主要营养素源组合对暗纹东方鲀生长、消化吸收和水环境的效应
2015-04-11王晓晨赵宁宁秦巍仑秦桂祥高婷婷
王晓晨 , 李 勇 , 张 静 , 赵宁宁 , 秦巍仑,秦桂祥 , 高婷婷
(1.中国科学院 海洋研究所, 山东 青岛266071; 2.中国科学院大学, 北京100049; 3.中国科学院 实验海洋生物学重点实验室, 山东 青岛 266071; 4.江苏中洋集团有限公司, 江苏 南通226600; 5.海洋生态养殖技术国家地方联合工程实验室, 山东 青岛266071)
暗纹东方 鲀 (Takifugu obscures), 俗称气泡鱼, 隶属于 鲀 形目(Tetraodontiformes)、鲀科(Tetraodontidae)、东方 鲀 属(Fugu), 分布于日本、朝鲜以及中国渤海、黄海、东海等海域, 属近海暖温水性底栖鱼类[1]。与鲥鱼(Tenualosa reevesii)、刀鱼(Coilia ectenesJordan)同为江海洄游性鱼类, 因过度捕捞、水环境恶化, 数量急剧减少, 已成为海江珍稀鱼类之一[2]。
目前, 东方鲀属(Fugu)鱼类已实现设施无毒化养殖, 但其营养饲料研究滞后, 如何研发出高效、低污染、低成本(一高二低型)专用配合饲料, 成为制约产业可持续发展、实现健康高效养殖的技术瓶颈。东方 鲀 属 鱼 类营养需求研究多见于红鳍东方鲀(Takifugu rubripes)[3-11], 暗 纹东方 鲀 仅见蛋白质和淀粉需求[12-13]。探求适宜营养素及营养源组合、获得暗纹东方 鲀 一高二低型专用配合饲料等研究尚属空白。
本研究在设施养殖条件下, 以现代动物营养学强调的组合效应、营养结构和系统调控等新理念为指导, 采用新颖的生态营养学原理和方法, 通过将两种营养素蛋白质和脂肪的不同水平与蛋白营养源鱼粉不同水平进行组配, 探 寻对暗纹东方 鲀 生长、消化吸收和水环境的影响, 初步确定主要营养素源的适宜组合水平, 为 开发暗纹东方 鲀 生态专用配合饲料提供理论和应用依据, 对珍稀鱼类优良遗传潜力发挥和规模化健康养殖具有重要价值和意义。
1 材料与方法
1.1 试验设计和分组
试验采用3×2×2三因素设计(表1), 2个营养素因素:蛋白质水平为43%、40%、37%分别表示为P43、P40、P37; 脂肪水平为10%、7%, 分别表示为L10、L7), 1个营养源因素: 鱼粉水平为高(49%~55%)、低(39%~ 45%),分别表示为 FH、FL。共 12个不同组合处理组:1#(P43L10FH)、 2#(P43L7FH)、 3#(P40L10FH)、4#(P40L7FH)、5#(P37L10FH)、6#(P37L7FH)、7#(P43L10FL)、8#(P43L7FL)、9#(P40L10FL)、10#(P40L7FL)、11#(P37L10FL)、12#(P37L7FL), 每处理3重复。
1.2 试验饲料
12种试验饲粮组成及营养水平见表1。 在江苏中洋集团河豚庄园营养中心饲料车间, 按照配方组成和比例, 将饲料原料进行一次混合, 超微粉碎, 再经二次混合, 制成粉状全价配合饲料, 包装备用。为适应设施养殖 暗 纹东方 鲀 的特殊采食习性, 饲喂前将粉状料、水以1.25/1的重量比例混合, 拌成黏性团状饲料, 进行投喂。

