许氏平鲉(Sebastes schlegelii)EST-SSR标记开发及通用性检测*
2015-03-22马海涛韩承慧孙国华姜海滨
薛 蕊 马海涛 韩承慧 王 斐 孙国华 姜海滨①
(1. 上海海洋大学水产与生命学院 上海 201306;2. 山东省海洋资源与环境研究院 山东省海洋生态修复重点实验室 烟台 264006)
微卫星标记是以 2—6个核苷酸单位组成的串联重复序列, 存在于绝大多数真核生物基因组中, 因此又称简单序列重复(Simple sequence repeats, SSRs)和短串联重复(Simple tandem repeats, STRs)(Tautzet al,1984)。它由核心序列和侧翼序列两部分构成, 核心序列高度多态, 而侧翼序列则一般高度保守。相较于以往的分子标记, 微卫星标记具有数量丰富、操作简便、重复性好、共显性遗传等优点(Schuget al, 1998),因此已成为水产动物种群遗传多样性分析、亲缘关系鉴定和遗传图谱构建等方面的有效工具(Chistiakovet al, 2006)。
EST(Expressed Sequence Tag)即表达序列标签,是一段通过单向测序得到的200—250 bp核苷酸序列(侯战辉等, 2008)。由于来源于cDNA文库, 因此EST是功能基因的一部分, 可以对其进行基因注释(邱樱,2013)。EST-SSR即存在于EST序列上的微卫星标记,与传统的基因组微卫星标记(Genomic-SSR)相比,EST-SSR标记开发成本低, 可以节省大量的人力物力,并且在不同物种间具有一定的通用性(易少奎等,2013; 董迎辉等, 2013), 因此已经成为新型高效的开发SSR途径。
许氏平 鲉(Sebastes schlegelii)又称黑, 俗称黑寨、黑老婆等, 是分布在西北太平洋近岸的温水性岩礁栖息鱼种, 广泛存在于黄海、渤海近海海域, 为卵胎生鱼类。由于其肉质鲜美、营养丰富, 而且生长较为迅速, 已成为我国北方深水网箱养殖的重要经济鱼种。进行许氏平 鲉的良种选育, 既可保护自然资源,又可提高养殖效率, 意义重大, 而开发具有良好多态性的微卫星标记是进行遗传育种的基础。已有研究者采用 传统方法开发了一些许氏平 鲉 Genomic-SSR标记(Yoshidaet al, 2005; Anet al,2009; Baiet al, 2011;Yasuikeet al, 2013), 但对EST-SSR标记的开发未见报道。本研究首次利用已发表的许氏平 鲉 EST序列进行EST-SSR标记开发, 分析了与Genomic-SSR标记的差异, 并检测了其在近缘种朝鲜平 鲉(Sebastes koreanus)、褐菖鲉(Sebastiscus marmoratus)的通用性,以期为许氏平鲉良种选育提供条件。
1 材料与方法
1.1 样品采集及DNA提取
许氏平 鲉野生群体样品30个取自长岛附近海区,朝鲜平 鲉 样品8个取自大连, 褐菖鲉样品8个取自威海乳山。尾鳍和肌肉样品暂存放在70%乙醇中, 带回实验室后于–20°C 冰箱保存。采用传统酚/氯仿/异戊醇法抽提基因组 DNA, 用 1%琼脂糖凝胶电泳检测DNA完整性、NanoDrop2000紫外分光光度计检测DNA浓度和纯度。DNA样品保存于–20°C冰箱中, 实验前进行稀释, 终浓度为50ng/μL。
