红麻雄性不育研究进展及展望
2015-03-24李德芳陈安国唐慧娟李建军黄思齐
李 辉,李德芳,陈安国,唐慧娟,李建军,黄思齐
(中国农业科学院麻类研究所,湖南长沙 410205)
红麻雄性不育研究进展及展望
李 辉,李德芳*,陈安国,唐慧娟,李建军,黄思齐
(中国农业科学院麻类研究所,湖南长沙 410205)
红麻是一年生多用途天然纤维作物。从红麻不育系的类型、不育系的选育和不育的机理研究等方面进行了综述,并讨论了红麻不育系的利用、不育机理的研究方向和思路。
红麻;雄性不育;不育机理;杂种优势
红麻(HibiscuscannabinusL.)是锦葵科木槿属一年速生纤维作物,抗旱、耐盐碱,适应性广,生物产量高,主要用作纺织和包装的原材料。近年来,随着红麻用途的不断开发,红麻还用于纸浆、饲料、生物质能源、复合材料等方面。因此红麻被认为是21世纪最具发展潜力的多用途作物[1]。
杂种优势的利用可以显著提高红麻的生物产量。雄性不育(cytoplasmic male sterility,CMS)是利用红麻杂种优势的一条有效途径。利用CMS进行杂交制种可以提高杂交种子的纯度,免去人工去雄,节省大量的人力、物力,增加农作物的产量。雄性不育机理的研究一直是国内外研究的重点。本文主要针对红麻雄性不育的类型、杂种优势的利用、不育机理的研究等方面进行概述,以期为红麻雄性不育系的选育和杂种优势利用提供一定的理论依据。
1 红麻雄性不育系的类型
植物雄性不育的类型有3种:(1)细胞质雄性不育,是由细胞质因子控制的雄性不育类型,不育性容易保持,但是育性不易恢复。(2)细胞核雄性不育,是由细胞核基因控制的雄性不育类型,育性容易恢复,但是不育性不易保持。(3)质核互作型雄性不育,是由细胞核和细胞质共同控制的雄性不育类型,不育性既能保持同时也能够恢复。已经发现的红麻雄性不育材料有3种类型:细胞质雄性不育、细胞核雄性不育、质核互作型雄性不育。1958年,Pate JB,Joyner JF最早报道了红麻雄性不育现象[2]。1976年,Ugale发现了红麻细胞质雄性不育株,但未见其进一步的利用报道[3]。2001年,周瑞阳在海南南繁材料中首次发现了一株红麻雄性不育突变体,为“营养亏缺型雄性不育”[4]。2003年中国农业科学院麻类研究所红麻育种课题组在海南三亚育种材料中发现了细胞核雄性不育株,随后的2004年又发现了质核互作型雄性不育株。这些不育材料的发现为红麻杂种优势的利用奠定了坚实的基础[5,6]。
2 红麻雄性不育系的选育
2001年周瑞阳在海南冬繁材料“UG93”后代中发现了一株雄性不育突变体,其花药瘦瘪,不能开裂产生有效花粉。随后以该突变株为母本,通过饱和回交的方法于2003年选育出红麻细胞质雄性不育系“K03A”。2004年周瑞阳以“B4”、“L23A”为母本,分别与栽培品种“福红3号”、“泰红763”、“L23”、“PA258”、“粤丰1号”、“B19”、“917”和“722”进行杂交,并与父本进行饱和回交,最后成功转育成“福红3A”、“P3A”、“763A”、“917A”、“722A”、“L23A”等6个野败型细胞质不育系,其不育度达到99.9%,各不育系不育性稳定,并且育成了相应的保持系,配制出强优势的杂交组合。2003~2004年中国农业科学院麻类研究所红麻育种组在海南三亚南繁基地的育种材料中发现了新类型的红麻雄性不育株,并选配出了5个性状优异的组合:KCNms-1、KCNms-2、KCNms-3、KCNms-4、KCNms-5[1]。同时该课题组利用优良的种质对发现的不育系材料进行遗传改良,选育出红茎晚熟型、育性稳定、配合力高的红麻不育系LC0301A和相应的保持系LC0301B[6]。祁建民等利用细胞质雄性不育系L23A,经过多代回交转育,育成了光钝感红麻雄性不育系福红航1A,同时选育出了超高产杂交红麻新品种福航优1号、福航优2号,并通过了福建、安徽等省的鉴定[7]。