表1 试验处理与饲粮组成及营养水平(干物质)Tab.1 Treatments, composition, and nutrient levels of the experimental diet (DM)
1.3 试验用鱼和饲养管理
试验在江苏中洋集团河豚庄园养殖温室进行,试验鱼由河豚庄园提供。正式试验前, 将同一养殖温室同源同批 暗 纹东方 鲀 在试验条件下喂食同一种商品饲料, 使之逐渐适应环境和配合饲料。驯养 25 d后, 禁食 24 h, 挑选健康无病、规格整齐(172.87g±6.71g)的试验鱼1 080尾, 随机分到 36个圆形桶(直径1.5 m、水深0.6 m)中, 每桶30尾。
试验养殖密度为 4.89 kg/m3, 水温为 22~26℃,盐度为8~12, DO≥5.0 mg/L, pH为8.06~8.46。养殖期为77 d。试验期间, 每天7: 30、16: 00投喂, 将饲粮放食台上, 放入养殖桶, 半小时后取出食台, 将残饵风干称质量, 以剩少许残饵视为饱食。每天换水1/3, 添加一定比例的海水和淡水, 保持水体的稳定盐度。每天记录投喂量、残饵量, 根据摄食情况及时调整投喂量。发现死鱼及时捞出并称质量记录。
1.4 样品采集和指标测定
1.4.1 样品采集
试验结束后, 禁食24 h, 称质量。 每桶随机选取6尾中等体质量的试鱼, 用20~30 mg/L的MS-222做快速深度麻醉, 冰盘中解剖剥离肠和肝胰脏, 迅速将样品分装入封口袋, -20℃保存待测。取饲粮样品用四分法缩减至 200 g, 分装入封口袋, 进行营养成分分析。
1.4.2 饲粮营养成分
干物质采用失重法, 105℃烘箱干燥至恒质量;粗蛋白采用凯氏定氮法; 粗脂肪采用油重法, 即石油醚为浸提溶剂, 利用索氏抽提法抽提脂肪; 粗灰分采用焚烧法, 即样品炭化至无烟, 移至马福炉 550℃焚烧至恒重; 钙采用高锰酸钾法; 总磷采用钼黄比色法。
1.4.3 生长指标
存活率(%)=(终末尾数/初始尾数)×100; 日增质量(g/尾/日)=(终末均质量-初始均质量)/试验天数;增质量率(%)=[(终末体质量-初始体质量)/初始体质量]×100; 饲料系数=(总投喂量-总残饵量)/(终末体质量-初始体质量)。
1.4.4 消化吸收酶活力
使用南京建成生物工程研究所生产的试剂盒,按说明书测定暗纹东方 鲀 肠中(胰蛋白酶、脂肪酶、淀粉酶、碱性磷酸酶、Na+-K+-ATP酶)和肝胰脏中(脂肪酶、淀粉酶)的活性。测定前首先样品预处理, 加入生理盐水制备组织匀浆, 2 500 r/min离心10 min,通过预试验确定稀释倍数后, 进行正式指标测定。酶活性用比活力( U/mg蛋白或U/g蛋白) 表示。
1.4.5 水环境指标
试验结束前一周, 早上7: 30投喂结束, 将残饵称质量后, 换水2/3。次日 7: 30投喂前, 依次从 36个桶中取水样100 mL, 进行水环境指标测定。氨氮和亚硝酸氮在2 h内完成测定; 磷酸盐和COD在48 h内完成测定。氨氮采用纳氏试剂比色法; 亚硝酸氮采用盐酸萘乙二胺法; COD采用重铬酸钾法; 磷酸盐采用总磷总氮联合消化法。