1.2 EST序列获取及分析
从 NCBI公共数据库(http: //www.ncbi.nlm.nil.gov/sites/entrez)下 载许氏平 鲉EST序列(检索词为Sebastes schlegelii)。对序列进行拼接后利用 SSR Hunter 1.3软件查找微卫星序列, 查找条件为: 二碱基重复5次以上(含5次); 三碱基重复4次以上(含4次); 四碱基、五碱基和六碱基重复均在 3次以上(含3 次)。
1.3 EST-SSR引物设计
用Primer Premier 5.0软件设计引物。引物长度控制在18—22bp; GC含量控制在40%—60%, 正反引物相差不超过10%; Tm值控制在40—60°C, 正反引物相差不超 10°C; 产物长度控制在 100—500bp。设计好的引物送生工生物(上海)有限公司合成。
1.4 EST-SSR引物筛选与优化
实验在Eppendorf普通梯度PCR仪中进行。第一步用3个DNA样品混合进行初步筛选, 选出能够稳定扩增目的条带的 EST-SSR引物; 第二步对扩增效果不理想的引物进行梯度优化。实验用酶为TIANGEN Taq DNA Polymerase ET101-02-04, 25μL反 应 体 系 如 下: 2μL DNA 模 板(50ng/μL), 2.5μL 10×buffer, 引 物 各 1μL (50μmol/L), 0.5μL dNTP(10mmol/L), 0.2μL Taq DNA 聚合酶(5U/μL), 剩余体积以灭菌水补齐。PCR程序设为: 94°C预变性5min,接着进行30个循环: 94°C 变性45s, 退火45s, 72°C延伸 1min, 循环结束后 72°C延伸 10min。扩增产物进行8%(W/V)非变性聚丙烯酰胺凝胶电泳、银染染色,并用扫描仪拍照。
1.5 野生群体多态性检测
筛选出的EST-SSR引物进行野生群体PCR扩增,反应体系和扩增程序同引物筛选部分, 不同引物的退火温度参见表1。扩增产物进行8% (W/V)非变性聚丙烯酰胺凝胶电泳、银染染色、扫描仪拍照。
1.6 多态性微卫星引物通用性检测
用多态性微卫星引物对朝鲜平 鲉 和褐菖 鲉DNA样品进行 PCR扩增, 反应体系和扩增程序同引物筛选部分, 不同引物的退火温度参见表 1。扩增产物进行8% (W/V)非变性聚丙烯酰胺凝胶电泳和银染染色,扫描仪拍照。
1.7 数据统计
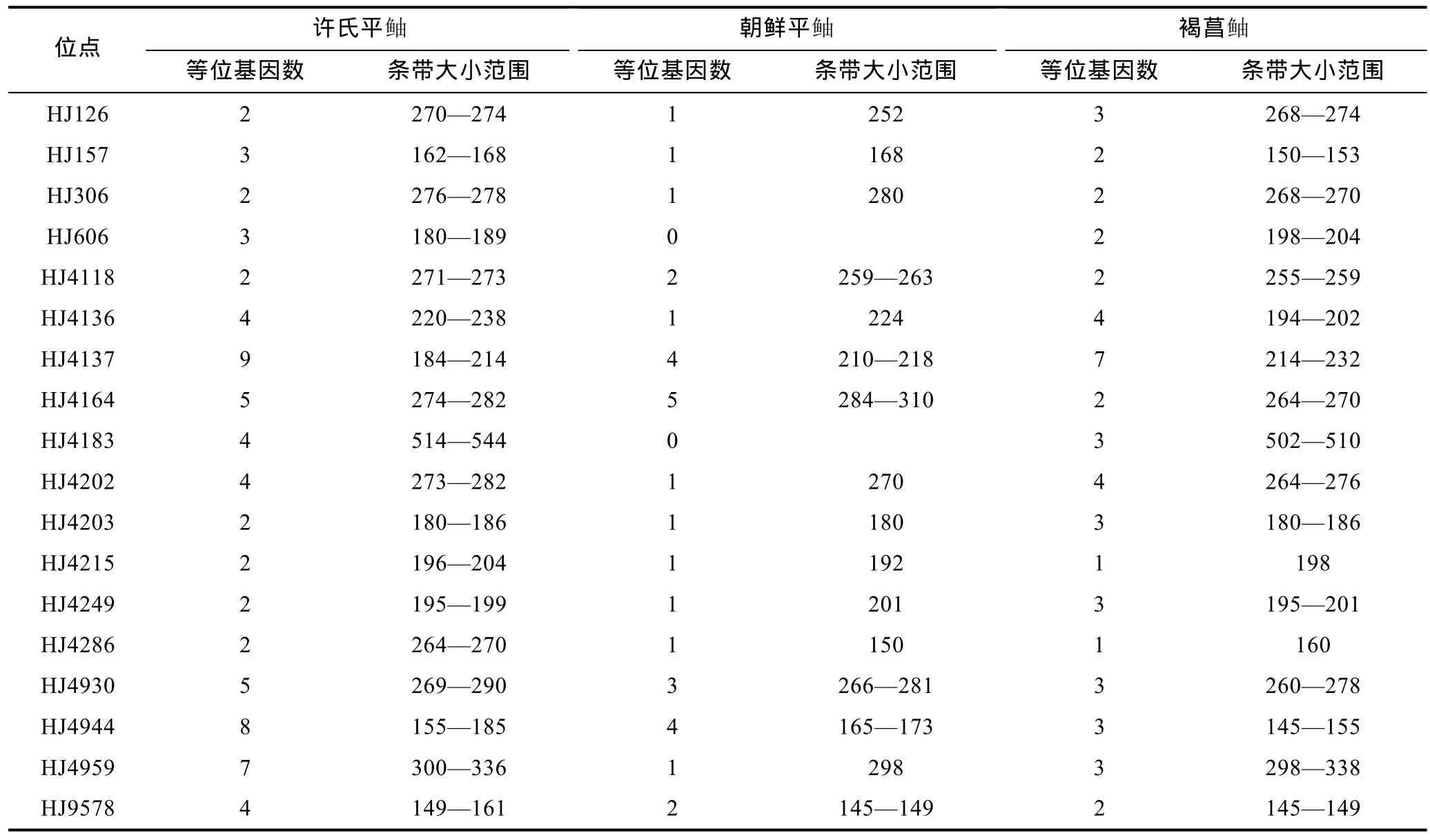
统计各SSR位点DNA带型, 估算条带分子量大小, 统计结果输入POPGENE 32 (Yehet al,1999)计算每个位点的等位基因数(Na)、观测杂合度(Ho)、期望杂合度(He)和香农多样性指数(I)。用软件CERVUS 3.0(Marshallet al, 1998)计算多态信息含量(Polymorphic information content, PIC)。GENEPOP v.4.1.4(Rousset,2008)检验Hardy-Weinberg平衡和连锁不平衡情况。另外统计18对多态EST-SSR 引物对朝鲜平 鲉和褐菖鲉个体的通用率和多态率, 分析各位点通用性情况。
