缢蛏(Sinonovacula constricta)EST-SSR标记与生长性状的相关性分析*
2015-03-22邵艳卿张炯明肖国强滕爽爽柴雪良
邵艳卿 方 军 柏 艳, 张炯明 肖国强滕爽爽 刘 博 柴雪良①
(1. 浙江省海洋水产养殖研究所 浙江省近岸水域生物资源开发与保护重点实验室 温州 325000;2. 温州医科大学 温州 325000)
缢蛏(Sinonovacula constrictaLamarck)俗称蛏(福建)、蜻(浙江)或跣(我国北方), 其肉味鲜美, 营养丰富, 含有丰富的蛋白质、维生素和无机盐。除供鲜食外, 还可制成蛏干、蛏油等, 是消费者喜爱的海产食品, 也是我国传统四大养殖贝类之一(王如才等,2008)。缢蛏养殖历史悠久, 过去主要集中在福建和浙江一带, 随着缢蛏养殖业的不断扩展和人工育苗技术日渐成熟, 江苏、山东等地纷纷引进苗种, 有力地推动了缢蛏养殖业的发展。然而, 目前人工苗的亲蛏绝大多数来自人工养殖, 其种质属于未经遗传改良的野生种, 在经过多年累代繁、养殖之后, 出现了成活率低、生长速度减慢和抗病力下降等生产性状退化现象; 加之, 无序的养殖、盲目引种和养殖环境的日益恶化, 已经严重制约了缢蛏养殖业的健康持续发展(王兴强等, 2006; 刘博等, 2013)。因此, 采用有效方法培育出高产、抗逆的缢蛏优良品种成为突破瓶颈、带动产业快速发展的当务之急。
利用分子遗传标记进行标记辅助选育, 是当今动物遗传育种研究的热点之一。微卫星也称简单重复序列(simple sequence repeat, SSR), 是由1-6个核苷酸组成的简单串联重复DNA序列(Tautz D, 1989), 按照其来源可分为基因组 SSR(G-SSR)和表达序列标签SSR(EST-SSR)。微卫星标记具有数量多、多态性丰富、共显性遗传、重复性好等优点(Zaneet al, 2002; 刘芳等, 2006), 在筛选水产动物形态、抗病、抗逆、性别等经济性状相关标记方面已被广泛使用, 如在虹鳟(Oncorhynchus mykiss)(Rodriguezet al, 2004)、鲤鱼(Cyprinus carpio)(张义凤等, 2008; 顾颖等, 2009)、南美白对虾(Litopenaeus vannamei)(Zhanget al, 2007)、马氏珠母贝(Pinctada martensii)(邓岳文等, 2013)、文蛤(Meretrix meretrix)(Luet al, 2013; Nieet al, 2013)等都已有相关报道, 而缢蛏在性状相关标记筛选方面则还未见报道。本研究首先构建缢蛏全同胞家系, 进而采用 SSR技术对其分子特征进行检测, 并结合其数量性状度量, 筛选与缢蛏生长性状显著相关的分子标记, 以期为缢蛏生长性状的QTLs定位、分子标记辅助育种和新品种培育提供参考依据。
