松嫩盐碱草地26 种植物根围丛枝菌根真菌多样性特征
2015-04-11杨春雪岳英男阎秀峰
杨春雪,陈 飞,岳英男,阎秀峰
(1.东北林业大学园林学院,黑龙江 哈尔滨150040;2.东北林业大学盐碱地生物资源环境研究中心,东北油田盐碱植被恢复与重建教育部重点实验室,黑龙江 哈尔滨150040)
土壤盐渍化是世界各国面临的共同难题,我国盐碱土面积已超过了370 万hm2,且正以每年1.4%的速度扩展[1],松嫩平原地处半湿润半干旱过渡带,因溶解在水中的盐分容易在土壤表层积聚,盐碱地主要成分是苏打盐,草甸土和盐碱常呈复合体形式分布[2-3]。现今,针对草地大面积盐碱化、退化等问题,我国各级部门先后采取过以稻治碱、以水治碱等治理措施,尽管投入了大量资金,但土壤盐碱化始终没有得到治理。因此,借助其他技术手段加以辅助和治理恢复草原生态系统就显得尤为重要。
丛枝菌根(Arbuscular Mycorrhizae,AM)真菌是土壤中广泛存在的一类共生真菌,能与90%以上的高等植物形成丛枝菌根[4]。植物长期生活在高盐条件下会出现营养亏缺、离子毒害及氧化胁迫等症状[5],AM 真菌能活化土壤,调节植物渗透平衡,增强植物对矿质元素和水分的吸收,提高光合作用,促进生物量的积累,提高植物的耐盐能力[6-7]。本研究探究松嫩盐碱草地中AM 真菌的资源分布和多样性,分析土壤因子与AM 真菌生态分布和多样性之间的关系,以期为筛选高效AM 真菌菌种资源,促进AM 真菌在盐碱土改良中的应用提供参考。
1 材料与方法
1.1 样品采集
本研究于2013 年7 月在黑龙江省肇东市周边松嫩盐碱草地采集了共11 科26 种耐盐植物根际土壤,样点分布见表1。根据“多点平行取样法”和“五点取样法”[8]按照东南西北4 个方向随机对26 种植物取样,重复3 次,用小铲轻轻刮去地表的杂物,向下垂直挖取12 ~20 cm 深的植物根围土壤,并保留粘附在植物根系表面的细土,最后经“四分法”[8]保留1 kg 土样,统一编号后带回,并记录采集时间、地点和周围环境。
1.2 AM 真菌的分离和鉴定
采集的土壤样品自然风干,每次称取土样100 g,利用湿筛倾析-蔗糖离心法分离AM 真菌孢子[9],于实体显微镜下统计孢子密度、物种丰度和Shannon-Weiner 指数(H);轻挑孢子于载玻片上,分别用水、乳酸、PVLG、Melzer’s 试剂为浮载剂制片,在OLYMPUS CX21 生物显微镜下观察孢子的颜色、纹饰、连孢菌丝等特征,测量孢子大小、菌丝壁厚度、菌丝宽度,拍照并记录。参照“VA 菌根鉴定手册”[10]和国际AM 真菌菌种保藏中心(International Collection Center of Vesicular and Arberscular Mycorrhizal Fungi,INVAM)提供的AM真菌种属描述以及Internet(http://invam. caf. wvu.edu)上提供的图片,并参阅近几年发表的有关AM 真菌分类的相关资料和原始文献对AM 真菌进行形态鉴定。
1.3 土壤理化性质的测定
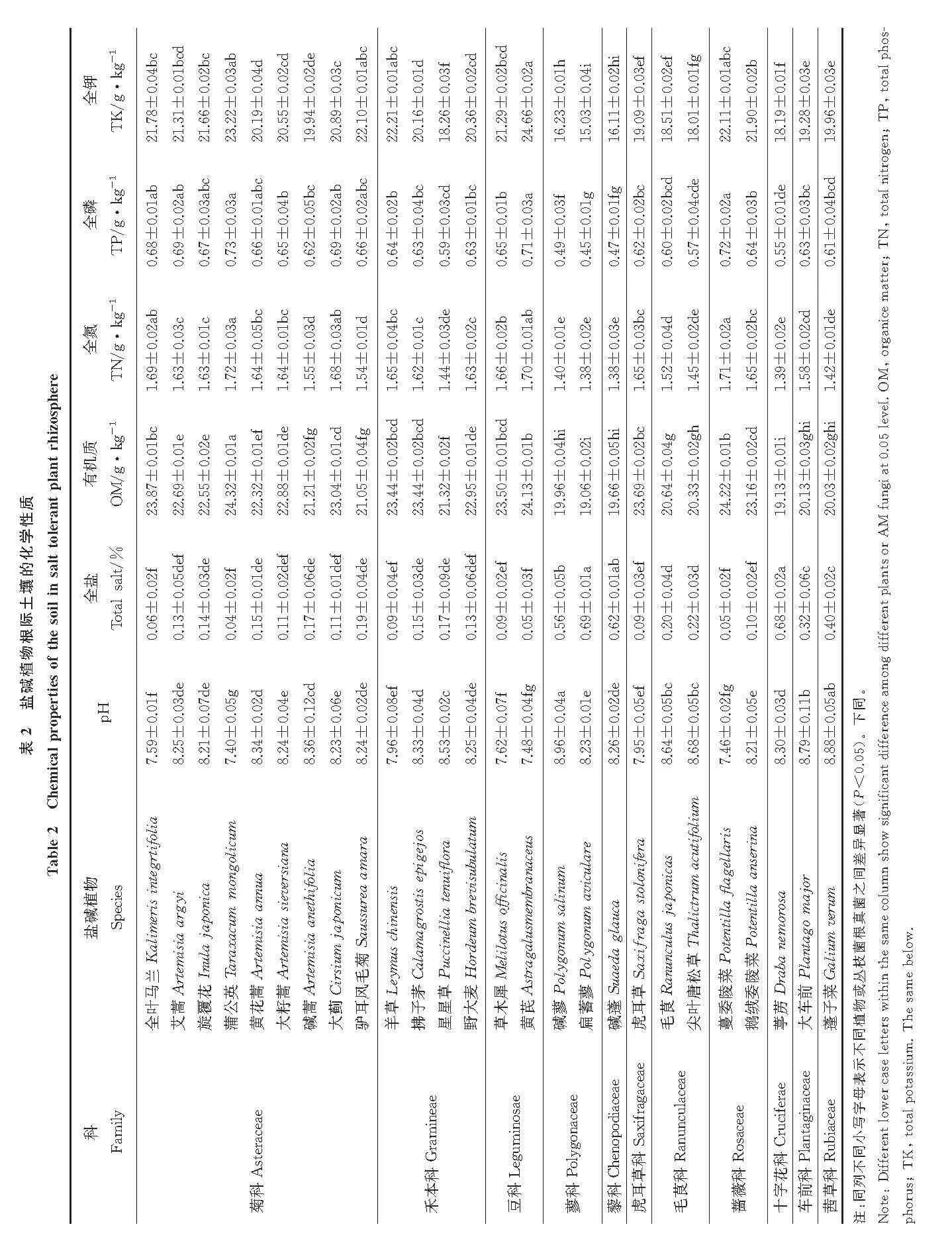