1.5 数据分析
试验数据用 SPSS 21.0 软件进行统计, 采用 3因素方差分析和Duncan检验法统计分析。结果均用“平均值±标准差(Mean ± SE)”表示。
2 结果与分析
2.1 生长性能
12种营养素源组合饲粮下, 暗 纹东方 鲀 的生长性能结果见表2。
2.1.1 存活率、末质量、日增质量
生长结果(表2)表明: 各组存活率基本在90%以上, 且差异不显著(P>0.05)。蛋白质、脂肪单因素均对日增质量有显著或极显著影响(P<0.05或0.01)。双因素蛋白质和脂肪、鱼粉分别互作对日增质量有显著或极显著影响。3因素互作对日增质量有极显著影响(P<0.01)。单因素和因素互作对末质量的显著性影响与日增质量具有相似特征。
2.1.2 增质量率
表2中增质量率结果表明, 单因素脂肪、鱼粉均影响极显著(P<0.01), 蛋白质影响不显著(P>0.05);双因素蛋白质和鱼粉互作有显著影响。值得注意的是, P40和FH、FL分别互作, 效应均与P43FH差异不显著(图1)。3因素互作有显著影响(P<0.05), 最高的1#、9#分别较8#提高79.98%、54.32%; 低鱼粉组11#与高鱼粉组4#、5#均无显著差异。
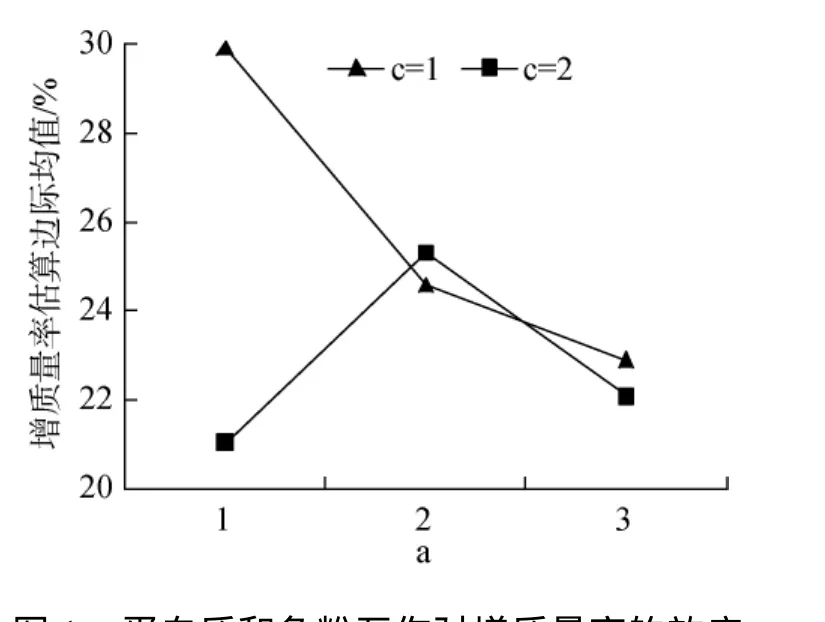
图1 蛋白质和鱼粉互作对增质量率的效应Fig.1 Effects of protein and fish meal interactions on WGR
2.1.3 饲料系数
表2中饲料系数结果表明, 单因素蛋白质、脂肪均有极显著影响(P<0.01)。双因素蛋白质和脂肪、鱼粉分别互作均差异极显著(P<0.01)。值得注意的是,P43L10和 P40L10, P43FH和P40FL分别差异不显著(图2, 图3)。3因素互作差异显著(P<0.05), 最低的1#和 9#分别较 8#降低 76.35%、64.15%; 较低的 3#和 7#差异不显著; 低鱼粉组 11#和高鱼粉组 2#、6#也无显著差异。
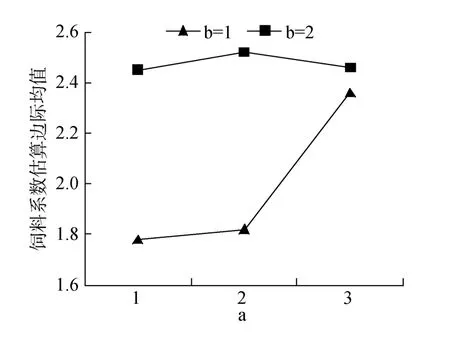
图2 蛋白质和脂肪互作对饲料系数的效应Fig.2 Effects of protein and lipid interactions on FCR

图3 蛋白质和鱼粉互作对饲料系数的效应Fig.3 Effects of protein and fish meal interactions on FCR

表2 主要营养素源组合对暗纹东方鲀生长和饲料利用的影响Tab.2 Effects of the main nutritional element and source combination on the growth and feed utilization in Takifugu puffer
2.1.4 增质量率和饲料系数的对应关系
12种营养素源组合饲粮下, 暗 纹东方 鲀 的增质量率和饲料系数的对应关系(图4)表明, 增质量率和饲料系数呈明显的负相关关系, 即增质量率越低,相应的饲料系数越高, 符合常规, 表明生长试验数据客观而可信。
