沙棘液流动态变化及影响因素分析
2015-04-11罗凤敏辛智鸣高君亮黄雅茹葛根巴图李新乐郝玉光
罗凤敏,辛智鸣,高君亮,黄雅茹,葛根巴图,李新乐,郝玉光
(国家林业局内蒙古磴口荒漠生态系统定位观测研究站/中国林业科学研究院沙漠林业实验中心,内蒙古 磴口015200)
沙棘(Hippophae rthamnoides)是胡颓子科(Elaeagnaceae)沙棘属的灌木,具有耐旱、耐盐碱[1]及抗风沙等优点,是西北地区重要的水土保持、防风固沙树种,同时也是重要的薪炭和饲料树种。乌兰布和沙漠位于我国干旱荒漠区,生态环境脆弱,蒸发强烈且降水量极少的自然条件严重影响该区植被存活及生长,部分呈现衰退甚至死亡。
干旱区植被适应干旱的主要途径是减少体内水分的消耗,而干旱区植物茎干液流变化特征在反映植物对干旱环境适应机制方面具有的重要作用。同一种植物其液流速率在不同地区有显著差异。如:在毛乌素沙地,柠条锦鸡儿(Caragana korshinskii)的茎干液流速率日变化为多峰曲线,其变化特征与环境因子大致相似[2-3];而在沙坡头,柠条的液流速率则表现为单峰曲线[4]。研究直径较小的植物液流时,包裹式茎流探头有很大的优越性,经过一些学者的完善[5-7],然后经过试验验证其准确性很高。有学者采用热平衡包裹式树干液流仪对沙冬青(Ammopiptanthus mongolicus)及沙木蓼(Atraphaxis bracteata)液流的连续监测结果表明,二者具有明显的昼夜节律,在植株生长季,沙木蓼液流总量与基径呈正比[8-9]。对梭梭(Haloxylon ammodendron)、多枝柽柳(Tamarix ramosissima)和唐古特白刺(Nitraria tangutorum)的液流检测结果表明,3 种灌木茎干液流季节变化为单峰型,环境因子对其液流速率影响显著[10]。六盘山生长的沙棘液流与太阳辐射等环境因子具有显著相关性[11]。余峰等[12]通过连续观测柠条锦鸡儿液流速率变化特征,并查明了其与环境因子的关系。