土壤理化性质参照《土壤农化分析》[11]和中国科学院南京土壤研究所的方法测定[12]。土壤pH 值采用玻璃电极法,用雷磁PHSJ-4A 测定;土壤全盐量采用浸出液电导率法,用雷磁DDSJ-308A 测定;土壤有机质采用重铬酸钾-浓硫酸法,用数显JXX1-HH-8 油浴锅测定;土壤全氮量采用半微量凯氏法,用ATN-1100 全自动凯氏定氮仪测定;土壤全磷量采用硫酸-高氯酸消煮法测定;土壤全钾量采用NaOH 熔融-火焰光度计法测定(上海,FP6450 型火焰光度计)。
1.4 AM 真菌的多样性测定
参考张美庆等[13]和秦燕燕等[14]的方法计算AM真菌孢子密度(Spore Density,SD)、物种丰度(Species Richness,SR)、分 离 频 度(Frequency,F)、相对多度(RelativeAbundance,RA)、Shannon-Wiener多样性指数(H)和重要值(Importance Value,IV)。

表1 松嫩盐碱草地26 种植物分布情况Table 1 The distribution situation of 26 plants in Songnen saline-alkaline grassland
SD=AM 真菌所有种的孢子数/土壤样本数;
SR=AM 真菌总种数目/土壤样本数;
F = AM 真菌某菌种出现的次数/总样本数×100%;
RA=AM 真菌某菌种的孢子数/AM 真菌总孢子数×100%。
利用Shannon-Wiener 多样性指数(H)来描述AM真菌的物种多样性,H=Piln Pi,利用重要值[IV=(F+RA)/2 ×100%]将AM 真菌优势度等级划分为优势属(种)、最常见属(种)、常见属(种)和稀有属(种)4 个等级,相应的IV 值分别为:IV >50%、30% <IV≤50%、10% <IV≤30%和IV≤10%。
1.5 数据分析
采用SPSS 17.0 对AM 真菌土壤理化性质和多样性指标进行单因素方差分析(Duncans)和多重比较(α=0.05),用平均值和标准误表示测定结果;分析土壤因子与AM 真菌之间的相关性;采用Microsoft Excel 对数据进行处理和绘图。
2 结果
2.1 土壤理化性质
松嫩盐碱草地26 种植物根际土壤的pH 值和全盐量相差不大(表2),pH 最高的植物为碱蓼,达到了8.96,最低的为蒲公英,仅为7.40;土壤全盐量平均为0.22%,这表明松嫩盐碱草地属于中盐度土质。各植物根际土壤的有机质、全氮、全磷、全钾含量差异明显,26 种植物根际土壤的有机质含量平均值为22.03 g·kg-1,全氮量平均值为1.57 g·kg-1,全磷量平均值为0.63 g·kg-1,全钾量平均值为22.12 g·kg-1。26 种植物中,土壤的有机质含量、全氮量、全磷量的最高值均出现在蒲公英根际,分别为24.32、1.72 和0.73 g·kg-1,黄芪根际土壤的全钾含量达所有植物中的最高值,为24.66 g·kg-1;扁蓄蓼的有机质含量、全氮量、全磷量和全钾量均达到所有植物中的最低值,分别为19.06、1.38、0.45 和15.03 g·kg-1。
2.2 AM 真菌种类及资源分布