1 材料与方法
1.1 实验材料
所用缢蛏样品为2011年10月在浙江省海洋水产养殖研究所清江基地培育的全同胞(F1)家系(包含父母本样品), 养殖一龄后随机取120颗, 测量壳长、壳高、体重等指标, 解剖取肌肉组织, 固定于90%乙醇中备用。
1.2 DNA提取、PCR扩增及检测
基因组 DNA 采用常规的苯酚/氯仿/异戊醇法抽提, 用紫外分光光度计测定浓度, 再用无菌水统一调至100ng/µL, –20°C保存备用。PCR反应体系总体积为25µL, 包括10×PCR buffer 2.5µL, 25mmol/L MgCl22.5µL, 2.5mmol/L dNTPs 2µL, 上下游引物(10µmol/L)各 1.0µL, 5U/µL Taq DNA 聚合酶(TaKaRa)0.13µL,DNA模板 1.0µL, 加灭菌双蒸水至 25µL。PCR扩增程序为: 94°C预变性5min, 进入30个PCR循环[94°C 30s, 退火温度(表 1) 30s, 72°C 30s], 最后 72°C 下延伸7min。扩增产物经6%的变性聚丙烯酰胺凝胶电泳检测, 硝酸银染色。
本实验所用引物来自刘博等(2012)已发表的和后续自主设计开发。所有引物均由上海生工生物工程有限公司合成, 根据亲本筛选出 16对多态性引物用于后续研究(表1)。
1.3 数据分析与统计
根据电泳图谱, 并参照引物设计时预期片段的大小判读、统计条带。用PopGene (Verion3.2)软件计算各位点在子代中的等位基因频率(allele frequency)、等位基因数(observed number of alleles,Na)、有效等位基因数(effective number of alleles,Ne)和观测杂合度(observed heterozygosity,Ho); 根据亲本的基因型推算各位点在子代的期望杂合度(expected heterozygosity,He); 根据Botstein等(1980)的公式计算出多态性信息含量(PIC)。
用SPSS19.0软件中的卡方检验分析各位点在子代中的分离比是否符合孟德尔遗传; 用一般线性模型(general linear model, GLM)对SSR位点基因型与主要生长性状(壳长、壳宽、壳高和体重)进行相关性分析, 经方差分析检验呈显著性差异的位点, 使用Duncan法进行多重比较。