沙棘作为乌兰布和沙漠典型的人工灌木树种之一,研究其液流变化可为在该区域水分平衡条件下的人工沙棘林培育和管理提供一定的科学依据。基于此,本研究对乌兰布和沙漠人工沙棘林的沙棘植株茎干液流变化进程进行昼夜连续观测,同时,借助生态站同步记录的气象因子数据,查明沙棘液流变化特征及其对气象因子的响应,以期为该区域植物水分利用过程的研究及植物资源的利用提供参考,同时也为沙区气候对灌木的影响提供科学依据。
1 试验设计与研究方法
1.1 研究区概况
研究区选择在位于乌兰布和沙漠东北部的国家林业局内蒙古磴口荒漠生态系统定位研究站。据该生态站多年气象数据显示,该区域多年平均气温7.5 ~8.5 ℃,多年平均日照时数3 100 ~3 300 h,多年平均风速3 ~3.7 m·s-1[13]。天然植被以抗旱能力较强的荒漠植被为主。
1.2 研究方法
为了排除土壤环境因子差异所产生的影响,本研究选择生长在相同立地条件下的沙棘作为样株。将距样株基部1.0 m 处的树皮刮掉,露出韧皮部(10 cm×20 cm)。然后安装测定探针,安装完后,将数据采集器(DT80)与TDP 馈线相连接后通电,连接笔记本电脑与数据采集器设置工作参数,并输入相应茎干面积值,采集时间间隔为10 min,定期采集数据,对沙棘液流速率进行连续观测。在测定沙棘液流的时期内,统计同时期生态站自动气象站所记录的地面2 m 内太阳总辐射、空气相对湿度、风速、气温及10 cm 处土温,时间间隔与茎流采集间隔一致。本研究采用的茎流测量系统探头型号、工作参数及沙棘样株特征如表1 所示。

表1 探头型号、工作参数及样株特征Table 1 Specifications and working parameters and the properties of the measured samples
2 结果与分析
2.1 沙棘茎干液流连日变化规律
由基径10.5 cm 的沙棘茎干液流在9 月3 -7日的变化曲线(图1)可知,9 月上旬沙棘液流昼夜变化规律明显,白天沙棘茎干液流速率的变化特征为多峰曲线,且流速较高,但夜间流速则显著降低,且波动范围小。研究区9 月3 -7 日均是晴朗天气,沙棘液流启动时间为07:50,流速上升速度显著增加,流速峰值出现在09:10 左右,9 月3 -7 日的液流峰值分别是322.47、292.07、294.61、340. 