382(6589):373-376.
[24] Temin H M. RNA-dependent DNA polymerase in virions of Rous sarcoma virus [J].Nature, 1970, 226(5252):1211-1213.
[25] Singer T. Actuary of the Cell:AQ & A with Nobelist Elizabeth Blackburn on Telomeres and Aging Cells [J].Scientific American,2011, 305(4):84-87.
[26] Varmus H E. Reverse transcription in bacteria [J].Cell, 1989, 56(5):721-724.
[27] Piccirilli J A. Origin of life. RNA seeks its make [J]. Nature,1995,376(6541): 548-549.
[28] Bohler C. Template switching between PNA and RNA oligonucleotides [J].Nature,1995,376(6541): 578-581.
[29] Lee D H. A self-replicating peptide [J].Nature, 1996, 382(6591):525-528.
[30] Kauffman S. Self-replication. Even peptides do it [J].Nature,1996,382(6591):321-322.
[31] 张飞雄. 普通遗传学 [M]. 2版. 北京:科学出版社,2010:241.
[32] Wu A, Su C, Wang D,etal. Sequential reassortments underlie diverse influenza H7N9 genotypes in China [J]. Cell Host Microbe,2013,14(4):446-52.
[33] Worobey M, Han G Z, Rambaut A,etal. Genesis and pathogenesis of the 1918 pandemic H1N1 influenza A virus [J]. Proc Natl Acad Sci USA, 2014,111(22):8107-12.
[34] Huy C D, Sakoda Y, Nishi T,etal. Potency of an inactivated influenza vaccine prepared from A/duck/Mongolia/119/2008 (H7N9) against the challenge with A/Anhui/1/2013 (H7N9) [J]. Vaccine,2014, pii: S0264-410X(14)00598-2.
[35] Nakata M. End-to-end stacking and liquid crystal condensation of 6 to 20 base pair DNA duplexes[J].Science,2007,318(5854):1276-1279.
[36] 徐学红,张楚瑜,杨德才. 液晶生物学——正在崛起的交叉学科 [J].自然辩证法通讯,1997,19(2):76-80.
[37] Nieto-Torres J L, Dediego M L, Verdiá-Báguena C,etal. Severe acute respiratory syndrome coronavirus envelope protein ion channel activity promotes virus fitness and pathogenesis[J]. PLoS Pathog, 2014, 10(5):e1004077.
[38] Wang Z, Feng A, Cui M,etal. Design, Synthesis, Anti-Tobacco Mosaic Virus (TMV) Activity and SARs of 7-Methoxycryptopleurine Derivatives[J]. Chem Biol Drug Des, 2014, 84(5):531-42.
[39] Xiang Y T, Yu X, Ungvari G S,etal. Outcomes of SARS survivors in China: not only physical and psychiatric co-morbidities[J]. East Asian Arch Psychiatry, 2014,24(1):37-8.
The Origination and Evolution of Virus Base on Contemporary Progress of Molecular Bio-Medicine
ZHOUXin1,YANGMan-yu1,KUANGLin-lin1,MAJianjie2,ISSAACSWilliams3,XUXue-hong1
(1. College of Life Sciences, Shanxi Normal University, Xi’an 710062, China;2. Ohio State University Medical Center;3. Hopkins University School of Medicine)
Along challenging against life-threaten from various viruses for instance avian flu, AIDS, SARS, ect., and enjoying the progresses of molecular biomedicine, it is the time for us to decipher the deep fact on virus origination and evolution. Virus is a non-cell organism existing between life and non-life. It shows a complete parasitic characteristic in its way of living. Concerning its origination and evolution, there are three different hypotheses. One is that virus was generated before cell on the earth of pre-biotic stage. As a former organism, it would evolutinate to prokaryotic cell and eukaryotic cell. The secondary is that both cell and virus were generated simultaneously in the pre-biotic stage. During evolution, virus developed its parasitism on cells. The third is that virus is a macromolecule or macromolecular complex, "being thrown out of cells". It came from cells and was parasitic to cells. Three hypotheses address that eventually, virus evolutinated its complete parasitic style of living. Although the most of biologists believed the third hypothesis previously, they are paying their most attention to all these hypotheses nowadays regarding the proceedings of medical molecular biology. Both proceedings of molecular biology and natural philosophy show that the understanding to the pluralism of virus origination would be the most credible solution to assure the position of virus on the evolution tree on the earth, which accomplished the central dogma of molecular biology.
virus origination; reverse transcription; prion;pluralism of origination and evolution
2014-07-15
国家自然科学基金资助项目 (31371256);中央高校团队资助项目(GK201301001);国家教委外事司邀请项目(MS2014SXSF038);秦巴山区持续发展协同中心项目(201310718021)
周 鑫(1989-),女,山西长治人,博士生,主要研究方向为医学分子细胞生物学。
Tel.:18729043246;E-mail: zhouxin19890714@163.com
徐学红(1962-),男,江苏徐州人,博士, 教授,博士生导师,研究方向为医学分子生物学和发育生物学。
Tel.:13289838136;E-mail: xhx0708@snnu.edu.cn
Q 111
A
1006-7167(2015)05-0005-06