2 结果与分析
2.1 缢蛏的生长指标分析
随机取缢蛏 F1家系 120个个体, 分别测量其壳长、壳宽、壳高、体重的生长指标, 测量结果见表2,4个生长性状都呈现连续变异的特点, 符合典型的数量性状或多基因遗传特点。通过K-S单样本正态分布检验, 结果显示其壳长、壳宽、壳高与体重的P值分别为0.56、0.30、0.25和0.57, 均符合正态分布(表2),适合作为标记与性状相关分析的材料。
2.2 缢蛏SSR遗传多样性分析及其基因型分布情况
用16个SSR位点对缢蛏亲本及其子代120个个体进行了PCR扩增, 其中有1个子代个体PCR产物电泳条带不够理想, 因此舍弃了该个体。遗传参数统计结果表明(表3): 16个位点共检测到39个等位基因,各位点的等位基因数2—4个, 平均2.44, 平均有效等位基因数为2.09; 观测杂合度在0.36—1.00之间, 平均为0.58; 期望杂合度在0.50—1.00之间, 平均为0.63;多态信息含量(PIC)在0.25—0.70之间, 平均值0.40。
根据亲本和子代的基因型判读统计。16个位点中有 4个位点子代具有 4种基因型, 为 1︰1︰1︰1(BC×AD/AB×BC/AC×BD)的分离类型; 10位点属 1︰1分离类型, 其中4个为雄亲杂合雌亲纯合, 6个为雌亲杂合雄亲纯合; 2个位点属1︰2︰1 (AB×AB)分离类型。根据卡方检验, 有 10个位点(62.5%)的子代基因型分布处于平衡状态(P>0.05), 其它位点(37.5%)都不同程度地偏离了平衡(P<0.05或P<0.01)(表3)。
2.3 缢蛏生长性状相关SSR位点分析与多重比较
对每个 SSR位点的不同基因型进行分类, 结合
表型数据进行关联分析, 结果在 16个 SSR位点中,发现共有 4个位点与缢蛏生长性状表现出一定的相关性, 其中YC-1与壳宽显著相关(P<0.05); YC-93与壳宽、壳高均极显著相关(P<0.01); YC-96与体重显著相关(P<0.05); YC-123位点与壳高、体重均显著相关(P<0.05)。
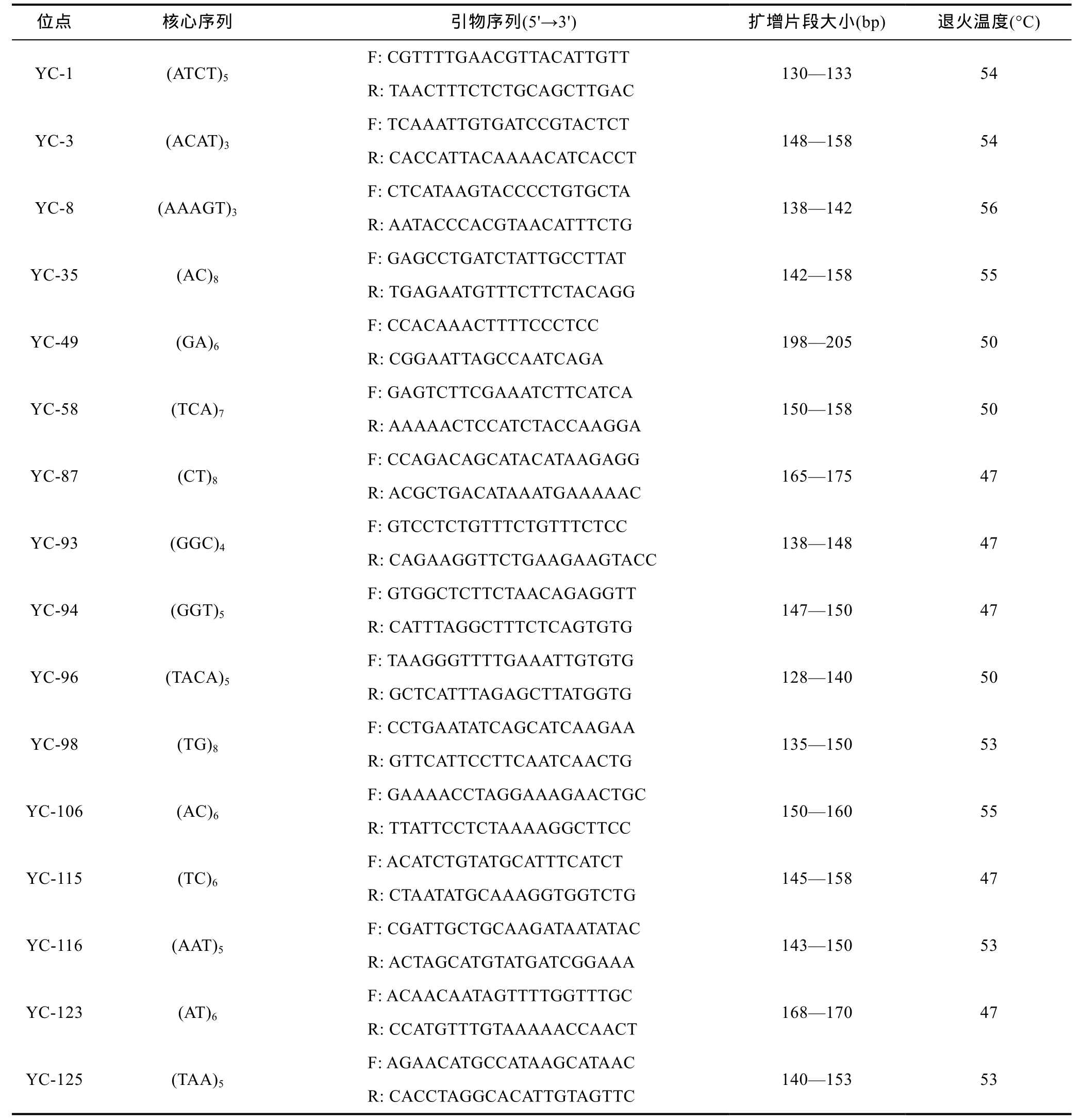
表1 本研究选用的微卫星位点、核心序列、引物序列和退火温度Tab.1 Microsatellite loci, repeat motif sequence, prime sequence and annealing temperature used in this study

