不同调控措施下橡胶凋落叶分解速率研究
2015-03-23吴志祥管利民赖华英
金 龙 吴志祥 杨 川 管利民 赖华英
(1.中国热带农业科学院橡胶研究所,农业部儋州热带作物科学观测试验站,海南 儋州 571737;2.海南大学环境与植物保护学院,海南 海口 570228)
不同调控措施下橡胶凋落叶分解速率研究
金 龙1,2吴志祥1,2杨 川1管利民1赖华英1
(1.中国热带农业科学院橡胶研究所,农业部儋州热带作物科学观测试验站,海南 儋州 571737;2.海南大学环境与植物保护学院,海南 海口 570228)
通过原位网袋法,研究6、12、20 a和28 a 4组林龄橡胶凋落叶在不同调控措施下1 a内的分解过程。结果表明:橡胶凋落叶在0.07 mm和1.00 mm孔径网袋内分解趋势基本一致,表现为随时间变化分解呈现先快后慢2个阶段,经1 a分解后,0.07 mm和1.00 mm孔径网袋内残留率分别为36.84%和32.09%。橡胶凋落叶分解速率不仅受埋层位置影响,还与土壤肥力有关。经240 d分解后,地上组(0 cm)和在保留根系下非肥坑地下组(0~30 cm)、肥坑地下组(0~30 cm)残留率分别为32.09%、7.40%和34.58%。土壤肥力对橡胶凋落物分解的影响主要体现在分解中后期,至240 d时,4块林地肥坑和非肥坑平均残留率(保留根系组+去除根系组平均值)分别为35.62%和8.46%。6、12、20 a和28 a 林龄4组橡胶凋落叶原位埋置分解中,前期分解速率差异小,后期分解速率呈显著差异,至240 d时,残留率分别为2.94%、7.05%、14.62%和2.18%。橡胶根系前期促进凋落物分解,后期转为抑制,但橡胶根系没有显著性改变凋落物的分解速率。
橡胶林;调控措施;凋落叶;分解速率
凋落物分解是森林生态系统物质循环和能量流动的重要过程,其分解速度不仅决定了土壤养分的有效利用,同时也会影响陆地与大气间的碳分配[1-2]。凋落物分解过程主要受凋落物基质质量(内因)和气候环境因素(外因)的影响,包括凋落物的物理形态和化学组成、生物类群组成及数量、水热环境、土壤肥力和放置位置等[3-7]。另外,由于凋落物分解研究主要采用网袋法,因此,网袋孔径大小也会影响凋落物的分解速率[4、8]。在全球尺度上,温度和降水是主导凋落物分解在不同气候带间差异的决定性因素,而在同一气候区域内,凋落物分解速率则更多的受制于凋落物质量、立地条件和土壤理化性质的协同作用[7]。其中土壤环境是影响凋落物分解的一个重要环境因子。相同类型的凋落物在有机质背景低的土壤中分解速度则更快[9],但王进[10]研究发现,马尾松(Pinusmassoniana)、栓皮栎(Quercusvariabilis)及其混合凋落叶在不同土壤养分基质中的分解速率表现为高养分最快,中养分次之,低养分最慢。