27 和295.85 g·h-1,差异不明显。在08:50 -18:40,液流速率较大,其液流量大于全天的80%;至18:40之后沙棘液流速率的明显减小,至21:30 左右,沙棘植株茎干液流速率变化曲线逐渐趋于平缓,并且降到极低值;但是,液流在夜间并没有停止,只是流速变得很小而已,其值维持在2.83 ~16.98 g·h-1,最小值出现在00:00 -07:00。乌兰布和沙区白天气温高,夜晚气温低,昼夜温差较大,且空气相对湿度较低。白天空气温度高,蒸腾作用强,夜晚沙棘叶片气孔关闭,蒸腾作用也停止,但是在根压作用下,沙棘根系仍然能够继续吸水,补充白天因蒸腾作用散失的水分,并且使其体内水分平衡恢复[14]。一个日周期的沙棘植株茎干液流的活跃时间约为13 h。
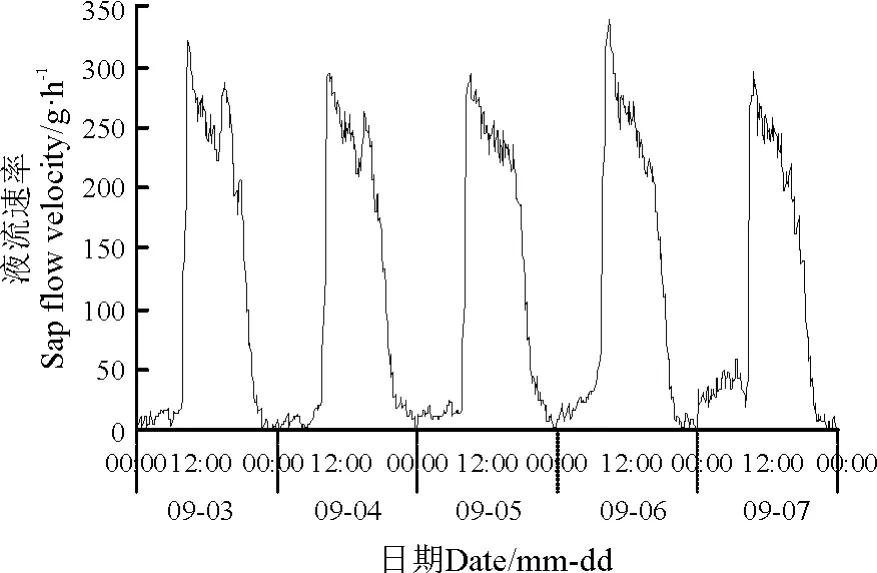
图1 沙棘茎干液流连日进程Fig.1 Diurnal course of sap flow velocity in Hippophae rhamnoides
2.2 不同基径沙棘茎干液流日变化特征
不同基径的沙棘,其液流速率变化特征及日累积量有很大差异(图2 和3)。3 个不同基径的沙棘液流速率不同,流速从大到小依次为:基径10.5 cm的沙棘,其次为基径8.3 cm 的沙棘,基径7.5 cm 的最小(图2),正如一些学者的研究结果表明,植物液流速率与其茎干直径大小之间为正相关关系[15-16]。3 个不同基径沙棘植株的液流速度在夜晚逐渐一致,差异较小。白天,基径10.5 cm 沙棘的液流峰值为291.80 g·h-1,基径8.3 cm 沙棘的液流峰值为281.11 g·h-1,基径7. 5 cm 沙棘的液流峰值为268.82 g·h-1,最大值是最小值的1.09 倍。在时间及环境因子一致的条件下,沙棘植株的茎干直径虽然不同,但是其液流速率变化趋势基本相同,在沙棘植株液流速率维持在较高值的阶段,其液流变化曲线波动也较大。分析以上结果可知,沙棘植株茎干液流速率越大,受环境影响的程度越大。这与沙拐枣(Calligonum mongolicum)、柠条锦鸡儿及多枝柽柳等植物茎干液流变化规律相似[15]。沙棘液流累计曲线符合典型生物生长曲线,基径为10.5、8.3和7.5 cm 的沙棘日累积液流量分别为16 593.26、14 907.08和14 101.83 g,即沙棘耗水量随其基径增加而增大,这也表明随着沙棘植株生长,其耗水量在增加。