3 红麻雄性不育机理的研究
3.1 红麻雄性不育的细胞生物学
红麻雄性不育系的成功选育为红麻雄性不育机理的研究提供了可靠的物质基础。近年来,中国农业科学院麻类研究所、福建农林大学、广西大学等单位对红麻雄性不育机理的基础研究做了大量的工作。
朱丽梅等[8]对同质异核的红麻细胞质雄性不育系L23A和K03A小孢子的发生和败育过程进行了细胞学比较观察,结果发现:这两个不育系花粉的败育过程具有一定的相似性,败育都发生在四分体及单核小孢子时期;但是两者败育的具体过程又有一定差异。周琼等[9]以红麻细胞质雄性不育系L23A、保持系L23B和细胞核雄性不育系L23BS为材料,采用石蜡制片技术,利用光学显微镜对三者花药发育过程进行比较观察,结果表明:在L23A小孢子发育的不同阶段均出现败育现象,有的花粉母细胞在减数分裂以前就退化解体,形成空的花粉囊,这是最早阶段的败育表现;有的花粉母细胞不能形成正常的四分体小孢子;有的花粉母细胞能够形成四分体小孢子,但是不能正常释放从而造成败育。而L23BS小孢子败育的时期集中于四分体至小孢子期间,有些小孢子发育异常,有些小孢子不能从四分体里正常释放出来而影响其发育。2011年,李德芳[10]以质核互作型红麻雄性不育系LC0301A和相应的保持系LC0301B为材料对其花药、花粉粒进行电镜显微观察发现,LC0301A的败育发生在小孢子从四分体到单核小孢子时期,能够形成四分体,但是细胞壁不分化,细胞质明显减少,属于单核期败育型。不育系的花粉粒发育畸形,I2-KI不能染色,保持系花粉粒发育饱满,能够被I2-KI染色。推测不育系小孢子败育与中层、绒毡层提前衰退有关。
不同类型的红麻雄性不育系发生小孢子败育的时期主要集中在从四分体到单核小孢子时期,这一时期不育系的物质代谢和能量代谢不能正常进行,导致小孢子败育。
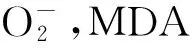
3.2 红麻雄性不育的分子生物学
在红麻雄性不育分子机理研究方面,我国科研工作者也做了大量的工作。白凤虎[12]通过构建红麻不育基因池和可育基因池的方法,利用RAPD引物进行筛选,最后获得了两条引物(S393、S1332)能够在不育基因池和可育基因池之间扩增出稳定的条带,通过亲本检测,发现这两引物所扩增的条带与不育基因是紧密连锁的。李辉等[13]利用BSA法构建近等基因池,用ISSR引物进行分子标记筛选,最后获得了一条能够在可育基因池和不育基因池之间产生稳定差异条带的ISSR引物U859,通过亲本鉴定这条引物所扩增的条带是与基因紧密连锁的。陈鹏等[14]以红麻不育系(P3A)和保持系(P3B)为材料,分别提取其线粒体DNA,用同源克隆的方法克隆了cox3基因,Southern印迹分析表明,cox3基因的组织形式在不育系和保持系中存在较大的差异,这种差异可能是由于线粒体基因组的重排导致的。赵杨[15]以红麻细胞质雄性不育系P3A及相应保持系P3B为实验材料,以红麻线粒体DNA为模板,通过简并引物克隆了可能与红麻雄性不育相关的基因nad1,nad2,nad3。唐向民等[16]以红麻细胞质雄性不育系L23A的花药为材料,采用3’RACE和RT-PCR技术克隆了一个与红麻雄性不育相关的基因Ms5。序列比对发现Ms5具有2种长度不一样转录本3’UTR,暗示了该基因的表达具有选择性转录终止的特征,不育性可能与该基因转录后水平的调控有关。陈艳翠等[17,18]以红麻质核互作型雄性不育保持系为材料,根据雄性不育相关基因ATPl的保守序列设计引物,通过RACE克隆的方法得到ATPl基因cDNA全长,测序后发现得到的基因序列在不育系和保持系间序列完全一致,这表明红麻质核互作雄性不育可能不是由ATPl基因的碱基差异造成的。