虽然,在世界范围内开展森林凋落物的研究已有100多年的历史,但是,基于全球森林生态系统类型、分布区域及自然条件的复杂多样,如要探索更具普遍意义的分解规律及影响机制,仍需要针对特定类型森林生态系统凋落物开展基础性研究工作。橡胶林是我国热区一种最典型的人工经济林,其生态系统的健康发展不仅有利于改善当地生态环境,而且对提高胶农收益、促进经济发展具有直接的现实意义。截至2013年,我国植胶面积接近113.33万hm2,干胶产量达8.5×105t,分别排在世界第三位和第六位[11]。然而,目前我国对橡胶林生态系统凋落物的研究主要集中在凋落物年凋落量、凋落过程、凋落物组成、凋落物分解过程和速率以及凋落物分解过程中主要养分的释放上[12-14],而对不同环境下橡胶凋落物分解过程中的影响机制还尚未见到报道,鉴于此,本研究集中比较了橡胶凋落叶在不同环境下的分解状况,包括林地环境、土壤肥力、埋层位置、橡胶根系及网袋孔径对橡胶凋落叶分解速率的影响,为全面深入认识橡胶林凋落物的分解规律及影响机制提供理论依据,同时也可为橡胶林生态系统的经营管理提供参考。
1 试验地概况
橡胶凋落物分解试验地设在海南儋州中国热带农业科学院试验农场一队和三队内,位于北纬19°31′47″,东经109°29′30″,平均海拔144 m。属热带季风气候,年平均温度20.5~28.5 ℃,最冷月平均温度16.5~17.6 ℃,全年日平均气温≥10 ℃的积温为8 500~9 100 ℃;全年旱雨两季分明,5—10月为雨季,11月至翌年4月为旱季,年平均降雨量为1 607~2 000 mm,其中7、8、9月总降雨量占全年降雨量的70%以上;年平均相对湿度83%。试验区地形为缓坡丘陵(相对高度差<10 m),土壤为花岗岩母质化所形成的砖红壤,土层厚度约100 cm,土壤pH 4.52~5.86。
该试验区为第二代胶园,其中,地表放置试验样地林下隔行间作有用于压青肥的天堂鸟(Strelitziareginae),其他植被多为当年生草本到多年生小草本植物。
2 试验方法
2.1 凋落物收集
于2013年12月底至翌年1月初橡胶集中落叶期,在晴朗天气下,分别收集6、12、20 a 和28 a等4个林龄橡胶林新近凋落叶,于室内风干备用。同期在各样地现用施肥坑和对应的株行中间1.5 m处,分别隔行挖长×宽×高为120 cm×60 cm×40 cm的埋置坑,分别记为施肥坑和非施肥坑(CK),所有坑穴并排排列,共计4排,每排3个重复,共计96个穴位。为排除边际效应和其他人为干扰,所有坑穴距离道路最低在15 m以上。
2.2 凋落物分解试验
2.2.1 地表放置处理 凋落物分解采用目前通用的网袋法。按树龄划为4组,每组分别称取风干凋落叶(50.00±0.50)g,直接装入大小为45 mm×35 mm,孔径分别为1.00 mm×1.00 mm和0.07 mm × 0.07 mm的网袋中,每林龄大小网袋各15袋,共计120袋。于2014年1月10日分别将凋落物网袋随机放入12 a林龄段中,让其与土壤接触,并用枯落物将其覆盖。