表2 壳长、壳宽、壳高、体重的表型数据及正态分布检验Tab.2 The phenotypic data of shell length, width, height, and weight, and normal distribution test
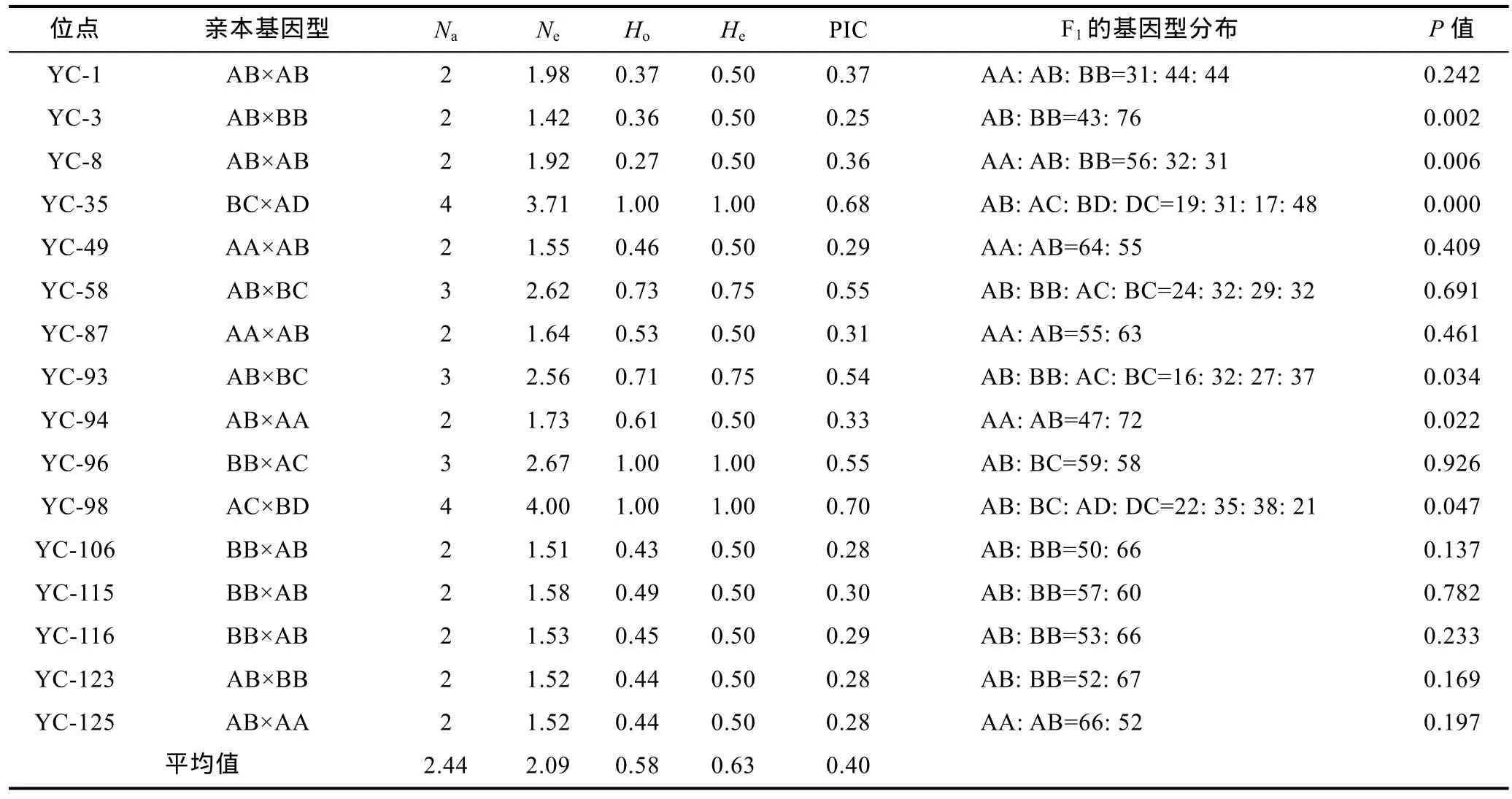
表3 16个微卫星位点在缢蛏F1家系的统计信息Tab.3 Statistic information for 16 microsatellite loci in an F1 family of S. constricta
将这 4个有显著相关的位点进行不同基因型间的不同性状的多重比较(Duncan法), 结果如下(表4):在标记YC-1中, 基因型BB个体的壳长、体重、壳宽值均高于AA和AB个体, 但仅在壳宽性状上呈显著差异(P<0.05), 推测 BB基因型是壳宽的优势基因型, 等位基因A可能起负面影响。
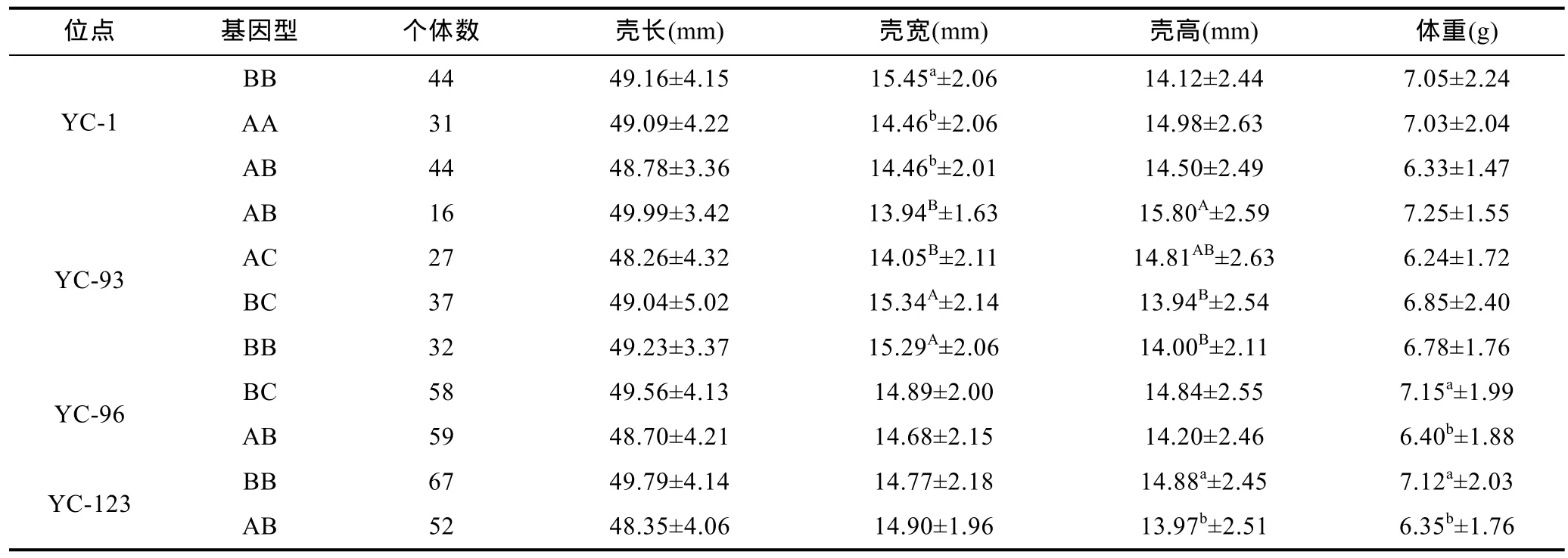
表4 4个微卫星位点不同基因型壳长、宽、高与体重的多重比较Tab.4 Multiple comparisons of major growth traits with different genotypes at 4 microsatellite loci
在标记YC-93中, 基因型BC和BB个体的壳宽值极显著高于AB和AC个体(P<0.01), 说明等位基因A对壳宽起负面影响; 在壳高性状上则是基因型 AB和AC个体极显著或显著高于BC和BB个体(P<0.01或P<0.05), 说明等位基因A对壳高起正面影响; 4种基因型在体重上AB型个体最大, 但与其它3种基因型个体均未达到显著差异水平。
在标记YC-96中, 基因型BC个体的平均壳长、壳宽、壳高、体重值均高于AB个体, 在平均体重性状上差异显著(P<0.05), 可说明 BC基因型是体重性状的优势基因型, 而等位基因A起负面影响。
在标记YC-123中, 基因型BB个体的平均壳高、体重值均显著高于AB个体(P<0.05), 说明基因型BB对壳高、体重起正面影响, 等位基因A起负面影响。
2.4 最优基因型组合筛选