从26 种植物根围共分离出AM 真菌5 属40 种(表3、图1),已鉴定38 种,尚有两个未确定种(sp1 和sp2)。其中球囊霉属(Glomus)21 种,占总种数的52.5%;无梗囊霉属(Acaulospora)12 种,占总种数的30%;盾巨孢囊霉属(Scutellospora)4 种,占总种数的10%;巨孢囊霉属(Gigaspora)两种,占总种数的5%;和平囊霉属(Pacispora)只有玻利维亚和平囊霉(Pacispora boliviana)1 种。根内球囊霉(Glomus intraradices)和地表球囊霉(G. versiforme)广泛分布在植物根围土壤中,二者是研究地区的常见菌种;极大巨孢囊霉(Gigaspora gigantea)、圆齿盾巨孢球囊霉(Scutellospora crenulata)和亮色盾巨孢球囊霉(S. fulgida)均只在一种植物根际土中出现,是该地区的稀有菌种。
2.3 AM 真菌的孢子密度、物种丰度和多样性指数
26 种植物根际土壤中均有不同程度的AM 真菌孢子存在(表4)。蒲公英的物种丰度和Shannon-Weiner 指数均达到所有植物中的最高值,分别为15.81 和1.43;蓼科植物AM 真菌资源分布少,Shannon-Weiner 指数低,其中扁蓄蓼孢子密度、物种丰度和Shannon-Weiner 指数均为所有植物中的最低值,仅有452.3 cfu·kg-1、9.92 和0.38。
2.4 AM 真菌分离频度、相对多度和重要值
试验结果表明,球囊霉属的分离频率、相对多度和重要值在26 种植物中较高,是优势属;无梗囊霉属为常见属,巨孢囊霉属的重要值仅有3.01%,是研究地的稀有属。根内球囊霉的分离频度、相对多度和重要值分别为80.77%、6.47%和43.62%,是该研究区的优势菌种,地表球囊霉仅次于根内球囊霉,是常见菌种,这表明二者对盐碱环境有较强的耐受力。极大巨孢囊霉、圆齿盾巨孢球囊霉和亮色盾巨孢球囊霉的重要值在26 种植物中均相对偏低,是研究区的稀有种(表5)。
2.5 土壤因子与AM 真菌的相关性
试验结果表明,土壤各因子与AM 真菌多样性密切相关(表6),土壤营养因子对AM 真菌的影响要强于盐度因子,其中pH 值与孢子密度和物种丰度之间表现出显著负相关(P <0.05),而全盐量与AM 真菌的孢子密度、物种丰度和多样性指数均没有显著相关性(P >0.05);土壤营养因子中,有机质和全氮量对AM 的影响最大,其中有机质和全氮量与孢子密度、物种丰度呈极显著正相关(P <0.01),与多样性指数呈显著正相关(P <0.05);全磷量与孢子密度、物种丰度和多样性指数均表现出显著相关(P <0.05),而全钾量与孢子密度、物种丰度和多样性指数之间没有显著相关性(P >0.05)。
3 讨论与结论
3.1 AM 真菌的多样性和分布
植物根围AM 真菌多样性指标反映了宿主植物与AM 真菌之间的亲和力,从采集的26 种常见盐碱植物根围中共分离鉴定出AM 真菌5 属38 种及两个未确定种,说明松嫩盐碱草地具有丰富的AM 真菌资源,这与张义飞[15]、周伟健[16]等对松嫩平原西部地区AM真菌资源分布的研究结果相似。26 种盐碱植物均能与AM 真菌形成良好的共生体系,但不同植物根际土壤的孢子种类、孢子密度、物种丰度、多样性指数、分离频度、相对多度和重要值均有明显差异,这说明AM 真菌与宿主植物形成共生体系时存在一定的偏好性,AM真菌与宿主植物间的亲和程度及二者的相互选择性在一定程度上决定了AM 真菌的生存和发育,这可能与不同植物根细胞对营养成分的种类和需求量不尽相同有关[17]。Tawaraya 等[18]发现,宿主植物种类和根系结构特征是AM 真菌共生与繁殖的重要因素,宿主植物不同,AM真菌种类、多样性也随之变化。
?