2.2.2 地下埋置处理 地下埋置处理采用各林龄橡胶凋落叶原位埋置法。另称取相等质量的各林龄橡胶凋落叶装入相同规格的1.00 mm孔径网袋内,每林龄144袋,共计576袋。同时将事先挖好的非肥坑和肥坑均分为相等的两部分,其中一部分用塑料薄膜直立环壁包裹四周,空留出顶部和底部,阻断根系环壁进入网袋且防止雨季雨水截留,即为去根处理;另一组保持原样,根系可自由进入,即为保根处理。于2014年1月10日将4种林龄凋落物网袋分别原位埋入6、12、20 a和28 a林龄样地施肥坑和非施肥坑中的去根和保根处理的0~10、10~20、20~30 cm土层中,并用土将其覆盖填埋。
2.2.3 凋落物回收 网袋每2个月回收1次。凋落物回收选在天气晴朗之时,于2014年3月10日开始第1次取样,之后每隔60 d取样1次(根据天气情况前后推移2~3 d),每次每林龄每处理各取回3袋,去除袋内根系等杂物,待网袋及凋落叶附着泥土风干后,用毛刷将其仔细去除干净,于105 ℃烘箱中烘干至恒质量并计算残留率。
2.3 残留率计算
枯落物残留率:
Ri=(mi/m0)×100%
式中:Ri为第i月的残留率;mi为第i月所取样品的剩余质量;m0为落叶初始质量。
2.4 数据分析
用Excel 2007、Origin 9.0等软件对所有取样点数据进行整理、分析、计算、作图。
3 结果与分析
3.1 不同孔径网袋内凋落叶分解状况
用1.00 mm和0.07 mm 2种不同孔径网袋研究橡胶凋落叶的分解进程,残留率以4个林龄组凋落叶平均值计算,结果见图1。
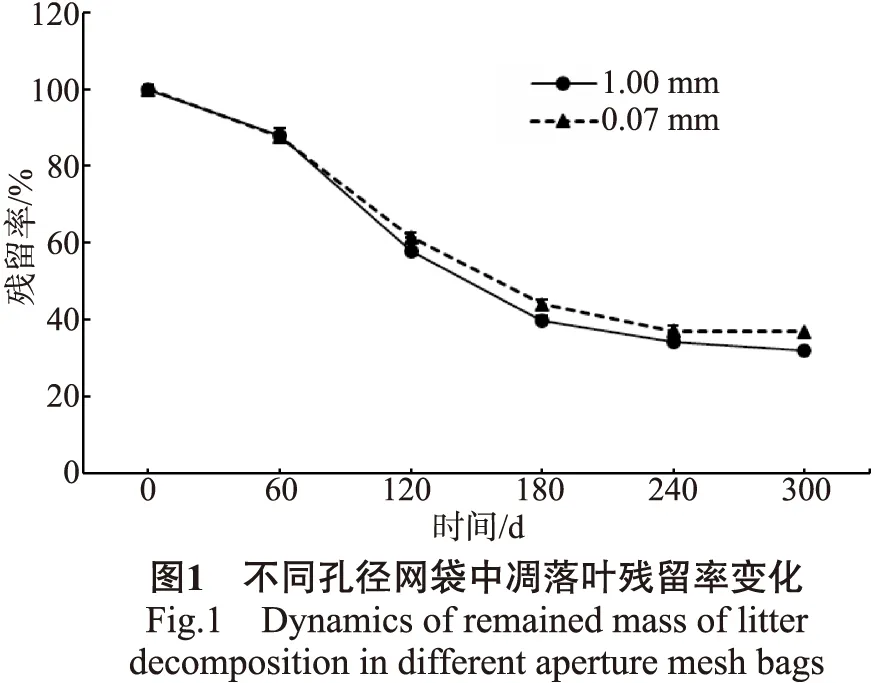
由图1可知,橡胶凋落叶在不同孔径网袋中的分解进程一致,即随时间推移残留率逐渐降低,残留率变化在180 d前呈快速下降趋势,180 d后开始趋缓。从分解速率看,大、小孔径网袋在分解前期(0~60 d)差异不显著,趋势线几近重合;60 d后,小孔径网袋分解速率显著低于大孔径网袋,分解至300 d时,大、小孔径网袋内残留率分别为32.09%和36.84%。整体看来,0.07 mm孔径网袋相比于1.00 mm孔径网袋减缓了凋落物的分解进程,这与张瑞清等的研究结果一致。
3.2 不同埋层中凋落叶分解状况
不同埋层凋落叶分解速率分别以4个林龄组1.00 mm孔径网袋内地表放置和0~10、10~20、20~30 cm保根处理组的平均残留率(合计为0~30 cm)表示,结果见图2。
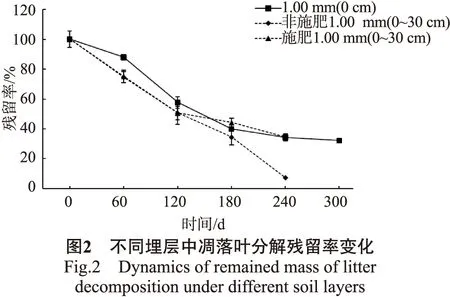
