甘南高寒草甸主要毒杂草光谱特征分析
2015-04-08胡远宁孟宝平杨淑霞梁天刚
胡远宁,崔 霞,孟宝平,杨淑霞,梁天刚
(1.草地农业生态系统国家重点实验室 兰州大学草地农业科技学院,甘肃 兰州730020;2.兰州大学西部环境教育部重点实验室,甘肃 兰州730000)
20世纪80-90年代,高光谱遥感技术得到了充分发展,成为当前遥感领域的前沿技术。由于具有光谱分辨率较高(<10 n m)且光谱覆盖不间断等特点,高光谱遥感能很容易地探测到本来在宽波段遥感中不可能探测的物质[1],从而拓宽了人们解读地物的方式和方法,为各种地物特征的研究提供了丰富的研究方法。此外,高光谱以高分辨率、信息丰富等特性在植被特征参数计算方面优于常规遥感,近年来,利用高光谱遥感对地表植被生物化学参数的无损伤估测理论与技术得到了充分发展,基于高光谱的草地植被遥感为植被分类研究及应用提供了新的思路[2]。
草地是陆地生态系统的重要组成部分,近年来草地退化问题日益严重,加强草地资源动态监测是目前草地管理的热点。高光谱的发展为草地监测提供了新的手段,分析地物光谱反射特性不仅有助于区分草地类型,进行草地动态监测,而且还可为卫星数字资料进行计算机处理提供重要信息。许多学者对不同草地类型进行了光谱观测和分析,范燕敏等[3]测定了新疆部分天然草地类型及植物的反射光谱数据,结果表明,不同草地类型、不同植物光谱曲线特征差异明显,同时环境因子也对植物的反射光谱影响显著,高光谱遥感在草地植物生理生化研究及草地调查和监测中应用潜力巨大。盖颖颖等[4]利用草地植物花期独有的光谱特征,找出了不同种类花的光谱差异,为花期物种识别提供了参量化的依据;并在此基础上,尝试利用线性解混模型对混合光谱进行分解,从而估算出花覆盖度并加以验证。钱育蓉等[5]从光谱仪采集的野外高光谱谱线中,提取出典型荒漠植被的高光谱特征,为基于高光谱遥感的草地分类研究提供了科学依据;同时,为及时、准确地掌握草地资源的变化情况,指导人们正确利用草地资源、保护草地生态环境提供了理论依据。然而,目前草地高光谱的研究多限于草地退化光谱响应[6]、草地土壤营养元素测定[7]及草地优势种光谱特征分析等方面[8-9],针对高寒草甸毒杂草光谱特征的分析研究还不多见。
针对以上问题,本研究以甘南高寒草甸主要毒杂草为研究对象,利用地物光谱仪采集的野外高光谱数据,对甘南高寒草甸主要毒杂草进行基于光谱特征参量及光谱指数等的分析,提取出甘南高寒草甸主要毒杂草的光谱内在特征,以期为草地毒杂草的动态监测研究提供科学依据。
1 材料与方法
1.1 研究区概况
研 究 区(102°23′- 102°26′ E,35°05′-35°07′N)位于甘肃省甘南藏族自治州夏河县桑科乡(图1)。夏河县地处青藏高原东北边缘(102°30′-103°40′E,34°10′-35°40′ N),海拔2 800~4 600 m,全县土地总面积为6 273.88 k m2,其中可利用草地面积50.26万h m2,占全县土地总面积的80.1%[10]。气候属寒冷湿润类型,高原大陆性气候特点比较明显。年均温2.6 ℃,年降水量在450 mm 以上,年均无霜期56~109 d[11]。全县主要的草地类型为高寒草甸类和山地草甸类,草地植物多达72科628种,可食用牧草近600种[10-12]。