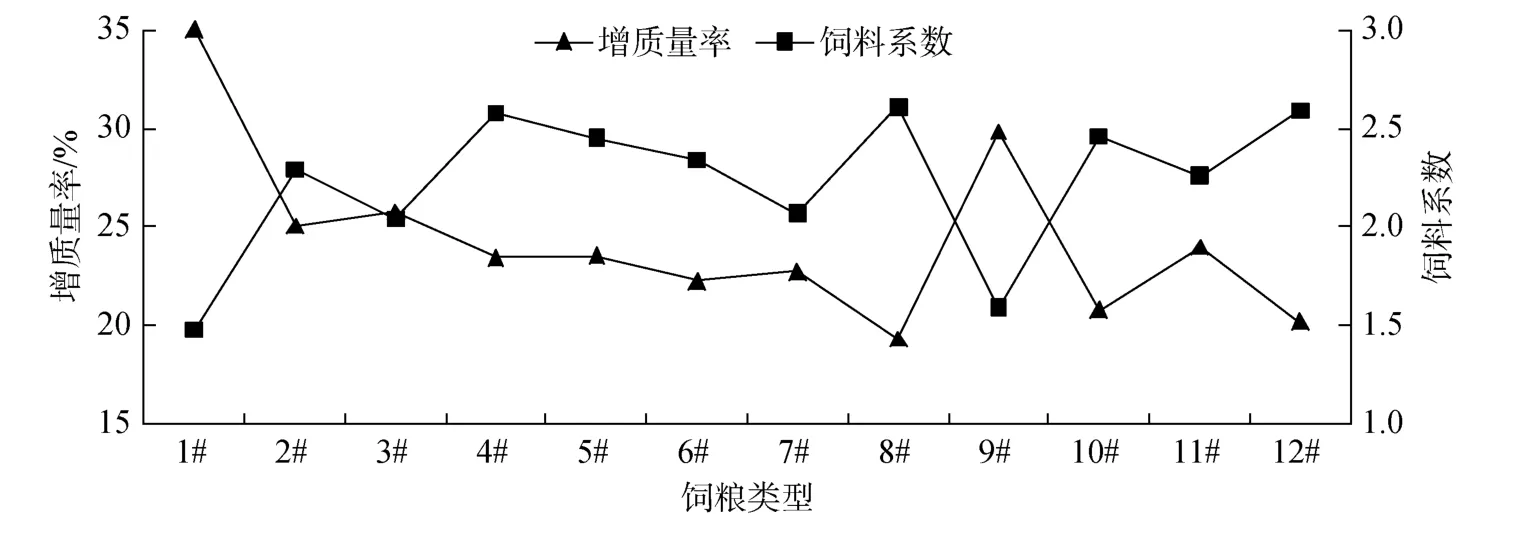
图4 主要营养素源组合对增质量率和饲料系数的影响对应趋势Fig.4 Trend of the main nutrient element and source combination effects on WGR and FCR
2.2 消化酶活性
12种不同营养素源组合饲粮下, 暗 纹东方 鲀 的消化酶活性结果见表3。
2.2.1 胰蛋白酶
表3中胰蛋白酶活性结果表明, 单、双、3因素互作均有显著或极显著影响(P<0.05或0.01)。P43、P40分别与鱼粉互作效应显著(图5)。L10FL互作效应显著优于 L7FL、L10FH、L7FH(图6)。3因素互作差异极显著(P<0.01), 9#活性最高, 8#、7#次之。
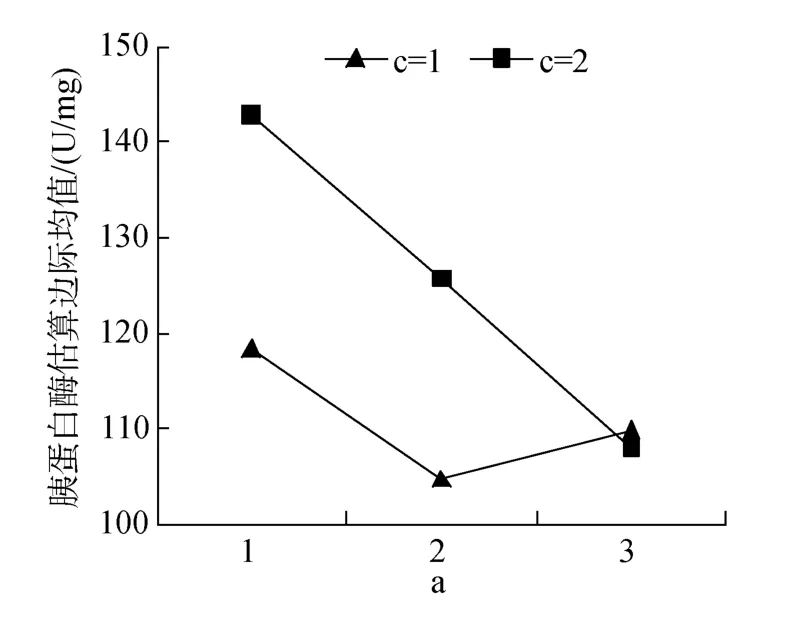
图5 蛋白质和鱼粉互作对胰蛋白酶活性的效应Fig.5 Effects of protein and fish meal interactions on trypsin activity

图6 脂肪和鱼粉互作对胰蛋白酶活性的效应Fig.6 Effects of lipid and fish meal interactions on trypsin activity
2.2.2 肠脂肪酶、淀粉酶
表3中肠脂肪酶和淀粉酶活性结果表明, 脂肪、鱼粉单因素对二者活性均有显著或极显著影响(P<0.05或0.01)。双因素蛋白质和脂肪互作对肠淀粉酶活性, 蛋白质和鱼粉互作对二者活性, 均有显著影响 (P<0.05), P40L10互作和 P43FH 互作效应显著。3因素互作差异极显著(P<0.01), 9#肠脂肪酶活性最高、11#、7#、1#次之; 3#肠淀粉酶活性最高, 1#、9#次之。
2.2.3 肝胰脂肪酶、淀粉酶
表3中肝胰脏脂肪酶和淀粉酶结果表明, 单因素均有极显著影响(P<0.01)。双因素蛋白质和脂肪互作对二者活性均有极显著影响(P<0.01); 蛋白质和鱼粉互作对脂肪酶活性, 脂肪和鱼粉互作对淀粉酶活性均影响极显著(P<0.01)。3因素互作均无显著差异(P>0.05)。
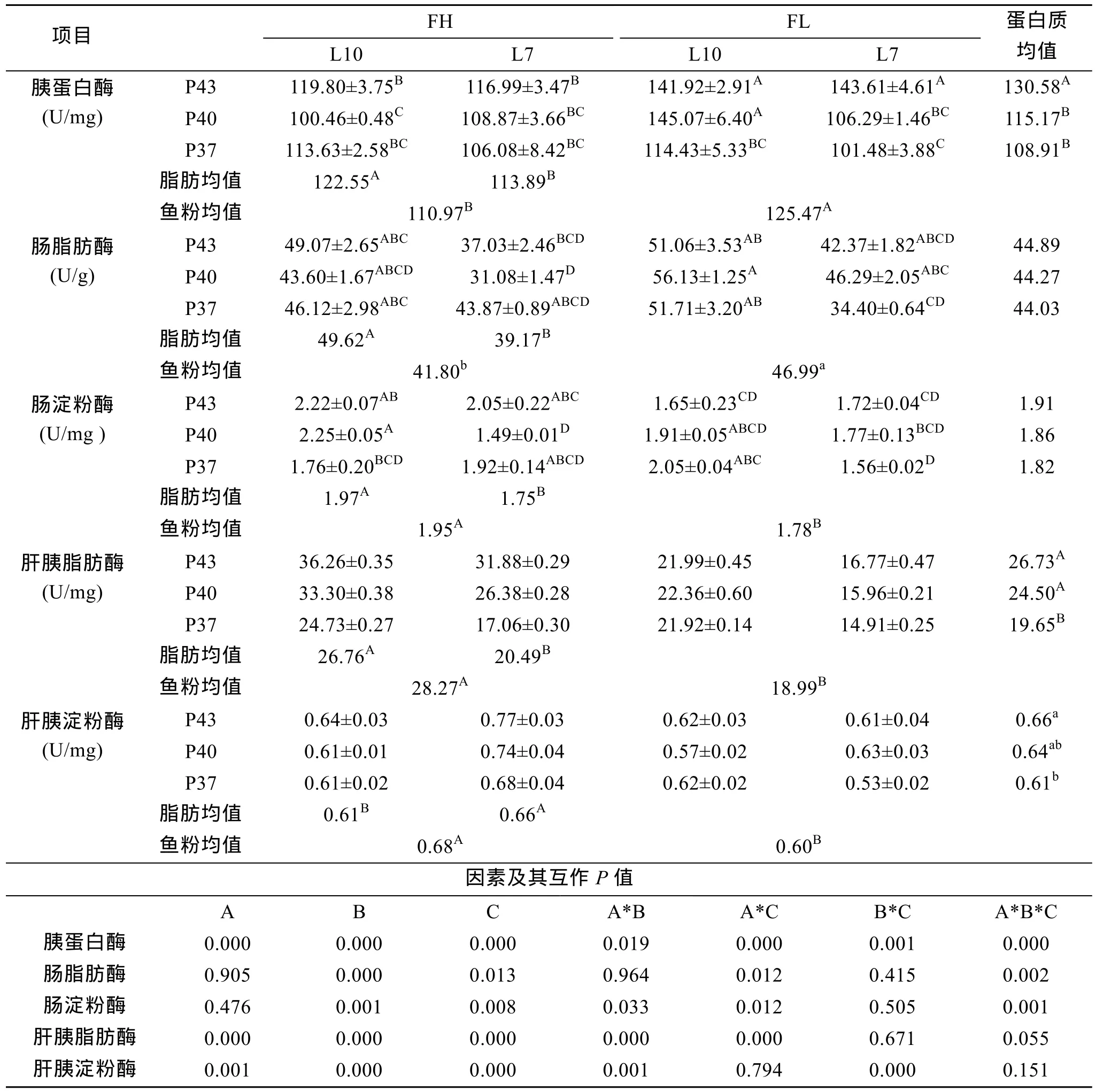
表3 主要营养素源组合对消化酶活性的影响Tab.3 Effects of the main nutritional element and source combination on digestive enzymes activities
2.3 吸收酶活性
12种不同营养素源组合饲粮下, 暗 纹东方 鲀 的吸收酶活性结果见表4。表4中吸收酶活性结果表明,单因素脂肪水平对二者活性均有极显著影响(P<0.01), 蛋白质水平仅对 Na+-K+-ATP酶活性影响极显著。其他单、双、3因素互作均差异不显著(P> 0.05),仍看出1#活性最高, 3#、9#次之, 7#、2#、11#较高。
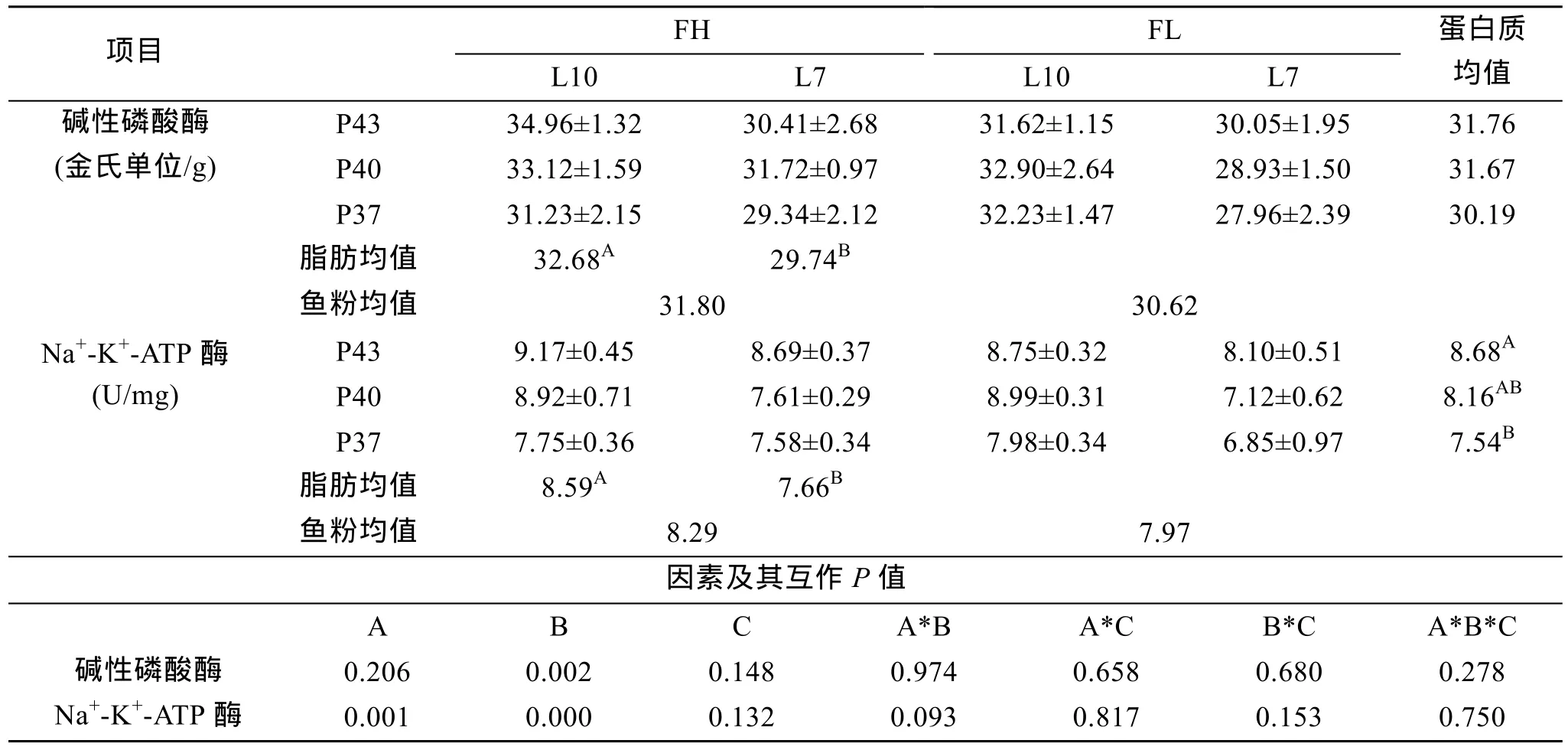