由图2可以看出,橡胶凋落叶在不同埋层中的分解过程有明显差异。其中,在非施肥坑0~30 cm土层中,残留率几乎呈直线下降趋势,至240 d时,残留率仅为7.40%;在施肥坑0~30 cm土层中,残留率在0~120 d变化较快,120 d后变化趋势开始放缓,至240 d时,残留率为34.58%;地表放置组中,凋落叶分解进程与施肥坑0~30 cm土层中基本相似,存在明显的快、慢2个不同分解阶段,即180 d前分解快,180 d后分解慢,至300 d时,残留率为32.09%。整个分解过程中,橡胶凋落叶分解速率表现为:地表放置处理<非施肥坑地下组;地上组分解速率只在前120 d显著高于施肥坑地下组,在后120 d,地上与地下两组分解速率相差不大。可见,不同肥力土壤埋置处理均可加速凋落物前期分解,而对凋落物分解后期的影响则与土壤肥力有关,即低肥力土壤能持续促进分解,而高肥力土壤的促进作用明显减弱,甚至转为抑制作用。
3.3 不同土壤养分下凋落叶分解状况
土壤肥力对橡胶凋落叶分解的影响分别以4个林地非施肥坑和施肥坑0~30 cm土层(保根+去根)凋落叶平均残留率来表示,结果见图3。
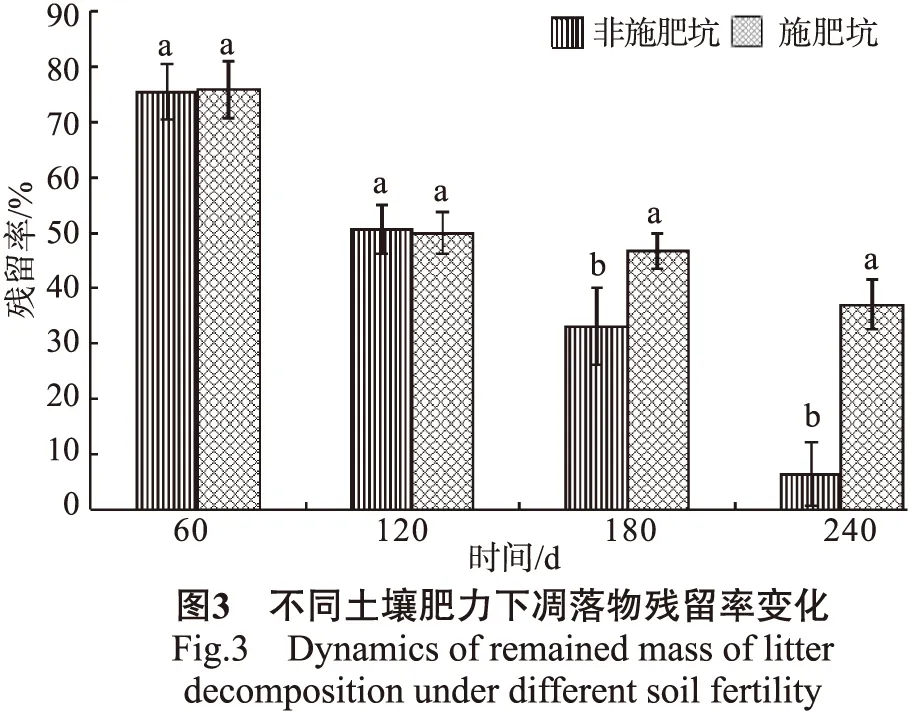
由图3可知,凋落叶在施肥坑与非施肥坑中的分解趋势表现出明显的差异,在240 d的研究期内,凋落叶残留率在非施肥坑中几乎是直线下降的过程;而在施肥坑中,除在前120 d有一个较大的下降过程外,之后表现为缓慢的降解过程。凋落叶在不同土壤肥力中质量损失差异,主要发生在分解的中后期(120 d后)。在分解的中前期(0~120 d),凋落叶在施肥坑和非施肥坑中的分解速率变化较为一致,残留率差异不显著;120 d后,凋落叶残留率在施肥坑与非施肥坑中表现为显著差异,且随分解进程的推移,差异也逐渐增大。至240 d时,非施肥坑和施肥坑橡胶凋落叶残留率分别为8.46%和35.62%,残留率相差值由13.26%(180 d)增加到27.16%(240 d)。说明相较施肥坑,较贫瘠的非施肥坑更易于凋落叶的分解。
3.4 不同林地环境凋落叶分解状况
橡胶凋落叶在不同林地中的分解速率,以橡胶凋落叶在该林地非施肥坑(保根+去根)中0~30 cm土层的平均残留率来表示,结果见图4。

由图4可知,林地土壤环境对凋落物分解的影响表现出一定的时间阶段性,在分解开始的120 d内,4个不同林地凋落叶分解速度差异不明显,残留率均以12 a林地最大,6 a林地最小;120 d后,6 a和28 a林地橡胶凋落叶分解速度开始显著增大,残留率显著低于12 a和18 a林地,其中又以28 a林地凋落叶残留率下降幅度最大;至240 d,4个林地平均残留率分别为2.94%、7.05%、14.62%和2.18%。可见,林地土壤环境可显著影响橡胶凋落叶的分解进程,而这可能与土壤生物及土壤环境(肥力)有关。
3.5 橡胶根系对其凋落叶分解的影响
橡胶根系对其凋落叶分解的影响分别以4个林地施肥坑和非施肥坑中的保根和去根处理的平均残留率表示,结果见图5。

由图5可知,0~60 d,在施肥坑和非施肥坑中,保根处理相较于去根处理均加速了凋落叶的分解,分解速率分别提高了1.47%和2.00 %。此后,根系的促进效应逐渐减弱并最终转变为抑制效应,并于180 d时达到根系最强抑制作用,此时施肥坑和非施肥坑中保根处理分解速率比去根分解速率分别低3.33%和9.02%。整个分解过程中,不同土壤肥力下的根系对凋落物分解的影响均没达到显著性水平,说明相比于土壤肥力对橡胶凋落叶分解速率的决定性影响,橡胶根系对凋落物分解的影响为次一级影响因素。