图1 研究区及地面实测点示意图Fig.1 Location of study area and investigated sites
1.2 数据获取
本研究选取了研究区中较为典型的5种常见毒杂草作为重点监测对象,分别为青藏狗娃花(Heteropappus bowerii)、箭叶橐吾(Ligularia sagitta)、披针叶黄华(Ther mopsis lanceolata)、乳白香青(Anaphalis lactea)和鹅绒委陵菜(Potentilla anserina)[13]。
1)青藏狗娃花,低矮,垫状,茎纤细,被白色密硬毛;叶两面密生白色长粗毛或上面近无毛,有缘毛。
2)箭叶橐吾,茎直立,叶片箭形、戟形或长圆状箭形,长2~20 c m。
3)披针叶黄华,茎直立,被棕色长伏毛。小叶倒披针形或长椭圆形,先端钝圆或急尖,背面被棕色长伏毛。
4)乳白香青,全株密被灰白色绒毛。根茎木质化,分枝;茎丛生,直立。
5)鹅绒委陵菜,多年生匍匐草本。羽状复叶,叶正面深绿,背后如羽毛,密生白细绵毛。
光谱测量时间为2013年9 月12-14 日,测量仪器为AvaField-3 地物光谱仪,其波段范围为300~2 500 n m,其中300~1 100 n m 的光谱分辨率为1.4 n m,光谱采样间隔为0.6 n m;1 100~2 500 n m 的光谱分辨率为15 n m,光谱采样间隔为6 n m。光谱测量要选择干燥、无风、晴朗无云或少云的天气进行,测量时间在12:00-14:00。纯毒杂草冠层光谱需要将多株毒杂草植株拢合成一束后测量,保证视场内只有毒杂草冠层,以避免土壤和其他植被类型的干扰[2,4-6]。光 谱 仪 视 场 角为25°,在距离纯毒杂草冠层顶部垂直高度约10 c m处用光谱仪探头垂直向下测量,光谱采集参数设置整合时间为100 ms,测量后及时进行标准白板校 正[2,5]。对每种毒杂草重复测量5次,剔除5次测量中的异常值后,对光谱数据进行平均及平滑处理,将得到的值作为处理后的5种毒杂草的光谱反射率值。
1.3 数据处理方法
本研究利用光谱特征参量法、光谱指数法和光谱重排法对光谱特征进行了提取[2,4],技术路线如图2。

图2 5种毒杂草光谱特征提取方法Fig.2 Methods to extract spectral characteristic of 5 poisonous weeds
1.3.1 光谱特征参量法 光谱特征参量法是将光谱上某个吸收特征或反射率峰特征参量化[2],本研究使用的光谱特征参量如表1所示。
本研究利用ENVI 4.8 软件光谱显示窗口中Plot_Function菜单下的1st Derivative模块求算原始光谱曲线的一阶微分曲线,以消除土壤背景影响及光谱数据之间的系统误差[2,4],突出植被的“绿峰”和“红边”特征。一阶微分的数学表达式如下[14]:

表1 光谱特征参量的定义及描述Table 1 Definition and description of the spectral feature parameters

其中,λi为i 波 段 波 长,ρ(λi)为i 波 段 反 射率,ρ′(λi)即为i波段一阶导数光谱值。
1.3.2 光谱指数法 本研究中使用的光谱指数如表2所示,并根据植被光谱的实际差异选择波段组合。其中R1和R2为波段的反射率值。