因为体重一般是水产动物最终收获的主要经济性状, 因此, 本研究以体重为参照指标对这4个位点不同基因型组合进行比较。由于4个位点中某些基因型组合出现频率太少, 缺少分析价值, 因此在实际统计分析中, 每种基因型组合至少有3次观察值才被考虑。YC-1、YC-93、YC-96和YC-123四个位点的基因型分别有3、4、2和2种基因型, 可形成48种组合, 因在标记YC-93和YC-96中分别有7个和2个个体未检测到基因型, 因此样本数为 110。在检测的110个个体中实际出现了32种基因型组合, 16种基因型组合的个体没出现。对32种不同基因型组合的体重表型值进行比较, 获得的最优基因型组合为BB/BB(BC)/BC/BB, 与4个位点单独分析对应的最优基因型基本完全一致, 符合加性作用模型(表5)。
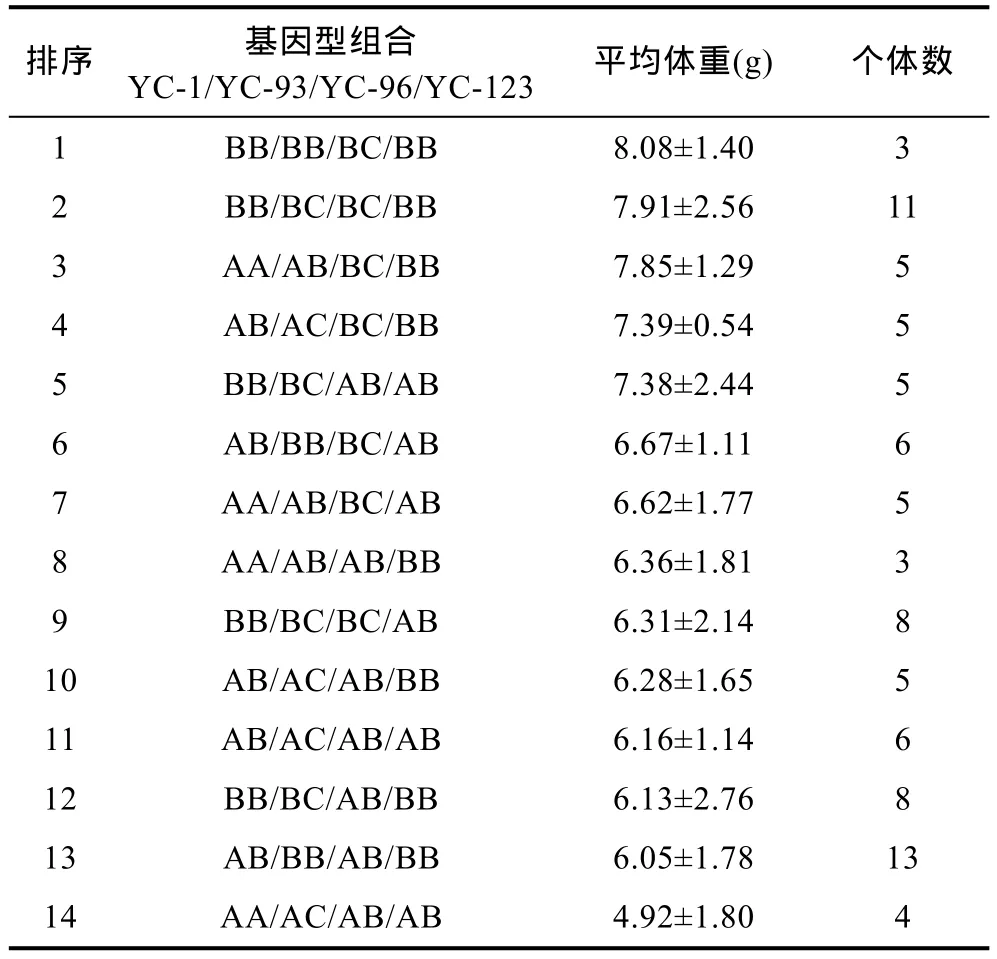
表5 缢蛏F1家系的不同基因型组合对应体重排序统计表Tab.5 The statistics to body weight value with different genotype combinations in an F1 family of S. constricta
3 讨论
3.1 缢蛏家系遗传结构分析
Ne、Ho、He和 PIC是反映群体遗传多样性的重要参数, 数值越大, 表明群体遗传结构越复杂, 进而说明基因丰富度越高(李春艳, 2009)。本文中, 缢蛏家系群体的Ne=2.09,Ho=0.58,He=0.63和PIC=0.40, 可以看出该缢蛏家系的遗传多样性水平处于中等水平,普遍低于已报道的缢蛏不同地理养殖群体和野生群体的遗传多样性水平(牛东红等, 2011; 刘达博等,2011; 刘博等, 2013), 出现这种现象的原因可能与本文研究的对象是全同胞家系群体有关。
另外, 本研究的 16个位点的期望杂合度多数与观测杂合度基本一致, 说明实验中测量缢蛏家系群体的这些位点的基因频率和基因型频率稳定性较好,除了位点YC-8的期望杂合度(He=0.50)明显大于观测杂合度(Ho=0.27), 表现出 F1代个体的分离方式严重偏离孟德尔分离规律(P<0.01)。分析该位点在F1群体中的基因型分布, 发现杂合子 AB个体很少, 仅占全部观测个体的 26.9%(理论上应 50%); 而纯合子 AA个体很多, 占全部观测个体的 47.1%(理论上只占25%)。这提示该位点附近可能存在着不利于杂合子AB个体而利于纯合子 AA个体存活的基因。位点YC-35严重偏离孟德尔分离定律(P=0.000), 其原因是由于基因型AB和BD个体(AB︰AC︰BD︰DC=19︰31︰17︰48, 理论上应 1︰1︰1︰1)出现频率显著偏低, 暗示该位点可能存在与B连锁的基因不利于缢蛏存活, 有待于进一步深入研究。