4 结论与讨论
4.1 不同孔径网袋内橡胶凋落叶残留率变化
分解结束时,0.07 mm孔径网袋内橡胶凋落叶残留率较1.00 mm孔径网袋低7.11%,即土壤中小型动物对凋落物分解的贡献在7%左右。这明显低于González等[15]的研究,即土壤动物对热带雨林地区凋落物分解的贡献可达66%,也低于西双版纳季节雨林大中型土壤动物对凋落物分解46%的贡献[8]。这可能与所选用的网袋孔径大小有关。由于本研究所使用的大孔径网袋孔径大小仅为1.00 mm,限制了很大一部分大中型土壤动物的进入,不可避免地降低了土壤动物的贡献值,而前人所使用的网袋孔径多为1.80 mm或2.00 mm,一方面有利于更大体径动物的进入,另一方面也会加大凋落物碎屑的渗漏(包含在土壤动物贡献之中)。另外,橡胶林属于人工林,相比于自然状况下的热带雨林,植被群落结构单一,土壤动物的类群数量可能不如后者,导致对凋落物分解的贡献也随之降低。
4.2 不同土壤肥力中橡胶凋落叶残留率变化
橡胶凋落叶前期分解速率在非施肥坑和施肥坑中基本一致,而后期非施肥坑中分解速率明显快于施肥坑。因为埋于土壤中的凋落物相较于周围土壤有近似于“培养基”的作用,新近的凋落物有大量可供微生物代谢利用的优质能源物质,这一阶段微生物生长繁殖受土壤养分含量影响较小,且该阶段凋落物损失主要为淋溶损失,因而,橡胶凋落叶在施肥坑和非施肥坑中前期分解速率差异不大。而后,随着易分解物质的耗尽,难分解物质的积累,加上外来新鲜压青肥及化肥的施入,尤其是集中大量施加N、P、K等化学肥料后,会引起施肥坑内网袋中的微生物大量死亡和外迁寻求更可口的食物[16],导致其内的凋落物分解速率开始减慢;对于非施肥坑,虽然凋落物品质也随时间推移不断降低,但养分含量和生存环境仍始终优于周围土壤,并经过一段时间的适应后,适宜于低品质凋落物分解的微生物开始生长增殖,因而,总体分解速度并没有因此而减缓。黄靖宇等[9]也发现,相同类型凋落物在有机质背景低的土壤中分解更快,这是因为土壤微生物从贫瘠土壤吸收的可利用能量要少于相对肥沃的土壤,为了最大化的繁衍种群,土壤微生物会尽可能的吸收环境中的可利用碳源。但王进[10]发现,随土壤肥力的增加,凋落物的分解速率加快,尤其是对低品质凋落物类型这种影响更为显著,这可能与土壤养分高、中、低等级的划分及土壤微生物数量与组成结构不同有关。
4.3 不同埋层中橡胶凋落叶残留率变化