2 结果与分析
2.1 许氏平鲉 EST序列中SSR位点种类、数量及出现频率
从NCBI公共数据库下载的1980条EST序列经拼接、SSR位点查找后, 发现 181条 EST序列含有SSR位点, 共计 224个, EST-SSR位点的发现率为9.14%, 平均每条EST序列上有SSR位点1.24个。其中, 含有一个EST-SSR位点的144条, 两个EST-SSR位点的33条, 三个及三个以上EST-SSR位点的4条。依据Weber(1990)提出的分类方法, 所有EST-SSR位点中完美型比例占到 91.52%, 二到六碱基重复均有出现, 位点个数分别为 109、88、4、2和 2, 分别对应百分比53.17%、42.93%、1.95%、0.98%以及0.98%。其中, 二碱基重复出现频率最高, 分为 10种重复类型, 以 TG/CA基序最多, 占 33.03%, 占总完美型位点的 17.56%; 三碱基重复有 23类, 以 CAG/CTG出现频率最高, 为 12.50%, 四到六碱基所有重复类型皆出现一次。

表1 18对许氏平鲉 EST-SSR引物特征Tab.1 Characteristics of 18 pairs of EST-SSR primers for S. schlegelii
2.2 引物筛选和多态性检测结果
对查找到的224个EST-SSR位点进行进一步分析, 挑选出部分适合设计引物的位点, 共设计引物56对, 筛选后共计 46对引物能够进行有效扩增, 野生群体多态性检测结果显示, 其中 18个位点具有多态性(表1), 位点HJ4137的电泳图见图1。
2.3 EST-SSR位点在野生群体中的多态性评价
所有位点共检测出70个等位基因, 每个位点观测等位基因数为 2—9, 平均观测等位基因数为3.89。观测杂合度(Ho)在0.0333—0.8000之间, 平均观测杂合度为 0.3037; 期望杂合度(He)在 0.0333—0.7927之间, 平均期望杂合度为0.3757; 香农指数(I)在0.0848—1.7819之间, 平均香农指数为0.7416; 每个位点的多态信息含量(PIC)在0.0323—0.7522之间,平均多态信息含量为 0.3419, 其中高度多态(PIC>0.5)位点5个, 中度多态(0.25 图1 位点HJ4137的PCR扩增结果Fig.1 The result of PCR amplification of locus HJ4137 表2 18个许氏平鲉 EST-SSR位点的遗传多样性参数Tab.2 Genetic diversity parameters of 18 EST-SSR loci for S. schlegelii 在朝鲜平 鲉中除位点HJ606和HJ4183其余位点均能有效扩增, 其中6个位点多态; 在褐菖 鲉中所有位点均能有效扩增, 除位点HJ4215和HJ4286单态外其余位点均为多态。18 个位点在朝鲜平 鲉 和褐菖鲉中的通用率分别是88.89%和100%, 多态率为33.33%和88.89%。