表2 本研究中使用的光谱指数及其表达式Table 2 Spectr al indices used in this study and their f or mulas
1.3.3 光谱重排法 有些情况下,不同植被的光谱在形状、反射率和变化趋势等方面差异不大,从原始光谱上很难将它们区分开来。因此,采用光谱重排的方法,打破原始光谱按照波长排列的次序,根据反射率大小对各个波段重新进行排列。经过光谱重排后,作为重排基谱的光谱曲线将呈现单调上升的趋势,而其他光谱曲线在按基谱的顺序重排后出现明显特征,且随基谱不同,相应特征出现的位置也不同[16]。
以两条光谱曲线r1=(r11,r12,…,r1L),r2=(r21,r22,…,r2L)为例来说明光谱重排的做法[16]。其中L为波段数。
1)以r1为基谱,将r11,r12,…,r1L的值按照从小到大的顺序重新调整,得到重排光谱为r*1=(r1k1,r1k2,…,r1kL),满足:当i<j 时,必有r1ki≤r1kj。
2)将r2也按照相同的顺序重新排序得到r*2=(r2k1,r2k2,…,r2kL)。
对重排后的光谱曲线r*1和r*2进行制图,比较二者间的差异。
2 结果与分析
2.1 5种毒杂草光谱反射率特征
箭叶橐吾的反射率最高,乳白香青的反射率最低(图3)。5种毒杂草光谱曲线趋势基本一致,但在不同波谱范围仍表现出不同的光谱特征。
2.1.1 510-600 n m 波段的主要特征 该范围是叶绿素的绿色强反射峰区[2],5种毒杂草在该波段区域的反射率较350-510 n m 均出现不同程度的上升(图3)。乳白香青和青藏狗娃花的“绿峰”幅值比其他3种草低很多(图4a,表3),这主要是因为乳白香青植株被灰白色绒毛覆盖,青藏狗娃花茎叶也生有白毛,其色素对可见光的反射大体相等。箭叶橐吾、鹅绒委陵菜和披针叶黄华的“绿峰”位置均介于521-525 n m,其中箭叶橐吾最大,鹅绒委陵菜和披针叶黄华次之,而其他两种草则无明显的该特征。
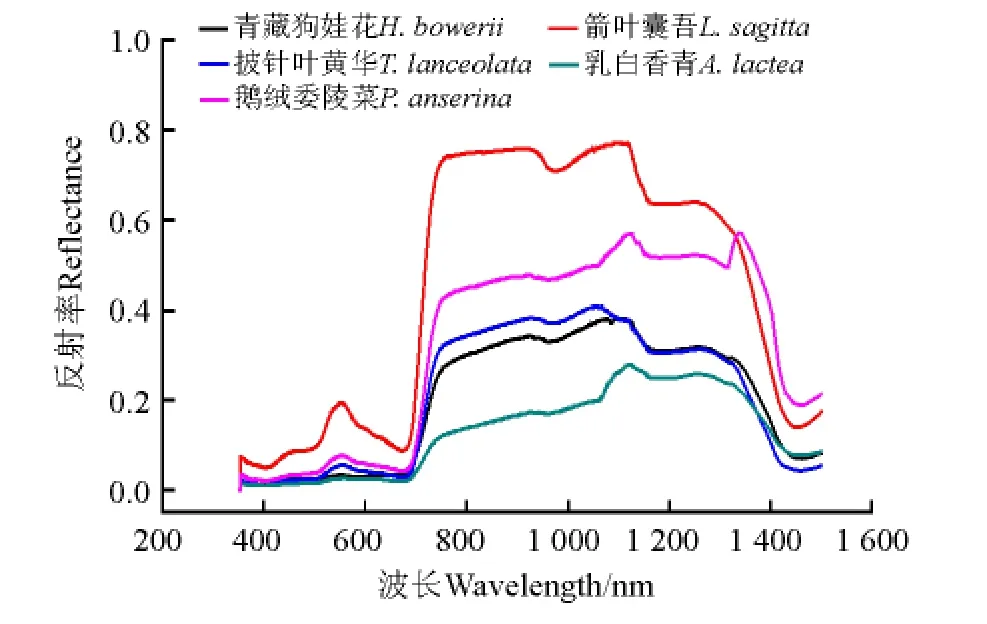
图3 平滑后的5种毒杂草光谱曲线Fig.3 Spectral curves of 5 poisonous weeds after filtering
2.1.2 600-680 n m 波段的主要特征 表征叶绿素等色素吸收的“红谷”[6],位于600-680 n m 的红橙光部分。在该波段范围内,植被的光合作用、生物量等形成过程、开花过程和光周期过程都以最大的速度来完成[2]。箭叶橐吾、鹅绒委陵菜和披针叶黄华有明显的“红谷”特征,而乳白香青和青藏狗娃花的“红谷”则不明显;且在550-670 n m,5种草的反射率一阶导数均为负数,表明这5种草在该范围吸收较强(图3,图4a,表3)。
2.1.3 680-800 n m 波段的主要特征 在680-800 n m 植被的光谱曲线出现反射率随着波长的增加而急剧升高的现象[2],称为“红边”参数,是植物所独有的光谱特征,用于表征植被营养、长势、水分、叶面积等特征[17-19]。“红边”位置会根据植被健康状况、成熟状态等发生偏移。在715-722 n m 的红光波段出现第2个峰值,且明显高于“绿峰”的峰值,表明反射率的变化率明显升高(图4a)。在680-800 n m,箭叶橐吾、鹅绒委陵菜和披针叶黄华的一阶微分曲线呈现明显单峰特征,而乳白香青和青藏狗娃花则没有明显该特征(图4b)。这5种草的“红边”位置分别为715、721、720、719和722 n m,对应的幅值分 别 为0.015、0.009、0.007、0.002 和0.005 n m-1(表3)。此外,在760 n m 附近,除乳白香青外,其他4种草均出现低谷特征。
2.1.4 800-1 400 n m 近红外波段的主要特征 在800-1 400 n m 波段范围内5种毒杂草的光谱反射率可分为两组:箭叶橐吾、鹅绒委陵菜的反射率较高,披针叶黄华、青藏狗娃花和乳白香青的反射率则相对较低(图3)。造成两组植物光谱差别的原因可能是不同植物的生长形态存在差异,前一组毒杂草叶面积指数大且生长密丛,多层叶子之间多次散射较强,因而具有较高的反射率;而后一组则由于茎秆直立细瘦或生长稀疏的形态特点,叶面积指数较小,自然生长状态下有效反射面积也较小。此外,在表征水分吸收的960和1 100 n m 处,乳白香青的水分吸收不明显,这可能是因为乳白香青茎部木质化,且光谱信号里混合了相当比重的枝条信息。
综上所述,箭叶橐吾反射率最高,而乳白香青反射率最低且无明显“绿峰”和水分吸收谷、其“红谷”几乎消失。因此,可依据这些光谱特征将箭叶橐吾和乳白香青从5种毒杂草中区分出来。