多态信息含量(PIC)是衡量基因变异程度高低的重要指标, 可以反应出基因座位在群体中的多态性高低。研究利用的16个SSR位点, 据Botstein等(1980)的界定标准, 有 5个位点表现为高度多态位点(PIC>0.50), 11个表现为中度多态位点(0.25 微卫星标记具有保守性好、呈共显性遗传的特点,是近年来发展迅速、应用广泛的分子标记, 也是分析与重要经济性状的遗传连锁关系最理想的标记之一,目前已经被广泛应用于水产动物生长、抗病、性别等性状相关分子标记筛选。例如 Lu等(2013)利用双向选择性分型筛选出3个与文蛤(Meretrix meretrix)生长QTL紧密连锁的 EST-SSR标记, 并用单向选择性分型对其进行了验证; Song等(2012)利用半滑舌鳎(Cynoglossus semiliaevis)F1家系构建高密度遗传连锁图谱, 并进行性状-标记之间的回归分析, 得到2个与生长性状相关(P<0.01)的SSR标记; Guo等(2012)基于全同胞家系构建长牡蛎(Crassostrea gigas)的性别平均连锁图, 检测到3个生长相关的QTLs和一个性别相关的QTL; Rodriguez等(2004)利用雄性硬头鳟和雌性虹鳟(Oncorhynchus mykiss)杂交产生的雄性F1回交雌性虹鳟产生70个家系(BC1), 从每个F1和BC1家系中取近100个体进行IHN病毒感染, 表型性状采用1(感染后死亡)和0(感染后存活), 筛选到6 个微卫星标记和IHNV抗性有关, 为进一步将与抗性有关的基因进行定位、克隆奠定基础。在本实验中, 共找到了4个与生长性状显著相关的 EST-SSR标记, 其中YC-93与缢蛏的壳宽、壳高性状极显著相关, YC-123与壳高、体重性状显著相关, 而位点YC-1和YC-96仅与壳宽或体重一个性状显著相关, 可初步确定这些位点为与生长相关的候选标记; 尤其是位点YC-96的BC基因型和YC-123的BB基因型的个体体重性状均显著高于同一标记的其它基因型的个体, 推测这两个基因型为体重优势基因型, 可用于对缢蛏体重性状进行辅助选育。 后续的研究工作中, 将对筛选到的这些候选标记进行进一步的分析与验证, 探讨其能否作为缢蛏标记辅助育种的有效分子标记。 王兴强, 曹 梅, 阎斌伦, 2006. 缢蛏Sinonovacula constricta(Lamarck)养殖期间发病原因及防治对策. 现代渔业信息,21(5): 13—16 王如才, 王昭萍, 2008. 海水贝类养殖学. 青岛: 中国海洋大学出版社, 231 牛东红, 冯冰冰, 刘达博等, 2011. 浙闽沿海缢蛏群体遗传结构的微卫星和线粒体COI序列分析. 