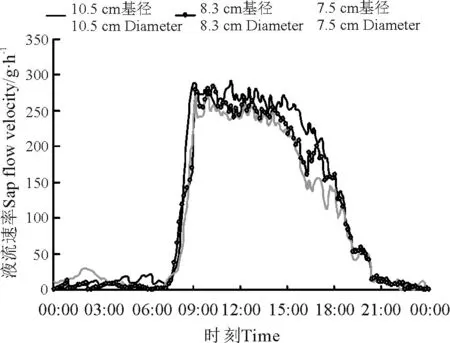
图2 3 种基径沙棘液流速率日变化特征Fig.2 Daily variation characteristics of sap flow velocity in Hippophae rhamnoides with three diameters
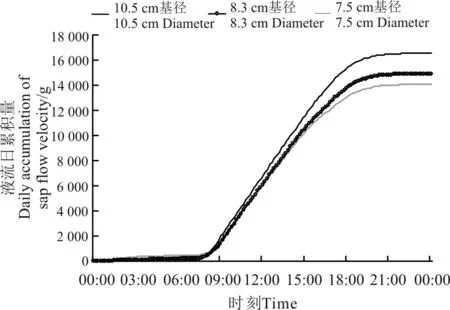
图3 3 种基径沙棘液流速率日累积量Fig.3 Daily accumulation of sap flow velocity in Hippophae rhamnoides with three diameters
2.3 沙棘植株液流与环境因子的关系
环境因子对其液流变化特征起着决定性的作用。本研究结果显示,同步测定的环境因子也具有明显的昼夜节律性,沙棘茎干液流速率变化趋势与同步观测的气温、太阳总辐射、风速以及10 cm 处土层土温变化趋势大致一致,但是与空气相对湿度的变化趋势相反(图4 -8)。
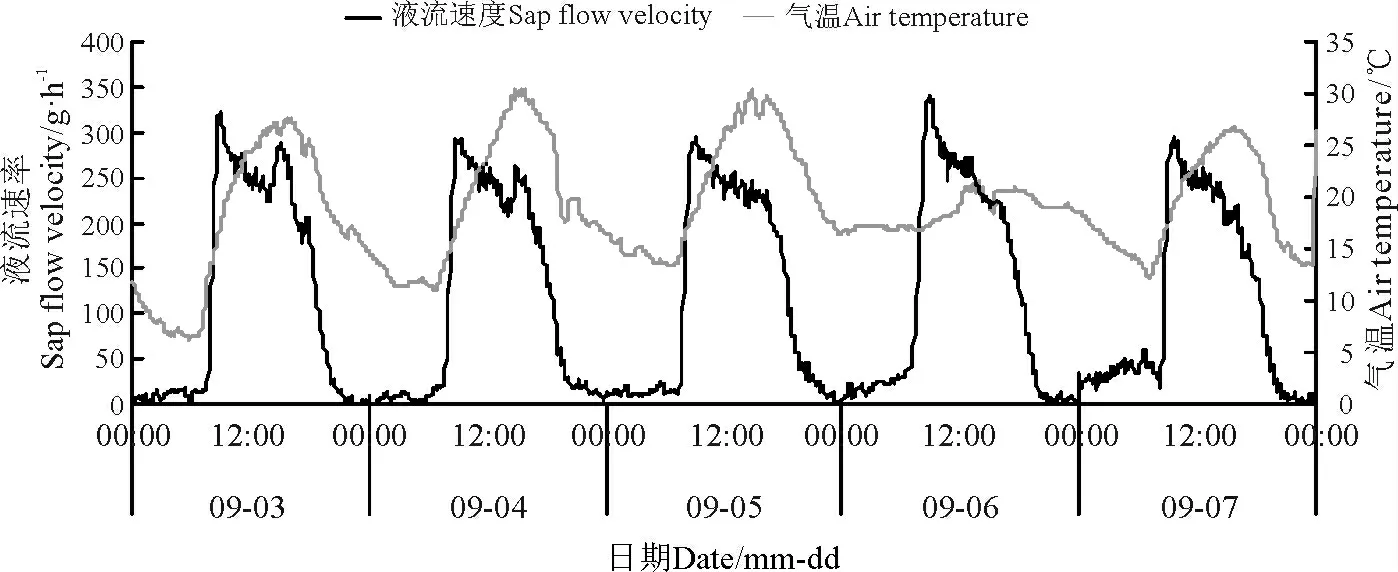
图4 液流速率与气温日进程Fig.4 Daily course of sap flow rate and air temperature
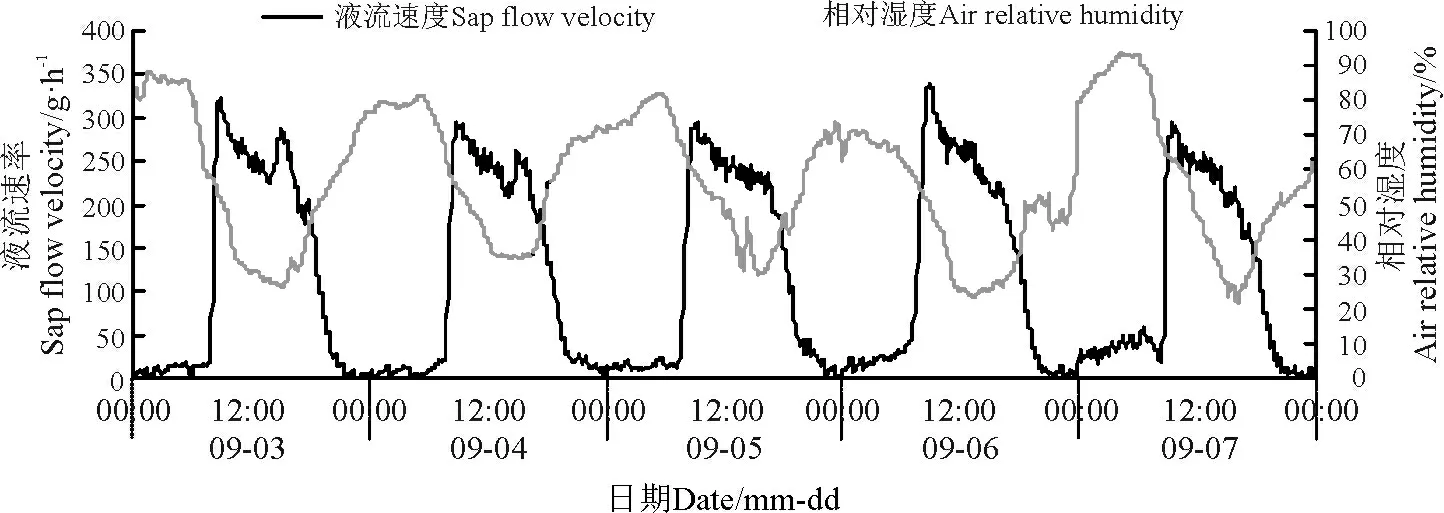
图5 液流速率与相对湿度日进程Fig.5 Daily course of sap flow velocity and air relative humidity
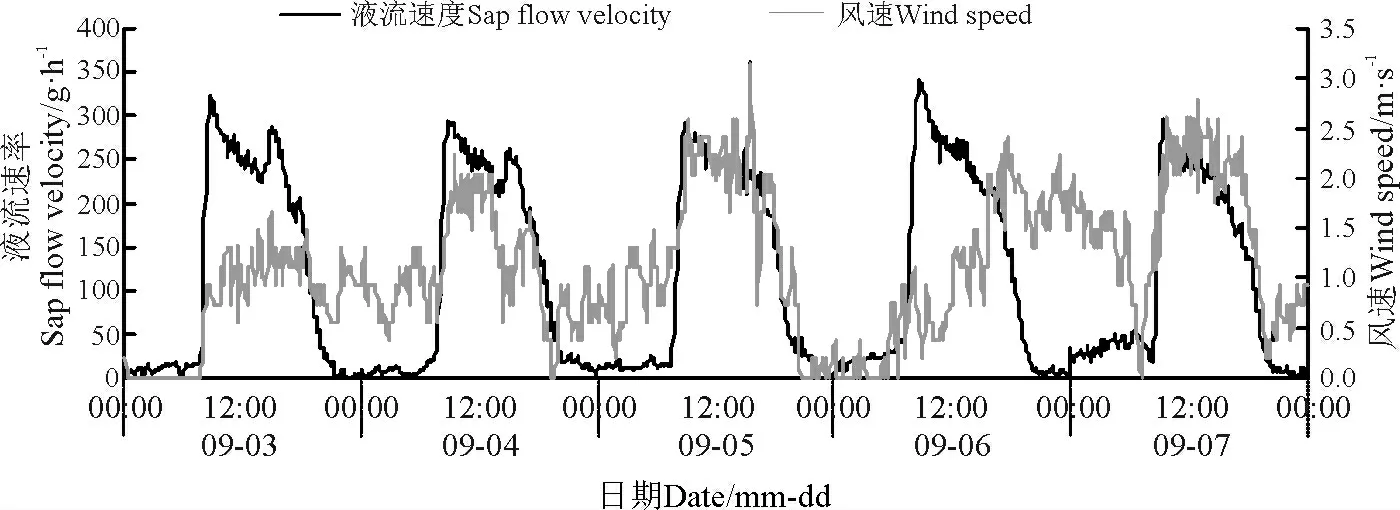
图6 液流速率与太阳总辐射日进程Fig.6 Daily course of sap flow rate and total solar radiation
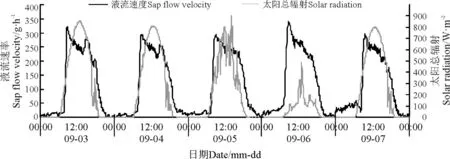