3.3 红麻雄性不育的蛋白质组学
李刚[19]以红麻细胞质雄性不育系(L23A)单核期、双核期的花药和保持系(L23B)单核期、双核期的花药为实验材料,分别提取相应的线粒体蛋白质,通过二维液相色谱结合串联质谱技术对其差异蛋白质组进行了分析、鉴定,结果表明,不育系花药线粒体中,参与能量代谢的蛋白质变化最大,其次是参与mtDNA转录、翻译、信号调节等的蛋白质。可见能量代谢的异常与细胞质雄性不育的关系最为密切。廖英明[20]以红麻细胞质雄性不育系(L23A)和保持系(L23B)为实验材料,分别选取幼蕾、大蕾、开花3个时期的功能叶进行差异蛋白组学分析,通过MALDI-TOF-MS/MS串联质谱鉴定得到了6个与红麻雄性不育相关的蛋白质点:核酮糖1,5-二磷酸羧化酶/加氧酶、糖体蛋白S3、苹果酸脱氢酶、二磷酸核酮糖羧化酶、转录因子LECl-A。这些差异蛋白的表达可能与红麻雄性不育密切相关。
红麻雄性不育的蛋白质组学研究表明,雄性不育的发生与能量代谢的紊乱,转录、翻译的异常,多个代谢途径的受阻等有关连,可见红麻雄性不育的发生是一个多因素参与的复杂的生物学过程。
4 讨论与展望
国内外科研单位对红麻不育从细胞生物学、分子生物学和蛋白质组学等方面进行了大量的研究并取得了一些成果,但是这些研究缺乏系统性、完整性。红麻为常异交作物,遗传基础复杂,遗传特点独特,雄性不育发生的机理复杂,所以红麻雄性不育机理至今还未揭开其神秘的面纱。为了更好地将红麻雄性不育系应用于红麻杂种优势的利用,进一步选育新的遗传基础广泛的红麻雄性不育系,加强红麻雄性不育机理的研究十分必要。
4.1 加强特殊配合力高的红麻雄性不育系的选育
红麻雄性不育系的特殊配合力是其杂种优势利用的关键。目前我国红麻质核互作雄性不育系和细胞质雄性不育系的遗传基础狭窄,不利于选配强优势杂交组合。为了更好地利用红麻杂种优势,可以尝试通过航天诱变、基因工程等技术获得具有不同遗传基础的红麻不育系源;还可以通过远缘杂交、植物细胞诱变等手段获得变异材料,从中选取新的不育材料,从而获得理想的红麻雄性不育材料。
4.2 加强红麻雄性不育机理的基础研究
红麻雄性不育的发生是一个复杂的生物学过程,涉及到能量代谢、信号的传导、基因的翻译、表达和修饰等过程,同时还可能有与环境互作的因素,这些都加大了不育机理研究的难度。红麻雄性不育机理的研究主要可以从三个方面进行:第一,从基因组的角度。克隆细胞质不育基因(线粒体、叶绿体)和细胞核不育基因,并对其进行序列测定,从基因的结构、功能方面来解释红麻雄性不育的机理。第二,从蛋白质组学的角度。蛋白质是生物功能的执行者,通过比较保持系和不育系的蛋白质组,对差异蛋白质进行筛选、鉴定并分析差异蛋白质的生物学功能,从而揭示红麻雄性不育的分子机制。第三,从植物细胞信号传导的角度。通过研究环境与基因表达互作的关系,来解释红麻雄性不育的机理。
[1] 李德芳,陈安国,唐慧娟.红麻质核互作型雄性不育系的发现及初步创制[J].中国麻业科学,2007,29(2):78.
[2] Pate JB,Joyner JF.The inheritance of a male sterility fact in kenaf(HibiscuscannabinusL.)[J].Agron J,1958,50:402-403.