表3 26 种植物根际土壤AM 真菌资源及地区分布Table 3 The resources and distribution of AM fungi in rhizosphere of the twenty six plants

续表3
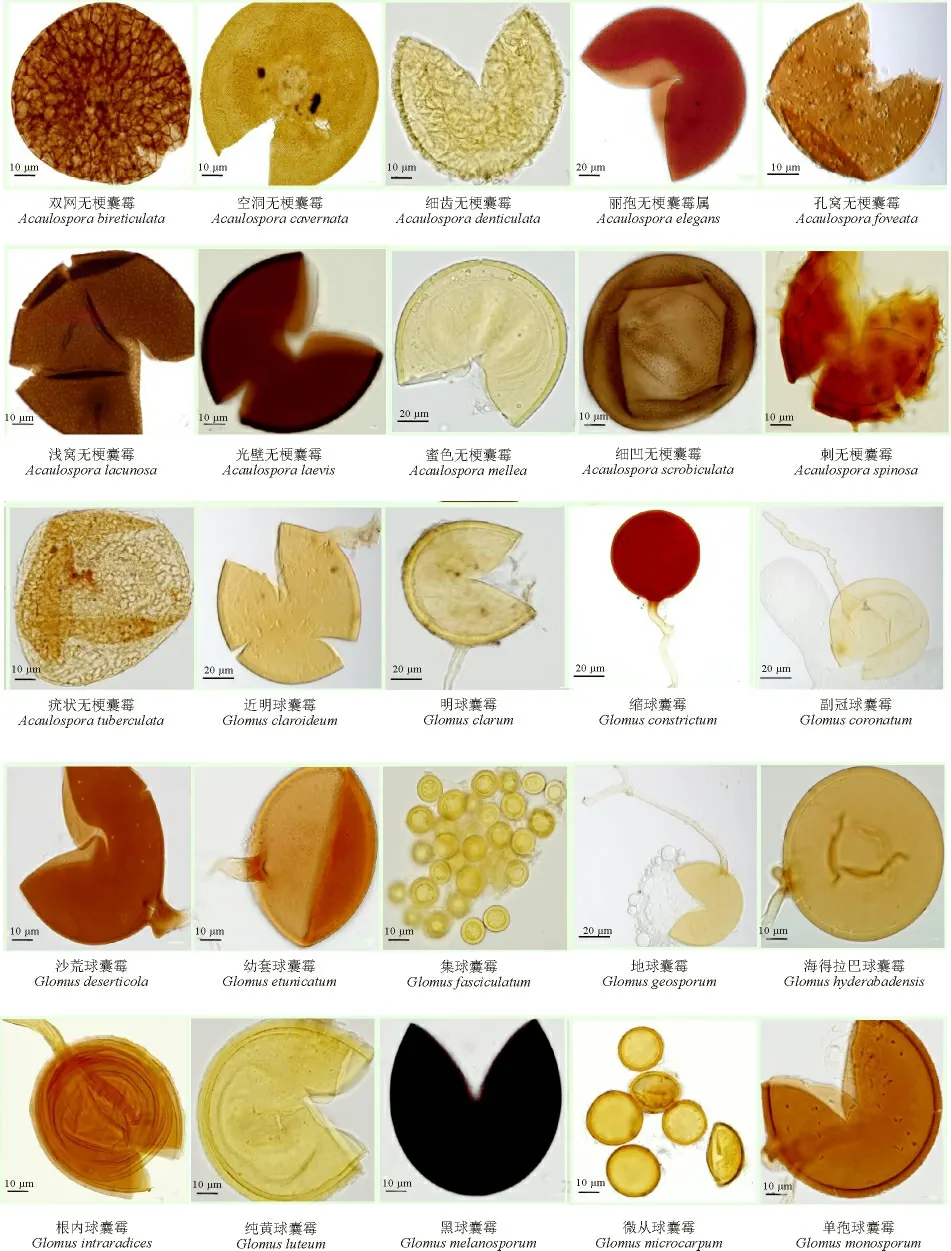
图1 本研究分离的5 属40 种AM 真菌Fig.1 40 AM fungi species of five genus isolated in this study
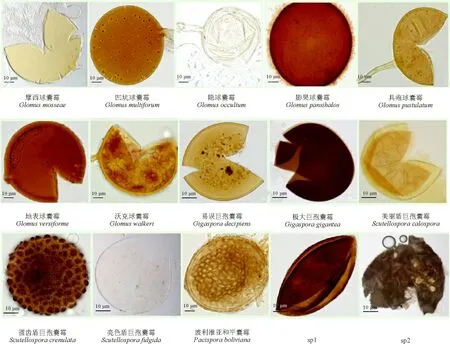
续图1 本研究分离的5 属40 种AM 真菌Continued Fig.1 40 AM fungi species of five genus isolated in this study
26 种盐碱植物根际土壤AM 真菌孢子的分离频度、相对多度和重要值总体上的趋势表现为球囊霉属>无梗囊霉属>盾巨孢囊霉属>巨孢囊霉属>和平囊霉属,其中球囊霉属和无梗囊霉属在不同植物中均占绝对优势,这个研究结果与球囊霉属和无梗囊霉属是广谱共生系统真菌的观点符合[19]。Joniper 和Abbott[20]发现,不同AM 真菌对盐胁迫的耐受力不同,盐碱对根内球囊霉和地表球囊霉的影响较小,但对美丽盾巨孢囊霉和光壁无梗囊霉孢子的萌发及芽管伸长有一定程度的抑制[20]。根内球囊霉和地表球囊霉是研究区共同的优势菌种,细凹无梗囊霉和集球囊霉在松嫩盐碱草地分布较多,是植物根际土中常见菌种,表明这几种AM 真菌对盐碱环境的适应力相对较强,同时与26 种植物均表现出较好的亲和力,可作为松嫩盐碱草地筛选优势菌种的依据。