表3 5种毒杂草的光谱特征参量Table 3 Spectral characteristic parameters of 5 poisonous weeds

图4 5种毒草光谱曲线一阶微分图Fig.4 First-or der differential spectral curves of 5 poisonous weeds
2.2 5种毒杂草的光谱指数及其特征
披针叶黄华在680 n m 处的反射率低于青藏狗娃花,但在800 n m 处其反射率高于青藏狗娃花,且二者间的差异较680 n m 处大(图5a)。可以尝试通过归一化光谱指数建立识别标志,数学表达式如公式(2):

其中,R800和R680分别表示植被在800 和680 n m的光谱反射率。
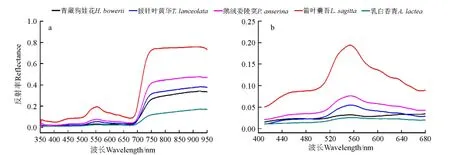
图5 5种毒杂草在350-950(a)和410-680 n m(b)间平滑后的光谱曲线Fig.5 Spectral curves of 5 poisonous weeds bet ween 350-950(a)and 410-680 nm(b)after filtering
青藏狗娃花、披针叶黄华及鹅绒委陵菜的光谱在740 n m 处反射率的差异比680 n m 处大。可以利用740和680 n m 两处光谱反射率的比值对这3种草进行区分,表达式如公式(3):

其中,R740和R680分别表示植被在740 和680 n m的光谱反射率。
披针叶黄华两种光谱指数的平均值均大于其他4种草的(表4)。因此,此标准可以把披针叶黄华从上述5种植被类型中区分出来。若基于前面的分组识别,该标准也可将披针叶黄华与青藏狗娃花、鹅绒委陵菜区分开来。

表4 5种毒杂草ND及SR 的平均值Table 4 The mean of ND and SR of 5 poisonous weeds
2.3 青藏狗娃花和鹅绒委陵菜的光谱重排
受光照和时相等外界条件的影响,青藏狗娃花和鹅绒委陵菜的反射率差异不足以表示二者之间的内在差别(图5b)。基于以上分析未能将二者很好地区分开来,因此,尝试利用光谱重排的方法进行更深入的分析,从而找出青藏狗娃花和鹅绒委陵菜的光谱差异。图6为对平滑后的青藏狗娃花和鹅绒委陵菜410-680 n m 波段范围内的462个波段光谱重排的结果。
以青藏狗娃花为基谱重排后,鹅绒委陵菜的光谱曲线在53波段之后表现出波动,并从203波段开始出现较大波动,与青藏狗娃花有明显不同(图6a)。以鹅绒委陵菜为基谱重排后,青藏狗娃花的光谱曲线在174波段后出现波动(图6b),波动的规律和尺度明显不同于以青藏狗娃花为基谱重排后鹅绒委陵菜光谱曲线的波动。因此,通过光谱重排的方法,可以有效地将青藏狗娃花与鹅绒委陵菜区分开来。
3 讨论与结论