图7 液流速率与风速日进程Fig.7 Daily course of sap flow velocity and wind speed
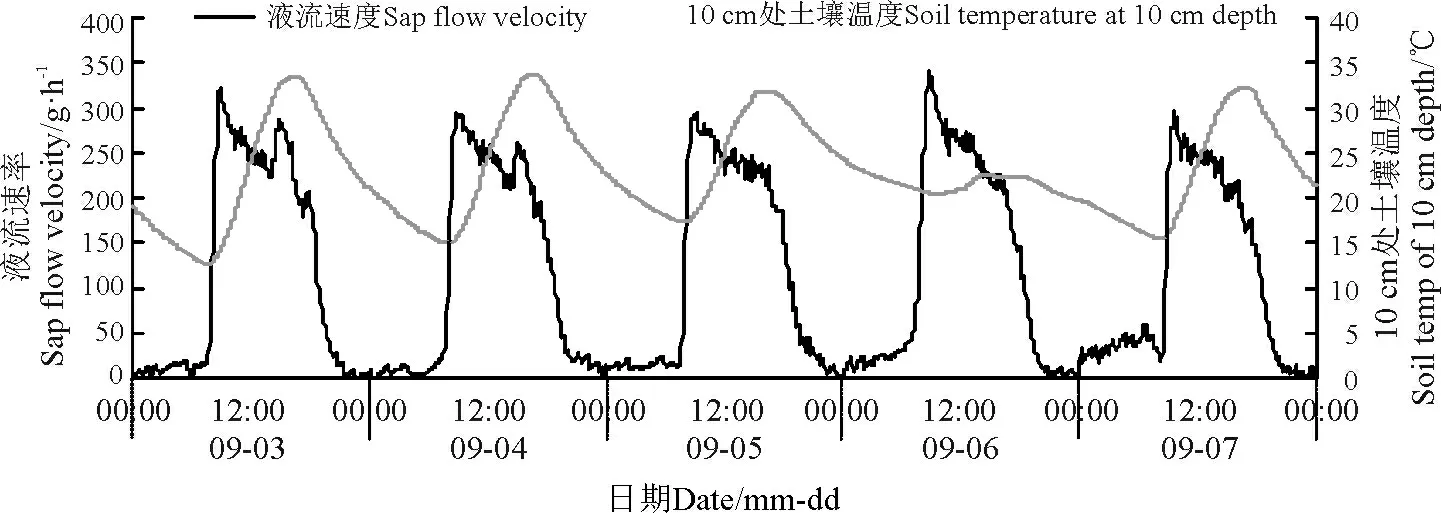
图8 液流速率与10 cm 处土壤温度日进程Fig.8 Daily course of sap flow velocity and soil temperature at depth of 10 cm
太阳辐射强度对植物液流具有直接影响,与此同时对气温及相对湿度也具有明显的影响。在晴朗的天气条件下,研究区自06:30 左右开始,太阳辐射强度逐渐增强(图4,图6),沙棘植株茎干液流也开始启动,但是沙棘液流启动时间却比太阳辐射开始时间滞后约1 h。随着太阳辐射强度逐渐增强,气温升高,沙棘植株茎干液流速率不断增加并快速达到高峰期。在高峰期内,沙棘茎干液流变化趋势为多峰曲线,产生这种现象的原因是太阳辐射强度的变化致使沙棘蒸腾速率发生波动。当植物根系吸收的水分不足以供应蒸腾作用所耗用的水分时,植物就会通过关闭其叶片的部分气孔进行调节,蒸腾速率也会逐渐减小,因为蒸腾拉力是植株茎流的主要动力,因此沙棘植株茎干液流速率降低。而当太阳辐射强度逐渐降低时,空气温度相应地降低,由此导致沙棘植株茎干液流速率减小,在21:00 左右降到最小值。晚间空气温度逐渐降低,因此,沙棘植株液流能够在一段时间内保持最低值。
9 月3 -7 日的风速变化规律均是白天大而夜间小,风速变化规律与沙棘液流变化趋势基本吻合(图7)。风速增加会使蒸腾速率变大;大风会导致气孔开度降低,甚至关闭[17-18]。蒸腾拉力对沙棘茎干液流速率的增大具有促进作用,所以风速大小也会对其液流产生一定程度的影响。此外,土壤具有良好的热容性和传导阻力特性,导致10 cm 处土层土温波动规律滞后于气温波动变化[19],与此同时,10 cm 处土温波动规律也滞后于液流速率,10 cm 处土壤温度下降速度较慢,在日出之前达到最小值(图8)。