表4 主要营养素源组合对吸收酶活性的影响Tab.4 Effects of the main nutritional element and source combination on absorption enzymes activities
2.4 水环境指标
12种不同营养素源组合饲粮下, 暗纹东方鲀的养殖水环境指标结果见表5。
2.4.1 氨氮
表5中氨氮浓度结果表明, 蛋白质、鱼粉单因素均影响极显著(P<0.01)。双因素蛋白质和脂肪互作差异极显著(图7)。3因素互作差异不显著(P<0.05), 仍看出12#浓度最低, 7#、11#次之。
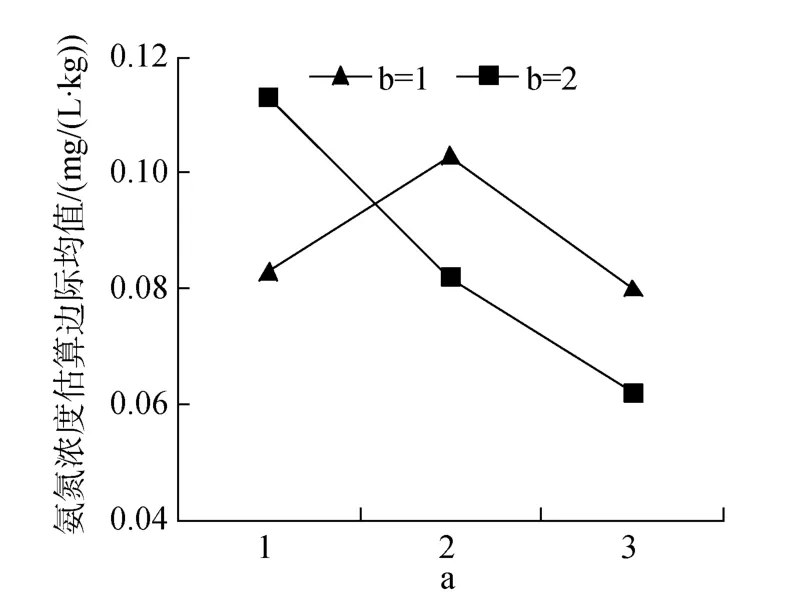
图7 蛋白质和脂肪互作对氨氮浓度的效应Fig.7 Effects of protein and lipid interactions on ammonia nitrogen concentrations
2.4.2 磷酸盐
由表5中磷酸盐浓度结果可见, 脂肪、鱼粉单因素分别影响显著或极显著(P<0.05或0.01)。双因素脂肪和鱼粉互作影响显著(图8)。3因素互作有极显著影响(P<0.01), 11#浓度最低, 与1#、9#无显著差异。
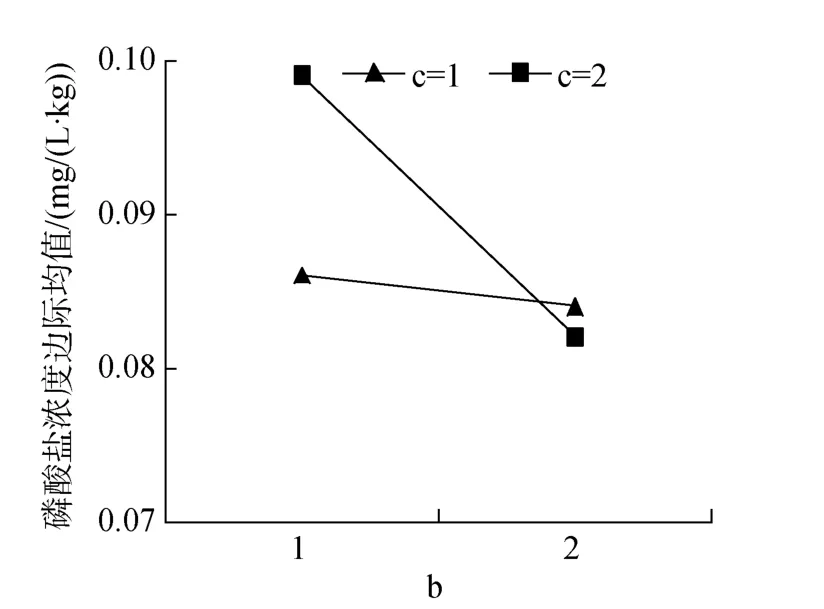
图8 脂肪和鱼粉互作对磷酸盐浓度的效应Fig.8 Effects of lipid and fish meal interactions on phosphate concentrations
2.4.3 亚硝酸氮、COD
由表5可以看出, 仅鱼粉单因素对COD有极显著影响(P<0.01)。双因素脂肪和鱼粉互作、3因素互作显著或极显著影响亚硝酸氮(P<0.05或0.01)。
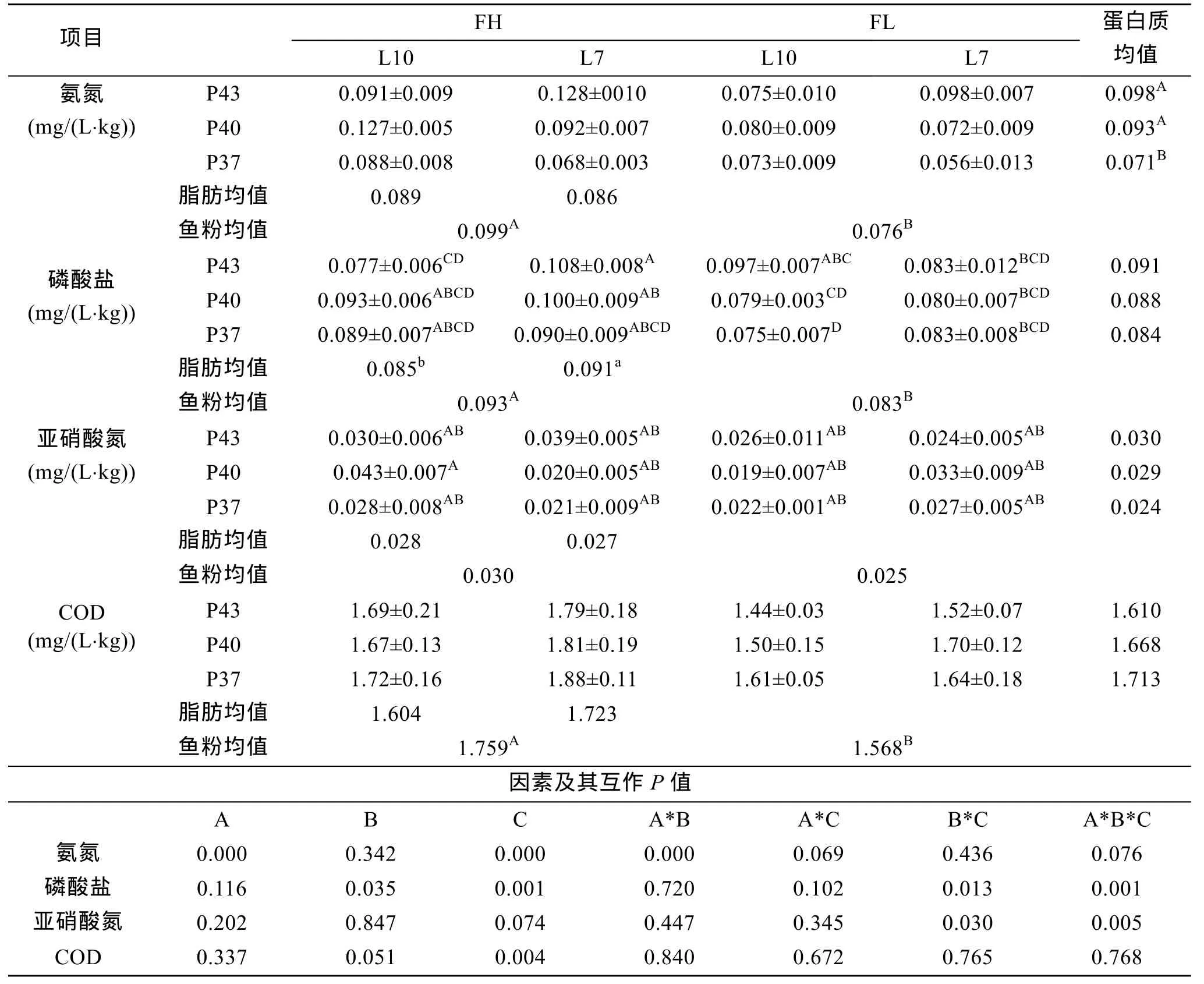
表5 主要营养素源组合对水环境指标的影响Tab.5 Effects of the main nutritional element and source combination on water environmental indicators
3 讨论
3.1 主要营养素源组合对生长和饲料利用的影响
红鳍东方 鲀 的研究表明, 幼稚鱼生长和生理机能最佳时, 蛋白质含量为 50%±3.7%[4]、41.08%[5]和41%[6]。当脂肪含量为 8.93%时, 幼鱼生长代谢机能最佳[8], 且含量小于11%时, 脂肪含量的提高才会增强其生长性能[7]。 暗纹东方 鲀 研究仅见蛋白质含量为46%~49%时, 幼鱼生长和饲料利用最佳[12]。适宜脂肪、鱼粉需求的研究仍未见有。主要营养素源组合对暗纹东方 鲀 的生长效应研究尚属空白。
本研究表明, 1#生长效果最佳, 其增质量率最高,饲料系数最低; 9#次之, 且与1#差异不显著; 3#、11#和7#相对表现也较佳。1#和3#分别显著或极显著改善生长性能, 符合常规, 因其营养素源组合高优。而值得讨论的新发现是, 低鱼粉组9#和11#、7#分别显著或明显改善生长和饲料利用, 营养素源组合正效应突出, 更加节约鱼粉资源和降低饲粮成本, 具有重要理论机制探讨意义和难得经济实用推广价值。