水产学报, 35(12):1805—1813 邓岳文, 高远镇, 王学颖等, 2013. 马氏珠母贝生长性状与EST-SSR标记的关联分析. 农业生物技术学报, 21(1):77—88 刘 芳, 李卫东, 王 强等, 2006. 微卫星标记及其在贝类遗传选育研究中的应用.水产科学, 25(5): 268—270 刘 博, 邵艳卿, 王 侃等, 2013. 4个缢蛏群体遗传多样性和系统发生关系的微卫星分析. 海洋科学, 37(8): 96—102 刘 博, 邵艳卿, 滕爽爽等, 2012. 缢蛏(Sinonovacula constricta)EST-SSR分布特征及引物开发利用. 海洋与湖沼, 43(1): 132—137 刘达博, 牛东红, 冯冰冰等, 2011. 乐清湾和三沙湾缢蛏群体遗传多样性的微卫星分析. 上海海洋大学学报, 20(3):350—357 李春艳, 丁 君, 常亚青等, 2009. 虾夷扇贝微卫星标记的分离及其养殖群体的遗传结构分析. 中国水产科学, 16(1):39—46 张义凤, 张 研, 鲁翠云等, 2008. 鲤鱼微卫星标记与体重、体长和体高性状的相关分析. 遗传, 30(5): 613—619 顾 颖, 曹顶臣, 张 研等, 2009. 鲤与生长性状相关的EST-SSRs标记筛选. 中国水产科学, 16(1): 15—22 Botstein D, White R L, Sckolnick Met al, 1980. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. The American Journal of Human Genetics, 32(3): 314—331 Guo X, Li Q, Wang Q Zet al, 2012. Genetic mapping and QTL analysis of growth-related traits in the Pacific oyster. Marine Biotechnology, 14(2): 218—226 Lu X, Wang H X, Liu B Zet al, 2013. Three EST—SSR markers associated with QTL for the growth of the clamMeretrix meretrixrevealed by selective genotyping. Marine Biotechnology, 15(1): 16—25 Nie Q, Yue