对沙棘液流速率与气温、空气相对湿度、太阳总辐射、风速及10 cm 处土温进行相关性分析表明(表2),沙棘植株茎干液流与气温、空气相对湿度、太阳总辐射、风速及10 cm 处土温呈显著相关关系。其中,沙棘植株液流与太阳总辐射相关性最大(r =0.799),然后依次为空气相对湿度(r = -0.739)、气温(r=0.661)、风速(r=0.541)、10 cm 处土温(r=0.308)。
采用多元逐步回归分析方法,以5%的可靠性作为变量入选和剔除临界值,建立沙棘植株液流和环境因子的回归模型:
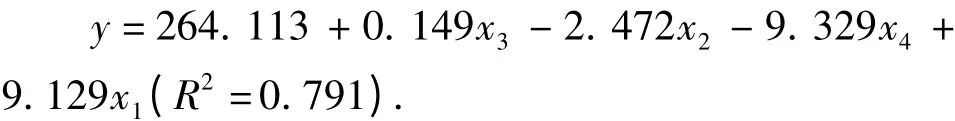
式中,y 为沙棘茎干液流(g·h-1);x1为气温(℃);x2为空气相对湿度(%);x3为太阳总辐射(W·m-2);x4为10 cm 处土温。经检验,该模型达到极显著水平(F=545.57,P=0.000)。
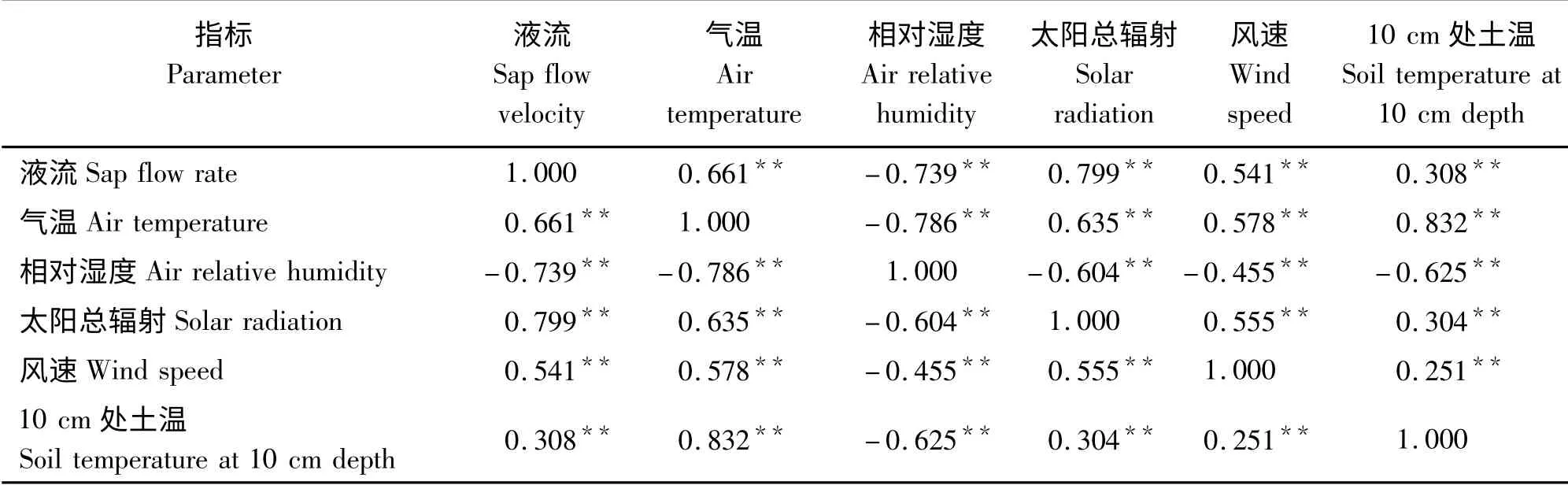
表2 沙棘茎干液流速率与环境因子的相关性分析Table 2 Correlation coefficients between sap flow velocity of Hippophae rhamnoides and environmental factors
3 讨论
同其他植物一样,沙棘的气孔在夜间时完全关闭,但水分也会散失,究其原因,可能是沙棘在夜间具有明显的茎干液流活动。这同前人研究结果相一致,如:六盘山生长的沙棘夜间存在液流运动[12],锦鸡儿属植物在夜间也存在液流运动[2,20]。这可能是因为,北方干旱区的植物白天蒸腾强烈,通常植物处于过度失水状态,只有补充了缺失的水分,才能使其因为过度蒸腾作用所导致受损细胞的生理机能恢复,因此,植物需要在夜间保持液流活动,来增强其耐旱性。
植物生物学结构、气象因素以及土壤水分这3个因素对植物茎干液流的变化趋势起着决定性的作用[21],植物液流的潜在能力由其自身的生物学特性决定,而土壤水分则决定了植物液流的总体水平,气象因素则是植物茎干液流瞬间变化的决定性因子[22-23]。环境因子对植物液流速率的影响作用很大,在地区、植物不同的情况下,环境因子对植物液流速率影响也存在不同程度差异。前人研究表明,气温、太阳总辐射、空气相对湿度及土温这4 个气象因子对植物茎干液流均具有显著影响[22,24-25]。沙棘液流速率连日变化及日变化与气象因子(除相对湿度外)变化规律基本一致,此结果说明环境因子对沙棘液流速率变化特征有明显的影响。