由试验 3因素方差分析知, 单因素蛋白水平对增质量率、鱼粉水平对饲料系数均无显著影响(P>0.05),高脂肪显著改善增重和饲料利用, 中蛋白显著降低饲料系数; 双因素 P40FL改善增质量率和饲料系数的效应均与 P43FH差异不显著, P40L10显著降低饲料系数。故 9#中蛋白高脂肪低鱼粉组合显著改善生长性能的原因包含其中。11#由于替换为低蛋白, 效果显著低于 9#, 但与高鱼粉组 3#无显著差异, 也包含上述单双因素效应的原由。
再从营养学机制和前人研究结果分析, 9#、11#饲粮鱼粉比例较低, 但均为高脂肪水平, 加之补充优质动植物蛋白源(美国肉骨粉、酵母蛋白、大豆蛋白、玉米蛋白等)间及与鱼粉组合后的总体交互作用(氨基酸、脂肪酸、活性物等互补互作)正效应较强,使总体生产性能结果不同程度分别接近 1#、3#; 当提供适量脂肪作为能源时, 蛋白质的需要量可以降低或被节约[14], 也是9#、11#显著改善生长效果的主要原因所在。已有研究表明, 高脂肪有助于红鳍东方鲀幼鱼的生长, 对降低饲料系数有一定影响, 同时可促进鱼体代谢[8]。低鱼粉水平时, 可通过增加油脂使用量, 提高代谢能, 使消化过程中能量消耗减少,热增耗降低, 净能增加, 充分体现脂肪增效作用[15]。
3.2 主要营养素源组合对消化吸收的影响
东方 鲀 消化酶研究, 多见环境因子[16-17]和无机离子[18]对蛋白酶活性的影响, 而脂肪酶、淀粉酶研究甚少。碱性磷酸酶可以协助鱼体肠上皮细胞吸收脂类、葡萄糖、氨基酸等[19], Na+-K+-ATP 酶活性可间接反映小肠黏膜的吸收功能[20], 二者活性研究曾见建鲤幼鱼饲粮蛋白质的影响[21], 而主要营养素源组合对暗纹东方 鲀 的消化吸收性能研究仍属空白。
由本研究消化吸收性能结果可知, 低鱼粉组9#、11#、7#显著或明显提高胰蛋白酶和肠脂肪酶活性,而对碱性磷酸酶和 Na+-K+-ATP 酶活性无显著影响,同时高脂肪具有增强多数消化吸收酶活力的作用。此结果特征与本试验生产性能结果相一致, 可视为低鱼粉组9#、11#、7#和高脂肪因素显著改善生长性能的消化生理原因。
由试验 3因素方差分析得, 单因素高脂肪极显著提高多数消化吸收酶活性; 低鱼粉显著提高胰蛋白酶和肠脂肪酶活性; 高、中蛋白显著提高胰蛋白酶和Na+-K+-ATP酶活性。双因素P40L10显著提高胰蛋白酶活性(且与P43L10无显著差异)和肠、肝胰淀粉酶活性; P40FL显著提高肠脂肪酶活性。两种吸收酶的双因素互作效应不显著, 但高脂肪单因素提高二者活性的效应仍较明显。9#多数消化酶活性强、吸收酶活性改善的原因从而得以解释。7#、11#酶活较高的原因也包含在单双因素效应之中。