X, Chai X Let al, 2013. Three vibrio-resistance related EST-SSR markers revealed by selective genotyping in the clamMeretrix meretrix. Fish & Shellfish Immunology,35(2): 421—428 Rodriguez M F, LaPatra S, Williams Set al, 2004. Genetic markers associated with resistance to infectious hematopoietic necrosis in rainbow and steelhead trout (Oncorhynchus mykiss) backcrosses. Aquaculture, 241: 93—115 Song W T, Li Y Z, Zhao Y Wet al, 2012. Construction of a high-density microsatellite genetic linkage map and mapping of sexual and growth-related traits in half-smooth tongue sole (Cynoglossus semilaevis). PLoS One, 7(12): e52097 Tautz D, 1989. Hypervariability of simple sequences as a general source for polymorphic DNA markers. Nucleic Acids Research, 17(16): 6463—6471 Zane L, Bargelloni L, Patarnello T, 2002. Strategies for microsatellites isolation: a review. Mol Ecol, 11: 1—16 Zhang L S, Yang C J, Zhang Yet al, 2007. A genetic linkage map of Pacific white shrimp (Litopenaeus vannamei):sex-linked microsatellite markers and high recombination rates. Genetica, 131(1): 37—493.2 缢蛏生长性状相关分子标记的筛选