3.2 土壤因子与AM 真菌多样性的关系
Christopher 等[21]研究证实,土壤因子通过作用于宿主植物从而间接影响AM 真菌的生长、繁殖及生态分布。从表6 可知,孢子密度和物种丰度与土壤有机质和全氮量均表现为极显著正相关(P <0.01),与全磷量表现为显著性正相关(P <0.05)。这是因为AM真菌能促进宿主植物对土壤中NH4+、铵态氮、硝态氮和氨基酸的吸收,通过菌丝网的进一步过滤,参与植物氮素代谢;菌根吸收NH4+后,释放H+,使土壤pH 值降低,从而刺激根系对土壤磷元素的吸收利用[22]。研究结果表明,有机质和全氮量表现出极显著正相关(P<0.01),有机质在一定范围内增进AM 真菌菌丝的形成和发育,但过高或过低均会抑制菌根的功能发挥[23],在对西藏高原针茅的研究中发现,土壤有机质含量在20.2 ~77.4 g·kg-1时,AM 真菌多样性表现为最优[24],故本研究认为全氮量也仅能在一定范围内促进AM 真菌的生长和发育,陈宁等[25]的研究证实了这一点,并提出当氮磷含量比为4∶ 2 时,最有利于AM真菌的生长,因为磷元素含量过高,反而会抑制菌丝的伸长和增粗,这可能是由于全磷量过高打乱了根系分泌物的平衡,使细胞膜透性受到破坏[26]。本研究结果显示(表6),土壤pH 值与孢子密度和物种丰度均呈现出显著的负相关,与多样性指数并无显著相关性(P>0.05),这表明土壤的盐度在一定程度上抑制了AM真菌的产孢能力,这与任爱天等[27]的研究结果一致。