[3] Ugale SP,Khuspe SS.Cytoplasmic genetic male sterility inHibiscuscannabinusL.[J].JMAU,1976,2(6):102-106.
[4] 周瑞阳.红麻雄性不育株的发现[J].中国农业科学,2002,35(2):212.
[5] 中国农科院麻类所红麻育种组.红麻雄性不育株的发现及其初步研究[J].中国麻业,2003,25(2):61.
[6] 李德芳,李建军,陈安国,等.质核互作型红麻雄性不育系LC0301A的选育[J].中国麻业科学,2013,35(4):172-174.
[7] 祁建民,林荔辉,方平平,等.红麻光钝感细胞质雄性不育系的选育与杂种优势利用[A].见:2012年中国作物学会学术年会论文摘要[C].福州:福建农林大学,2012.
[8] 朱丽梅,艾素云,周瑞阳.红麻细胞质雄性不育系小孢子败育的细胞学观察[J].作物学报,2007,33(6):999-1003.
[9] 周 琼,黎 桦,赵 嘉,等.红麻GMS与CMS小孢子败育过程的细胞学及组织化学比较[J].作物学报,2010,36(8):1414-1424.
[10] 李德芳.红麻优异雄性不育系的培育及其杂种优势利用的研究[D].北京:中国农业科学院博士学位论文,2011.
[11]周 琼,周瑶瑶,唐向民,等.红麻细胞质雄性不育系与保持系花药活性氧代谢差异比较[J].广西植物,2013,33(4):443-448.
[12]白凤虎.红麻杂种优势、配合力分析及雄性不育基因的RAPD标记[D].北京:中国农业科学院硕士学位论文,2007.
[13]李 辉,李德芳,陈安国,等.红麻雄性不育系的选育和不育基因的ISSR分子标记[J].中国农学通报,2008,24(8) 80-83.
[14]陈 鹏,周瑞阳.红麻线粒体中细胞质雄性不育候选基因cox3的克隆与分析[J].生物技术通讯,2011,22(1):37-40.
[15]赵 杨.红麻细胞质雄性不育相关基因nad的克隆与再生体系初探[D].南宁:广西大学硕士学位论文,2011.
[16]唐向民,金 刚,李 刚,等.红麻Ms5基因的克隆及多转录本分析[J].华中农业大学学报,2012,31(5):536-540.
[17]黄思齐,陈艳翠,李建军,等.红麻雄性不育相关基因atp1的表达初步分析[J].中国麻业科学,2013,35(5):221-225.
[18]陈艳翠,黄思齐,李建军,等.红麻质核互作型雄性不育相关基因atp1的克隆与分析[J].中国麻业科学,2013,35(2):63-68.
[19]李 刚.红麻细胞质雄性不育系线粒体蛋白质组差异分析与不育相关基因的发掘[D].南宁:广西大学硕士学位论文,2009.
[20]廖英明.红麻蛋白质组2-DE建立及不育系和保持系差异蛋白分析[D].福州:福建农林大学硕士学位论文,2011.
Progress and Prospect of Research on Kenaf Male Sterility
LI Hui,LI De-fang*,CHEN An-guo,TANG Hui-juan,LI Jian-jun,HUANG Si-qi
(Institute of Bast Fiber Crops,Chinese Academy of Agricultural Sciences,Changsha,Hunan 410205,China)
Kenaf is an annual multi-use natural fiber crop.In this paper,the research progress was summarized based on the type of kenaf male sterility line,the breeding of kenaf male sterility line and the mechanism of male sterility line,the using of kenaf male sterility and the research direction and ideas of male sterility mechanism were discussed.
kenaf(HibiscuscannabinusL.);male sterility; mechanism of sterility; heterosis
2014-10-16
李 辉(1980-),男,河北定州市人,博士后,主要从事红麻育种,Email:guangjunmuzi@126.com。
*通信作者:李德芳,男,研究员,主要从事红麻育种,Email:chinakenaf@126.com。
国家麻类产业技术体系红麻育种岗位资助(CARS-19-E07);亚州区域国际合作项目资助。
S563.503.2
A
1001-5280(2015)02-0206-04
10.3969/j.issn.1001-5280.2015.02.25