增加饲粮脂肪可提高鱼体脂肪酶活力, 反之降低[22]; 低鱼粉组适宜的动植物蛋白比例一定程度的刺激消化酶分泌, 提高各种营养的消化率和蛋白质利用率[23-24]; 且玉米蛋白粉对鱼体消化道生理环境及消化吸收率的干扰较少[24], 加之脂肪的节约效应,表现为低鱼粉组9#消化酶活性显著提高。饲粮蛋白可诱导鱼体蛋白酶分泌, 促进蛋白质的消化与吸收[25],故7#、9#的高、中蛋白水平使得胰蛋白酶活性较高。高、中蛋白水平下Na+-K+-ATP酶活性也较高, 有利于肠道内营养物质的消化后吸收。这一结果特征在豹纹鳃棘鲈(Plectropomus leopardus)也有发现[26]。大菱鲆(Psetta maxima)为适应植物性蛋白源豆粕含量的增加, 蛋白酶活力有增大趋势[27], 也与本研究结果相符。研究还发现, 高鱼粉组其淀粉酶活性较高, 但肉食性鱼类淀粉酶对消化性能的作用微弱[28]。
3.3 主要营养素源组合对养殖水环境的影响
本研究中, 除 COD外, 氨氮、亚硝酸氮、磷酸盐三指标的相对浓度变化呈现出相似趋势或特征,即高鱼粉组及低鱼粉高蛋白组的含量普遍高于其他低鱼粉组, 以11#、9#、7#、1#相对较低; 亚硝酸氮、磷酸盐二指标相对浓度变化出现组间显著差异,9#、11#、1#不同程度显著降低; 另外发现, 高脂肪具有降低水体磷酸盐和COD 的趋势。
由试验 3因素方差分析结果可知, 单因素低鱼粉极显著降低氨氮、磷酸盐、COD浓度, 也有明显降低亚硝酸氮浓度的趋势; 高脂肪显著降低磷酸盐浓度, 且有明显降低 COD的趋势; 高、中蛋白极显著提高氨氮浓度。双因素L10FL显著降低亚硝酸氮浓度, L10与FH、FL互作均降低磷酸盐浓度; P43L10可显著降低氨氮浓度。由此说明11#、9#、7#高脂肪低鱼粉组合显著改善水环境的原因。1#因互作效应显著降低氨氮、磷酸盐浓度, 也使水质明显改善。
由于不同营养水平组合的不同互作产生的正负效应, 导致不同处理组试鱼生长和饲料利用的差异,使除 1#外的其他高鱼粉组的增质量率、饲料系数与低鱼粉组相近, 而前者饲粮均不同程度含有较高质量和数量的氮磷营养(蛋白质、氨基酸、有效磷等), 造成氮磷食入多、消化吸收少、排泄多, 从而导致氨氮、亚硝酸氮、磷酸盐相对浓度较高。已有研究表明[23], 提高饲粮脂肪一定程度降低氮排泄率, 低蛋白水平可显著降低氨氮浓度, 低蛋白和高脂肪互作可有效降低大西洋鲑(Salmo salar)的氨氮排泄量。另有研究证实, 大豆浓缩蛋白和玉米蛋白粉可降低欧洲狼鲈的总磷排放, 增加磷储留[29]; 多种蛋白源混合饲料在不影响鱼生长性能的情况下, 具有较低磷排泄[30], 均与本研究相符;同时, 本试验低鱼粉组饲粮的总磷含量偏低, 也是水体磷酸盐含量低的原因之一。
4 结论
主要营养素源及其互作显著影响暗纹东方 鲀 的生长、消化吸收和水环境。1#、9#生长性能显著提高, 3#、11#和 7#明显提高; 低鱼粉组 9#、11#、7#显著提高胰蛋白酶和肠脂肪酶活性, 高脂肪极显著提高吸收酶活力; 低鱼粉中低蛋白组的氨氮、亚硝酸氮、磷酸盐相对浓度变化呈现普遍低于其他处理组的特征。值得关注的新发现是, 低鱼粉组9#、11#、7#营养素源组合正效应突出, 节约鱼粉资源且降低饲粮成本, 具有重要理论和应用价值。下一步工作需在本研究基础上, 进行效应显著低鱼粉组(9#、11#、7#)的重复试验, 以最终确定其显著效果, 及早应用到生产实践中。
[1] 王奎旗, 陈梅, 高天翔.东方鲀属鱼类的分类与区系分布研究[J].青岛海洋大学学报(自然科学版), 2001,6: 855-860.