与置于地表凋落物分解速率相比,埋于非施肥坑中的橡胶凋落叶分解速率明显较快,但埋于施肥坑中的橡胶凋落叶分解速率只在前期快于地表,后期分解速率与地表基本持平。马祥庆等[17]和章志情等[18]也发现,埋置处理明显加速了杉木凋落叶分解,并认为导致这一差异是由于受光照影响,地下土壤水热条件要好于地表,有利于凋落物的分解。另外,本研究认为,埋于地下的凋落物与土壤接触面积更大,土壤生物参与凋落物分解的机会也更大,因此,在环境及生物联合作用下,地下组凋落物分解速率比地上组更快。但也有研究得出相反的结论,认为地表受到降水淋溶作用及其他外部环境的影响要好于地下,导致凋落物分解速率埋置处理前期显著低于地表放置,后期由于淋溶作用影响减弱,地下组和地上组分解速率无显著差异[5],因此,埋置处理是否加速凋落物分解可能还与区域气候条件有关。需要注意的是,经240 d的分解后,埋于施肥坑中的橡胶凋落叶分解速率明显降低,残留率与地表基本相同。这可能与施肥坑中人为集中大量施加N、P、K等化肥影响微生物活动及酶活性有关。有研究表明,微生物的数量和酶的活性只有在适宜的氮含量范围内才能发挥正效应,高浓度氮对后期凋落物分解具有明显的抑制作用。可见,埋置处理对凋落物分解的影响与区域气候环境、凋落物类型及土壤肥力密切相关。
4.4 不同林地环境下橡胶凋落叶残留率变化
基于橡胶凋落叶野外原位分解研究发现,观测结束时,不同林龄林地埋置处理橡胶凋落叶残留率以6、28 a林龄林地显著低于12 a和20 a林龄。林地环境对凋落物分解的影响在多种森林类型中均有体现,逯军峰等[19]研究指出,随林龄增长,凋落物分解速率增加;适度间伐可以减小油松(Pinustabulaeformis)木质素/N和C/N比,有利于凋落物分解[20];不同密度巨桉林凋落物分解速率表现为:833株/hm2>1 333株/hm2>2 222株/hm2[21]。不同林龄橡胶林由于其林下植被和土壤环境存在差异[22],可能是导致不同林地凋落物分解速率差异的主要原因。
4.5 不同根系处理下橡胶凋落叶残留率变化
橡胶根系对凋落物分解的影响在分解前60 d表现为促进作用,此后,这种促进效应逐渐转变为抑制效应。张科[23]也发现,在分解的0~60 d,种植植物加快了凋落物的分解,而后这种促进效应逐渐被其抑制效应所代替,至分解结束时(120 d),种植植物比不种植植物多分解6.8%。这是因为前期根系分泌物促进了微生物对凋落物的分解,而后期,根系促进微生物活动的效应,逐渐被根系吸水所产生的抑制微生物的生境效应所掩盖。同样,对于橡胶林,每年1—2月为其落叶高峰期,直到3—4月才长出第一蓬叶,因此,这一阶段为一年中蒸腾作用最小时期,根系吸水损失也最小;5月后,橡胶林郁闭度进入稳定期,温度也维持在较高水平,蒸腾作用旺盛,根系吸水损失增大。但值得注意的是,如果仅仅是因为根系吸水导致凋落物湿度降低而抑制其分解速率的话,本试验所在地区1—2月为旱季,土壤含水率应较雨季(4—10月)低,地下组凋落物湿度也相应更低也更不利于其分解才对。因此,是否是因为橡胶根系吸水而抑制了凋落物后期分解速率还有待进一步研究。