植物由于蒸腾作用所消耗水分的99.8%左右均由茎干液流供给,所以常采用茎干液流量表示植物的蒸腾耗水[26]。因此,研究干旱区灌木茎干液流变化规律可为区域造林树种的选择提供一定的理论依据;同时还可以根据区域水分状况,并结合蒸散量选择适合当地条件的造林密度及人工补充水分总量。
4 结论
1)乌兰布和沙区沙棘茎干液流日变化曲线为宽峰多峰线型,具有显著的昼高夜低的变化趋势,沙棘液流速率在白天时较高,且波动明显,而夜间液流速率相对较低,且变化缓慢;沙棘液流在07:50 左右开始启动,至09:10 时达到峰值,且在09:10 -18:40 之间,流速始终保持较高状态,这期间的液流量大于全天总量的80%;至21:30 左右,液流速率逐渐降低,最小值出现在00:00 -07:00 之间。
2)沙棘茎干液流与其茎干直径呈正相关,直径较大者耗水量较大。茎干直径10.5、8.3 和7.5 cm的沙棘,其日累积液流量分别为16 593. 26、14 907.08和14 101.83 g。
3)沙棘植株液流与太阳总辐射相关性最大(r=0.799),然后依次为空气相对湿度(r = -0.739)、气温(r=0.661)、风速(r=0.541)、10 cm 处土温(r=0.308)。沙棘液流与环境因子之间的关系可用多元回归方程y =264. 113 +0. 149x3-2.472x2-9.329x4+9.129x1(R2=0.791)来表达。
[1] 徐文远,王晓春,穆丽蔷,姜雪昊. 齐泰高速公路5 种护坡灌木对碱胁迫的生理响应[J]. 草业科学,2014,31(5):850-857.
[2] 赵奎,丁国栋,吴斌,张宇清,段玉玺,张静虎,孙毅.宁夏盐池毛乌素沙地柠条锦鸡儿茎流及蒸腾特征[J].干旱区研究,2009,26(3):390-395.
[3] 郭跃.毛乌素沙地沙木蓼和花棒耗水特性研究[D].北京:北京林业大学硕士学位论文,2011.
[4] 黄磊,张志山.荒漠人工植被区柠条和油蒿茎干液流动态研究[J].中国沙漠,2011,30(3):683-688.