[2] Yang Z, Chen Y.Length-weight relationship of obscure puffer(Takifugu obscurus)during spawning migration in the Yangtze River, China[J].Journal of Freshwater Ecology,2003, 18: 349-352.
[3] Takii K, Ukawa M, Nakamura M, et al.Suitable lipid level in brown fish meal diet for tiger puffer [J].Fisheries Science, 1995, 61 (5): 841-844.
[4] 朱钦龙.红 鳍东方 鲀 (雉鱼)饲料的最适蛋白质含量[J].科学饲料, 2003, 3: 41-41.
[5] 王淑敏.红 鳍东方 鲀 幼鱼饲料最适蛋白质含量的研究[D].石家庄: 河北师范大学, 2008.
[6] Kim S S, Lee K J.Dietary protein requirement of juvenile tiger puffer (Takifugu rubripes) [J].Aquaculture,2009, 287: 219-222.
[7] Kotaro, Takeshi, Nakahiro, et al.Effect of dietary lipid levels on the growth, feed utilization, body composition and blood characteristics of tiger pufferTakifugu rubripes[J].Aquaculture, 2009, 298(1): 111-117.
[8] 孙阳, 姜志强, 李艳秋, 等.饲料脂肪水平对红鳍东方 鲀 幼鱼生长、体组成及血液指标的影响[J].天津农学院学报, 2013, 3: 14-18.
[9] Takii K, Ukawa M, Nakamura M, et al.Suitable sugar level in brown fish meal diet for tiger puffer [J].Fisheries Science, 1995, 61(5): 837-840.
[10] Kato K, Ishibashi Y, Murata O N, et al.Qualitative water-soluble vitamin requirements of tiger puffer [J].Fisheries Science, 1994, 60 (5): 589-596.
[11] Jinee E, Lee K J.Effect of dietary ascorbic acid on growth and non-specific immune responses of tiger pufferTakifugu rubripes[J].Fish & Shellfish Immunology, 2008, 25: 611-616.
[12] 杨州, 杨家新.暗 纹东方 鲀 幼鱼对蛋白质的最适需要量[J].水产学报, 2003, 5: 450-455.
[13] 刘襄河, 叶超霞, 郑丽勉, 等.饲料糊精水平对暗纹东方 鲀 幼鱼生长、消化酶活性和血液生化指标的影响[J].水产学报, 2013, 9: 1359-1368.
[14] National Research Council (NRC).Nutrient requirements of fish [M].Washington, DC: National Academy Press, 2011.
[15] 叶元土, 蔡春芳.鱼类营养与饲料配制[M].北京:化学工业出版社, 2013: 235.
[16] 赵清良, 殷宁, 王培军.pH 对 暗纹东方 鲀 仔稚鱼蛋白活性的影响[J].水利渔业, 2002, 5: 24-25.
[17] 万蓁蓁, 高天翔, 张秀梅.pH 对 红鳍东方 鲀 幼鱼蛋白酶活性的影响[J].齐鲁渔业, 2009, 11: 7-10, 5.
[18] 殷宁, 赵强, 李朝晖, 等.暗 纹东方 鲀 蛋白酶活性的研究[J].南京师大学报(自然科学版), 2001, 1:101-104.
[19] Villanueva J, Vanacore R, Goico, et al.Intestinal alkaline phosphatase of the fishCyprinus carpio: regional distribution and membrane association[J].Journal of Experimental Zoology, 1997, 279(4): 347-355.
[20] Shils M E, Shike M, Ross A C, et al.Modern nutrition in health and disease.10thed[M].Philadephia: Lippincott Williams & Wilkins, 2006: 1132-1139.
[21] 刘勇.蛋白质对幼建鲤生长性能、消化功能和蛋白质代谢的影响[D].四川: 四川农业大学, 2008: 40-46.
[22] 柳阳, 李勇, 周邦维, 等.脂肪和蛋白质水平对工业养殖大西洋鲑消化酶、非特异性免疫及水质的影响[J].水产学报, 2014, 3: 433-442.
[23] 向枭, 周兴华, 陈建, 等.饲料蛋白水平及鱼粉蛋白含量对齐口裂腹鱼生长、体组成及消化酶活性的影响[J].中国粮油学报, 2012, 5: 74-80, 106.
[24] 陈然, 华雪铭, 黄旭雄, 等.玉米蛋白粉替代鱼粉对异育银鲫生长、蛋白酶活性及表观消化率的影响[J].上海交通大学学报(农业科学版), 2009, 4: 358-362, 367.
[25] 钱曦, 王桂芹, 周洪琪, 等.饲料蛋白水平及豆粕替代鱼粉比例对翘嘴红 鲌 消化酶活性的影响[J].动物营养学报, 2007, 2: 182-187.
[26] 周邦维, 李勇, 高婷婷, 等.主要营养素源对工业化养殖豹纹鳃棘鲈生长、体色和消化吸收的影响[J].动物营养学报, 2014, 26(5): 1387-1401.
[27] 王海英, 孙谧, 薛长湖, 等.大菱鲆配合饲料中植物蛋白替代鱼粉的可行性研究[J].海洋科学, 2008,32(6): 9-12, 34.
[28] Wilson R P.Utilization of dietary carbohydrate by fish [J].Aquaculture, 1994, 124: 67-80.
[29] Ballestrazzi R, Lanari D, Dagaro E, et al.The effect of dietary protein level and source on growth, body composition, total ammonia and reactive phosphate excretion of growing sea bass (Dicentrarchus labrax) [J].Aquaculture, 1994, 127 (2-3): 197-206.
[30] 陆静, 邵庆均.水产动物对饲料中磷的利用研究进展[J].中国饲料, 2013, 1: 34-38.