凋落物分解往往是一个长期而又复杂的过程,不同分解阶段分解特性具有很大的不同。本研究受客观因素限制,只研究了橡胶凋落叶在不同环境下1 a内的分解过程,研究结果能够反应橡胶凋落叶阶段性的分解特征,但还不足以表征整个分解过程。因此,今后关于橡胶凋落叶跨年度全周期分解研究还有待进一步进行。
[1] Xi Z B. Approach to slow release nitrogen fertilizer and its agrochemical evaluation[J]. Phosphate and Compound Fertilizer , 2003,18(5):1-5.
[2] 皮荷杰,曾清如,蒋朝晖,等. 两种硝化抑制剂对不同土壤中氮素转化的影响[J].水土保持学报,2009,23(1): 68-72.
[3] 温明章,郭继勋.不同凋落物量对东北羊草草原土壤生物的影响[J].中国草地学报,2008,30(5):7-12.
[4] 王星丽,殷秀琴,宋博, 等. 羊草草原主要凋落物分解及土壤动物的作用[J].草业学报,2011,20(6): 143-149.
[5] 李海涛,于贵瑞,李家永,等. 井冈山森林凋落物分解动态及磷、钾释放速率[J].应用生态学报,2007,18(2): 233-240.
[6] 刘海岗,刘一,黄忠良. 森林凋落物研究进展[J].安徽农业科学,2008,36(3): 1018-1020.
[7] Berg B,Johnansson M B,Meentemeyer V. Litter decomposition in a transect of Norway spruce forests: substrate quality and climate control[J]. Canadian Journal of Forest Research,2000,30(7): 1136-1147.
[8] 杨效东,邹晓明. 西双版纳热带季节雨林凋落叶分解与土壤动物群落:两种网孔分解袋的分解试验比较[J].植物生态学报,2006,30(5): 791-801.
[9] 黄靖宇,宋长春,张金波,等. 凋落物输入对三江平原弃耕农田土壤基础呼吸和活性碳组分的影响[J].生态学报,2008,28(7): 3417-3424.
[10] 王进. 土壤基质与凋落物分解互作效应的研究[D].武汉:华中农业大学,2014.
[11] 邹文涛,刘湘洪,孙娟. 中国天然橡胶“走出去”战略: 形势、问题与对策[J].世界农业,2014,(12): 157-161.
[12] 任泳红,曹敏,唐建维, 等. 对西双版纳季节雨林与橡胶多层林凋落物动态的比较研究[J]. 植物生态学报,1999,23(5):418-425.
[13] 赵春梅,曹建华,李晓波,等. 橡胶林枯落物分解及其氮素释放规律研究[J].热带作物学报,2012,33(9): 1535-1539.[14] 曹建华,陶忠良,赵春梅,等. 不同年龄橡胶树枯落物养分归还比较[J]. 热带作物学报,2011,32(1):1-6.