但Baker 等[28]在对惠农盐化灌淤土植物的研究中却得到了相互矛盾的结果,当灌溉水NaCl 的浓度达到0.3 mol·L-1时,几乎完全抑制了AM 真菌的生长繁殖 能 力, 而 雅 美 紫 菀(Aster tripolium)的根系却依然有菌根侵染,所以Johnson 等[29]报道,认为AM 真菌的多样性和土壤盐度关系不大,主要是受土壤养分因子的限制,这与本研究结果一致,相比于土壤盐度,土壤养分因子对AM 真菌多样性的影响相对更大。土壤全钾量与全盐量对AM 真菌的多样性均无显著相关关系,这可能是由于钾元素易与周围土壤形成难溶于水的矿物质(KAlSi3O8),致使植物无法吸收的缘故,Ouimet 和Camiré[30]的研究也证实了这一点。

表4 26 种植物AM 真菌孢子密度、种的丰度和Shannon-Winner 多样性指数Table 4 Spore density,species richness and Shannon-Winner diversity index of the AM fungi in 26 plant species
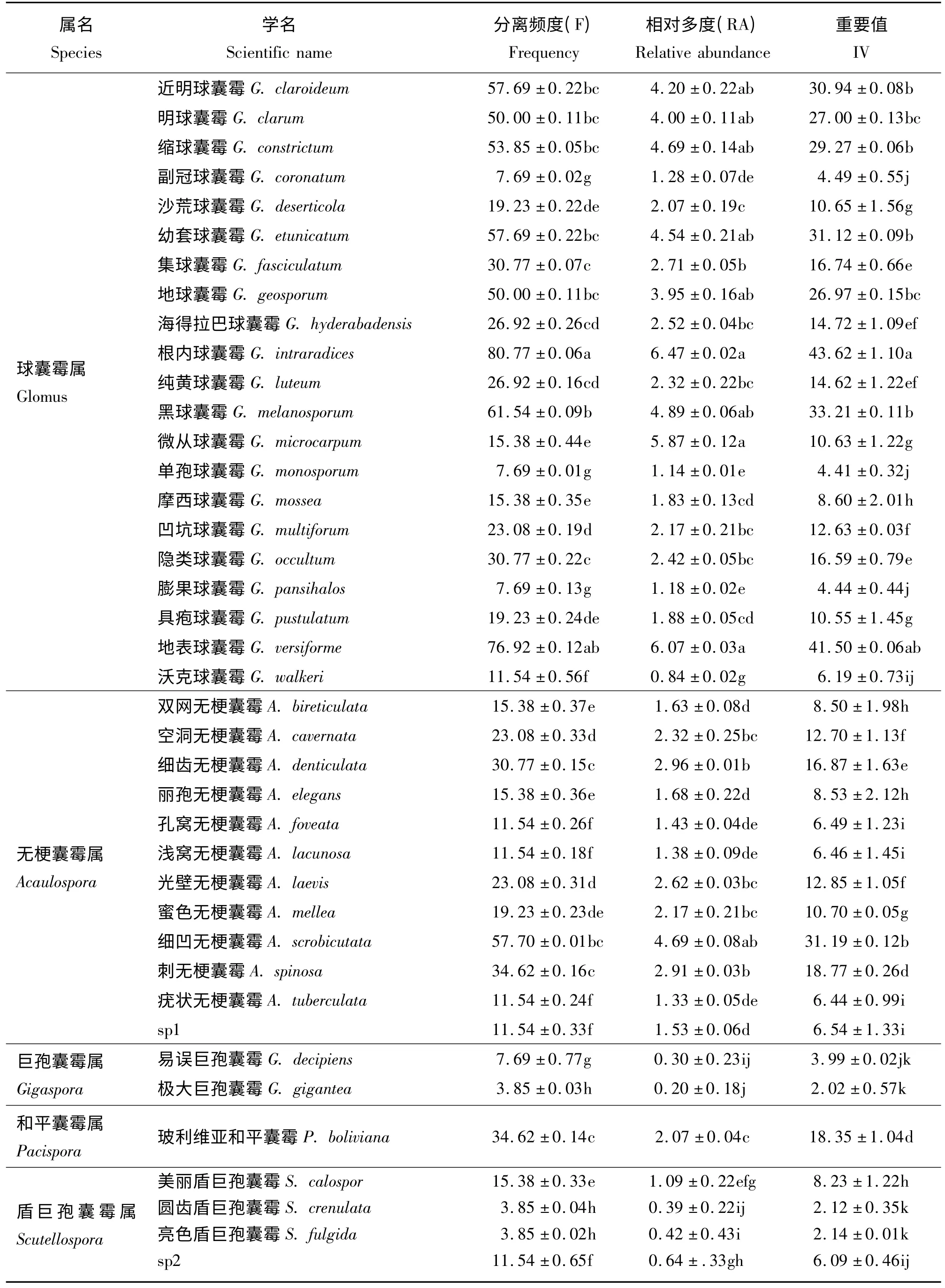
表5 26 种植物AM 真菌分离频度、相对多度、重要值Table 5 Frequency,relative abundance and importance value of the AM fungi in twenty six plants %
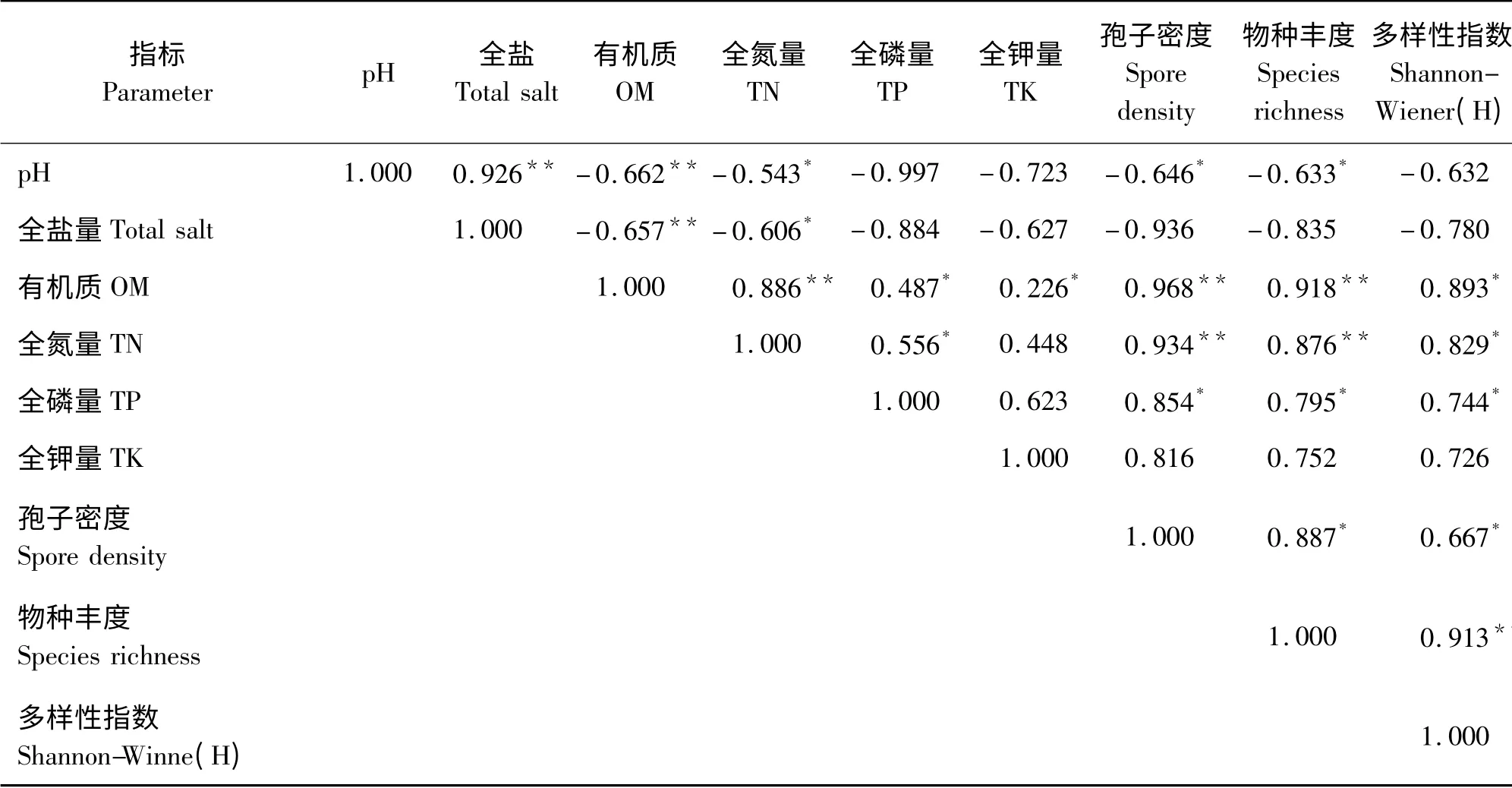
表6 土壤特性与AM 真菌多样性之间的相关性Table 6 The correlation between soil properties and AM fungi species diversity