图6 基于青藏狗娃花和鹅绒委陵菜的光谱重排Fig.6 Reordering results respectively based on Heteropappus bowerii and Potentilla anserina
草地植被的光谱受其色素、长势、叶面积及生长形态的影响很大,特别是可见光波段。采用光谱特征参量法、光谱指数法和光谱重排法,综合分析甘南高寒草甸5种主要毒杂草的光谱特征,可对其进行识别和区分。此外,本研究还存在以下有待进一步深入研究和解决的问题:1)滤波方法及参数的选取[2,4-5]。本研究采用地物光谱仪自带的Viewer软件对光谱曲线进行了简单的移动平均平滑处理,平滑窗口宽度作为重要参数之一,会对光谱曲线的某些特征产生影响:选用的平滑窗口的宽度如果太小,去噪、平滑的效果不佳;选用的窗口宽度若太大,一些有用的信息也会被平滑掉,从而造成光谱信号的失真[20]。在今后的工作中将采用Savitzky-Go1ay卷 积平滑滤 波 法[2,20]及 小 波 滤 波 方 法[2,4]对 原 始 光谱和导数光谱进行滤波处理。2)光谱指数的选取和构建。光谱指数是根据植被在不同反射波段的特性,通过对不同波段进行线性或非线性的组合计算出来的,可反映地表植被生长状况的数值。选择光谱指数时,可以根据植被光谱的实际差异进行构造,也可采用常见的植被指数。光谱指数可以扩大不同植被类型间的光谱差异,为植被类型的识别提供依据[2,4]。但研究对象的不同及人为因素的扰动对光谱指数的选取和构建存在较大影响,本研究只使用了ND 和SR 两种光谱指数,并根据植被光谱曲线的实际差异确定了所使用的波段组合,这些在一定程度上影响了区分效果。在今后的工作中,将使用多种光谱指数对研究对象进行区分,比较不同光谱指数的区分效果,并选择已经过验证或效果较优的波段组合。3)识别精度的验证。由于多种毒杂草混生的样方较为罕见,因此,本研究未设置毒杂草与毒杂草的混生样方。此外,目前常见的基于光谱技术的毒杂草识别研究多使用成像高光谱数据开展农田杂草研究[21-22],农田作物种类较少,混合样方情况较为简单,因而,对于农田成像光谱数据,现在已有成熟的像元解混算法便于对精度进行验证;而本研究的研究对象为甘南高寒草甸的主要毒杂草,所利用的光谱数据为单一的毒草或杂草的光谱曲线,主要开展了5种毒杂草在不同光谱波段范围内的特征差异分析,鉴于草地样方内的植物种类远比农田作物样方复杂,精度验证尚需开展相关研究。4)采集时间对植被光谱的影响。草地植被的内部结构和外部形态会随着生长发育阶段的不同而发生变化,相应地,其物理光学特征也将有所不同[14,23]。不同植被具有不同生长周期,不同季相植被的生长形态及内部的生物化学指标决定了其光谱形态特征的差异。此外,随季相变化,草地植被覆盖度及土壤裸露状态也发生变化,土壤背景的变化一定程度上会对草地植被的光谱响应值产生影响[2]。5)毒杂草与优势种的区分。由于9月份甘南高寒草甸优势种垂穗披碱草已经开始枯黄,光谱曲线受到较大影响,因此本研究未将其与毒杂草的光谱特征进行比较。6)在地物光谱仪测得的光谱曲线与遥感影像(如T M、HJCCD 数据)波段的比较,以及遥感影像反映的地物光谱反射率的精度评价方面,仍然需要进行深入研究。7)甘南高寒草甸光谱库的建立。由于测量种类较少、数据量较少,本研究未能建立甘南高寒草甸光谱库。在今后的工作中,将基于以上7点,加深对滤波及解混算法的研究,加入对优势种及不同季相植被的光谱特征研究,以期建立完整准确的甘南高寒草甸光谱库,为基于高光谱的甘南高寒草甸植被分类与毒杂草的定量研究奠定基础。
[1] 陈文霞,陈安升,蔡之华.基于高光谱吸收特征参数的分类研究[J].计算机工程与应用,2009,44(28):230-232.
[2] 李建龙.草地退化遥感监测[M].北京:科学出版社,2012:102-154.
[3] 范燕敏,武红旗,靳瑰丽.新疆草地类型高光谱特征分析[J].草业科学,2006,23(6):15-18.