[15] González G,Seastedt T R. Soil fauna and plant litter decomposition in tropical and subalpine forests[J]. Ecology,2001,82(4): 955-964.
[16] Sun B J,Zhang X P,Jia S X. The effect of soil physical an chemical properties on soil microbial community in agro ecosystem[J]. Soil and Crop,2013,2( 3) : 138-144.
[17] 马祥庆,刘爱琴,何智英. 杉木幼林生态系统凋落物及其分解作用研究[J].植物生态学报,1997,21(6): 564-570.
[18] 章志琴,林开敏,邹双全,等. 不同调控措施对杉木枯落物分解的影响[J].浙江农林大学学报,2006,23(1): 65-69.
[19] 逯军峰,王辉,曹靖,等. 油松人工林凋落物对土壤理化性质的影响 [J].西北林学院学报,2007,22(3): 25- 28.
[20] 徐扬. 抚育间伐对油松人工林凋落物分解影响的研究[D].北京:北京林业大学,2007.
[21] 向元彬. 华西雨屏区不同密度巨桉人工林凋落物分解、养分释放及其土壤呼吸特征[D]. 雅安:四川农业大学,2010.
[22] 周会平,岩香甩,张丽谦,等. 西双版纳橡胶林下植被多样性调查研究[J].热带作物学报,2012,33(8): 1444-1449.
[23] 张科. 森林凋落叶分解受不同来源土壤和植物生长的影响[D].北京:中国农业大学,2005.
(责任编辑 赵粉侠)
Study on Decomposition of Rubber Leaf-Litter with Different Control Measures
Jin Long1,2, Wu Zhixiang1,2, Yang Chuan1, Guan Limin1, Lai Huaying1
(1.Rubber Research Institute,Chinese Academy of Tropical Agricultural Sciences, Danzhou Investigation & Experiment Station of Tropical Crops,Ministry of Agriculture,Danzhou Hainan 571737, China;2. College of Environment and Plant Protection, Hainan University, Haikou Hainan 570228, China)
Leaf-litter of 6-year-old,12-year-old,20-year-old and 28-year-old rubber plantations was used to studied the decomposition progress in one year under different control measures by mesh bag method. The results as following: leaf-litter in 0.07 mm and 1.0 mm mesh bags was in a same decomposition dynamic, which was increased first and then drop, and the litter residues rates at the end of year were 36.84 % and 32.09 %, respectively. Leaf-litter decomposition rates not only by the position of the buried layer influence, but also with soil fertility-related. After 240 d, litter residues of ground plane (0 cm), comparison hole (0-30 cm) and fertile hole (0-30 cm) not included roots removed were 32.09%, 7.40% and 34.58%, respectively. Litter decomposition of rubber was reflected by soil fertility mainly focus on the late stage of decomposition. To 240 d, residues of litter in comparison hole and fertile hole (the average value of the group of roots reserved and roots removed) were 8.46% and 35.62%, respectively. The difference of litter decomposition rates of 6-year-old,12-year-old,20-and 28-year-old rubber plantations were little at early stage but significant at late stage. To 240 d, residues of litter were 2.94%, 7.05%, 14.62% and 2.18%, respectively. Although rubber roots promoted litter decomposition at early stage, and convert to inhibition at the late stage, no significant influence was detected about rubber roots on litter decomposition at all stages.
rubber plantation; control measures; leaf litter; decomposition rate
2015-03-23
中国热带农业科学院橡胶研究所基本科研业务费专项(1630022014011)资助。
吴志祥(1970—),男,副研究员。研究方向:热带作物栽培和农业气象生态。Email:zhixiangwu@21cn.com。
10.11929/j.issn.2095-1914.2015.05.004
S714.5
A
2095-1914(2015)05-0021-06
第1作者:金龙(1987—),男,硕士生。研究方向:热带作物栽培生态。Email: jl812273236@163.com。