[5] Sakuratani T.Improvement of the probe for measuring water flow rate in intact plants with the stem heat balance method[J].Journal of Agricultural Meteorology,1984,40:273-277.
[6] Baker J M,van Bavel C H M.Measurement of mass flow of water in the stems of herbaceous plants[J].Plant,Cell and Environment,1987,10:777-782.
[7] Steinberg S L,van Bavel C H M,McFarland M J.A gauge to measure mass flow rate of sap in stems and trunks of woody plants[J].Journal of the American Society for Horticultural Science,1989,114(3):466-472.
[8] 郭树江,徐先英,杨自辉,李爱德,李得禄,柴成武,王多泽.干旱荒漠区沙冬青茎干液流变化特征及其与气象因子的关系[J].西北植物学报,2011,31(5):1003-1010.
[9] 樊文会,丁国栋,吴斌,张宇清,乌日娜.沙木蓼蒸腾耗水规律研究[J].干旱区资源与环境,2012,26(11):188-193.
[10] 徐先英,孙保平,丁国栋,郭树江,柴成武.干旱荒漠区典型固沙灌木液流动态变化及其对环境因子的响应[J].生态学报,2008,28(3):895-905.
[11] 沈振西,徐丽宏,王彦辉,于澎涛,熊伟,左海军,曹恭祥,王云霓.宁夏六盘山沙棘液流变化及耗水特性[J].中国水土保持科学,2014,12(3):59-65.
[12] 余峰,陈林,王峰,刘华,杜建民,郭永忠,李浩霞.宁夏中部干旱带人工柠条茎流及蒸腾特征研究[J].水土保持研究,2011,18(2):143-147.
[13] 高君亮,辛智鸣,刘芳,原伟杰,冯伟,徐军,赵英铭,郝玉光.乌兰布和沙漠东北部大气降尘特征及影响因素分析[J].干旱区资源与环境,2014,28(8):145-150.
[14] 王华田,马履一,孙鹏森.油松、侧柏深秋边材木质部液流变化规律的研究[J].林业科学,2002,38(5):31-37.
[15] 许浩,张希明,王永东,魏疆,梁少民.塔里木沙漠公路防护林乔木状沙拐枣耗水特性[J].干旱区研究,2006,23(2):216-222.
[16] 辛智鸣,黄雅茹,罗凤敏,葛根巴图,李新乐,孙非,郝玉光.沙棘果期茎干液流变化特征及其与气象因子的关系[J].干旱区资源与环境,2015(11):202-207.
[17] 潘瑞炽.植物生理学[M].北京:高等教育出版社,2001:16-20.
[18] 王瑞辉,马履一,李丽萍,樊敏,孔俊杰.元宝枫树干液流的时空变异性研究[J].北京林业大学学报,2006(S2):12-18.
[19] 马长明,管伟,叶兵,袁玉欣,王金凤,李淑梅.利用热扩散式边材液流探针(TDP)对山杨树干液流的研究[J].河北农业大学学报,2005,28(1):39-43.
[20] 岳广阳,赵哈林,张铜会,云建英,牛丽,何玉惠.不同天气条件下小叶锦鸡儿茎流及耗水特性[J].应用生态学报,2007,18(10):2173-2178.
[21] 孙鹏森,马履一,王小平,翟明普.油松树干液流的时空变异性研究[J].北京林业大学学报,2000(5):1-6.
[22] 于红博,杨劼,臧春鑫,徐延达. 皇甫川流域中国沙棘树干液流日变化及其相关因子[J]. 生态学杂志,2008,27(7):1071-1076.
[23] 唐霞,崔建垣,赵学勇,云建英,连杰,王新源,李衍青.科尔沁沙地玉米叶面蒸腾与棵间蒸发特性[J].草业科学,2011,28(5):788-792.
[24] 潘占兵,蒋齐,郭永忠,温学飞,左忠.柠条蒸腾特征及影响因子的研究[J].中国生态农业学报,2006,14(2):70-71.
[25] 张宁南,徐大平,Morris J,周光益,周国逸,吴仲民. 雷州半岛尾叶桉人工林树液茎流特征的研究[J]. 林业科学研究,2003,16(6):661-667.
[26] Granier A,Bobay V,Gash J H C.Vapour flux density and transpiration rate comparisons in a stand of maritime pine(Pinus pinaster Ani.)in Les Landed forest[J].Agricultural and Forest Meteorology,1990,51(3-4):309-319.