本研究结果表明,在分离鉴定出的5 属38 种AM真菌中,球囊霉属是研究区的优势属,无梗囊霉属为常见属,由此可见,二者对盐碱环境有较强的耐受力;而巨孢囊霉属和盾巨孢囊霉属所占的比例较少,是研究区的稀有属,这与张美庆等[19]对我国北方农田土壤AM 真菌生态分布的研究结果相似。
从研究结果可知,肇东地区松嫩平原盐碱植物根际土中AM 真菌资源十分丰富。根内球囊霉和地表球囊霉是不同采样区域的共同优势菌种,而集球囊霉和缩球囊霉也高频率地出现在不同的植物根际,这说明同一样地同一宿主植物可以同时被多种AM 真菌侵染,不同样地同一菌种在同一宿主植物上出现的频度也可能存在差异[31],以上研究结果为进一步探究松嫩平原AM真菌的多样性提供了依据。
[1] 李取生,李晓军.松嫩平原苏打盐碱地治理与利用[J].资源科学,2003,25(1):15-20.
[2] 邱天.松嫩平原芦苇的生物学和生态学特征[J].草业科学,2014,31(2):300-305.
[3] 汤洁,李昭阳.松嫩平原西部草地的时空变化特征[J].资源科学,2006,28(1):63-69.
[4] 王斌,姚兆群,赵思峰.AM 真菌对盐胁迫下番茄幼苗生理特征及AVP1 表达的影响[J].西北植物学报,2013,33(10):2016-2023.
[5] Sheng M,Tang M,Zhang F F,Huang Y H.Effect of soil factors on arbuscular mycorrhizal fungi in saline alkaline soils of Gansu,Inner Mongolia and Ningxia[J].Biodiversity Science,2011,19(1):85-92.