[4] 盖颖颖,范闻捷,徐希孺,闫彬彦,王焕炯,刘媛.基于高光谱数据的呼伦贝尔草原花期物种识别和覆盖度估算[J].光谱学与光谱分析,2011,31(10):2778-2783.
[5] 钱育蓉,于炯,贾振红,杨峰,帕力旦·吐尔逊.新疆典型荒漠草地的高光谱特征提取和分析研究[J].草业学报,2013,22(1):157-166.
[6] 刘波,沈渭寿,李儒,杨兆平,林乃峰.雅鲁藏布江源区高寒草地退化光谱响应变化研究[J].光谱学与光谱分析,2013,33(6):1598-1602.
[7] 徐丽华,谢德体,魏朝富,李兵.紫色土土壤全氮和全磷含量的高光谱遥感预测[J].光谱学与光谱分析,2013,33(3):723-727.
[8] 张凯,郭铌,王润元,司建华,王小平.西北荒漠草甸植被光谱反射特征研究[J].地球科学进展,2006,21(10):1063-1069.
[9] 张凯,郭铌,王润元,司建华.甘肃省两种主要草地类型的光谱反射特征比较[J].农业工程学报,2009,25(2):142-148.
[10] 郭宏伟.夏河县畜牧业发展面临的问题及对策[J].草业与畜牧,2009(11):60-62.
[11] 杨延彪,刘占龙.关于夏河县草地畜牧业可持续发展的思考[J].家畜生态,2001,2(1):23-25.
[12] 杨延彪,张瑛.夏河县草地退化及综合治理措施[J].草业科学,1999,16(6):50-56.
[13] 张炳炎,郭致杰,郑果.中国西部农田杂草与综合防除原色图谱[M].兰州:甘肃文化出版社,2010:79-217.
[14] 张风丽,尹球,匡定波,李凤霞,周秉荣.环青海湖地区天然草地时序光谱特征参量分析[J].生态学报,2005,25(12):3154-3160.
[15] 宫兆宁,赵雅莉,赵文吉,林川,崔天翔.基于光谱指数的植物叶片叶绿素含量的估算模型[J].生态学报,2014,34(20):5736-5745.
[16] 耿修瑞.高光谱遥感图像目标探测与分类技术研究[D].北京:中国科学院遥感应用研究所,2005.
[17] 张敏.苹果褐斑病的高光谱遥感监测估算分析研究[D].西安:西安科技大学,2011.
[18] La mb D W,Steyn-Ross M,Schaare P,Hanna M M,Silvester W,Steyn-Ross A.Esti mating leaf nitrogen concentration in ryegrass(Loliu m spp.)past ure using the chlorophyll red-edge:Theoretical modelling and experi mental observations[J].Inter national Jour nal of Re mote Sensing,2002,23(18):3619-3648.
[19] Cho M A,Skid more A K.A new technique for extracting t he red edge position fro m hyperspectral data:The linear extrapolation method[J].Remote sensing of environ ment,2006,101(2):181-193.
[20] 褚小立,袁洪福,陆婉珍.近红外分析中光谱预处理及波长选择方法进展与应用[J].化学进展,2004,16(4):528-542.
[21] 刘波,方俊永,刘学,张立福,张兵,童庆禧.基于成像光谱技术的作物杂草识别研究[J].光谱学与光谱分析,2010,30(7):1830-1833.
[22] 李颖,张立福,严薇,黄长平,童庆禧.地面成像光谱数据的田间杂草识别[J].遥感学报,2013,17(4):855-871.
[23] 赵英时.植物遥感[A].黄荣辉.遥感应用分析原理与方法[M].北京:中国科学技术出版社,2003:366-412.