[6] Prasad A,Kumar S.Heavy metals and arbuscular mycorrhizal (AM)fungi can alter the yield and chemical composition of volatile oil of sweet basil (Ocimum basilicum L.)[J].Biology and Fertility of Soils,2011,47:853-861.
[7] Isabel B,Mário C,Luís A.Managing arbuscular mycorrhizal fungi for bioprotection:Mn toxicity[J].Soil Biology & Biochemistry,2014,68:78-84.
[8] 孟繁荣.林木菌根学[M].哈尔滨:东北林业大学出版社,1996:137-150.
[9] Ianson D C,Allen M F.The effects of soil texture on extraction of vesicular arbuscular mycorrhizal spores from arid soils[J].Mycologia,1986,78:164-168.
[10] Schenck N C,Péréz Y.Manual for the Identification of VA Mycorrhizal Fungi[M].2nd edn.INVAM,USA,Gainesville:University of Florida,1989.
[11] 鲍士旦.土壤农化分析[M].北京:中国农业出版社,2000:22-42.
[12] 中国科学院南京土壤研究所土壤物理研究室.土壤物理性质测定法[M].北京:科学出版社,1978:11-86.
[13] 张美庆,王幼珊,邢礼军.我国东、南沿海地区AM 真菌群落生态分布研究[J].菌物系统,1998,17(3):274-277.
[14] 秦燕燕,蒋斌,曹秀文,冯宜明,李丹春,杨萌萌,向梅,陈蓉,李波,车宗全.间伐强度对白龙江林区云杉人工林下植物多样性及其更新的影响[J].草业科学,2014,31(4):599-606.
[15] 张义飞.松嫩平原盐碱化草地丛枝菌根真菌资源及其生态作用的研究[D].长春:东北师范大学博士学位论文,2011.
[16] 周伟健.丛枝菌根真菌与施肥对东北草甸草原的植物生产力和物种多样性的影响[D].长春:东北师范大学硕士学位论文,2013.
[17] Encarnación C L,Félix G O.Fisheries structural policy in the European Union:A critical analysis of a subsidised sector[J].Ocean and Coastal Management,2014,102:200-211.
[18] Tawaraya K,Saito M,Morioka M.Effect of phosphate application to arbuscular mycorrhizal onion on the devel opment and succinate dehydrogenase activity of internal hyphae[J].Soil Science and Plant Nutrition,1994,40:667-673.
[19] 张美庆,王幼珊,张弛,黄磊.我国北方VA 菌根真菌某些属和种的生态分布[J].真菌学报,1994,13(3):166-172.
[20] Juniper S,Abbott L K.Soil salinity delays germination and limits growth of hyphae from propagules of arbuscular mycorrhizal fungi[J].Mycorrhiza,2006,16,371-379.
[21] Christopher N,Elke G,Liliane R.Collembola grazing on arbuscular mycorrhiza fungi modulates nutrient allocation in plants[J].Pedobiologia International Journal of Soil Biology,2014,57(3):171-179.
[22] 李侠,张俊伶.丛枝菌根真菌对氮素的吸收作用和机制[J].山西大同大学学报(自然科学版),2008,24(6):75-78.
[23] 刘润进,陈应龙.菌根学[M].北京:科学出版社,2003:10-110.
[24] 彭岳林,杨敏娜,蔡晓布.西藏高原针茅草地土壤因子对丛枝菌根真菌物种多样性的影响[J].应用生态学报,2010,21(5):1258-1263.
[25] 陈宁,王幼珊,杨延杰.不同氮磷比例营养液对AM 真菌生长发育的影响[J].植物营养与肥料学报,2007,13(1):143-147.
[26] Mnthukumar T,Udaiyan K.Growth and yield of cowpea as influenced by changes in arbuscular mycorrhiza in response to organic manuring[J].Journal of Agronomy and Crop Science,2002,188(2):123-132.
[27] 任爱天,鲁为华,杨洁晶,马春晖.石河子绿洲区苜蓿地丛枝菌根真菌的多样性及与土壤因子的关系[J].草业科学,2014,31(9):1666-1672.
[28] Baker A,Sprent J I,Wilson J.Effects of sodium chloride and mycorrhizal infection on the growth and nitrogen fixation of Prosopis juliflora[J].Symbiosis,1995,19(1):39-51.
[29] Johnson Green P,Kenkel N C,Booth T.Soil salinity and arbuscular mycorrhizal colonization of Puccinellia nuttalliana[J].Mycological Research,2001,105(9):1094-1110.
[30] Ouimet R,Camiré C. Effect of soil K,Ca and Mg saturation and endomycorrhization on growth and nutrient uptake of sugar maple seedlings[J].Plant and Soil,1996,179:207-216.
[31] 贺学礼,李斌.VA 菌根真菌与植物相互选择性的研究[J].西北植物学报,1999,19(3):471-475.