其中, 位点 HJ4215和HJ4286在许氏平鲉中显示多态而在朝鲜平 鲉 、褐菖 鲉个体中为单态;位点 HJ4136、HJ4202以及 HJ4959 对许氏平 鲉、褐菖 鲉显示多态而在朝鲜平 鲉中为单态; 位点HJ606和HJ4183 对许氏平 鲉、褐菖 鲉 显示多态而在朝鲜平鲉中不能有效扩增; 位点 HJ126、HJ4203以及 HJ4249对许氏平 鲉 、褐菖 鲉显示多态, 且对褐菖 鲉多态性更高; 位点 HJ4137、HJ4930、HJ4944以及 HJ9578在三个物种中均具有较高多态性, 且在许氏平 鲉中多态性最高(表3)。位点HJ4136 在朝鲜平 鲉 、褐菖 鲉和许氏平 鲉中的多态性检测结果见图2。 到目前为止, NCBI 公共数据库中许氏平 鲉EST序列共 1980条, 相比研究较早的水生生物来说数目较少。本研究中许氏平 鲉EST-SSR位点发现率为9.14%, 而海带为 5.03%(王国良, 2010)、坛紫菜为5.64%(杨惠等, 2009)、泥蚶为 6.50%(董迎辉等, 2013)、中华鳖为 7.45%(许晓军等, 2013)、牙鲆为 7.95%(陈松波等, 2010), 这说明相较于其它水生生物, 许氏平鲉EST-SSR位点发现率较高, 具有较好的开发潜力。本研究查找到的许氏平 鲉EST-SSR核心序列以二碱基重复出现频率最高, 这一结果与有关马氏珠母贝(邱樱, 2013)、牙鲆(陈松波等, 2010)以及中华鳖(许晓军等, 2013)的研究结果相同, 而在其它学者的研究中也存在不同结果, 比如在泥蚶、海带和坛紫菜中均以三碱基重复最丰富(杨惠等, 2009; 王国良, 2010; 周小龙等, 2013)。 图2 位点HJ4136 在朝鲜平 鲉 和褐菖 鲉 以及许氏平 鲉中的检测结果Fig.2 The results of transferability of locus HJ4136 in Sebastes koreanus, Sebastiscus marmoratus and Sebastes schlegeliia: 位点HJ4136 在朝鲜平鲉 中的通用性检测结果; b: 位点HJ4136在褐菖鲉中的通用性检测结果; c: 位点HJ4136在许氏平鲉中多态性检测结果 表3 18个位点通用性检测结果Tab.3 The results of transferability of 18 loci 许氏平 鲉EST-SSR核心序列以TG/CA基序最为常见, 在 205个完美型位点中占到 17.56%, 李霞等(2004)有关剑尾鱼的研究结果与之类似。18个多态EST-SSR位点中, 有4个位点核心序列是TG或CA基序, 重复次数范围在5—14之间, 占完美型多态位点比例 26.67%, 因此在本研究中许氏平 鮋EST-SSR位点TG/CA基序的开发效率较高; 上述 4个多态位点分别是 HJ126、HJ4136、HJ4959以及 HJ4137, 对应的核心序列重复次数和等位基因个数分别为5、5、12、14和 2、4、7、9, 由此可见随着核心序列重复次数增加位点多态性有逐渐增强的趋势。通常认为微卫星多态性的形成与复制过程中的滑链错配有关(张云武等, 2001), 由此形成的插入或缺失突变会导致微卫星核心序列长度的变化(乔洪金等, 2012), 而核心序列重复次数越高其出现插入或缺失突变的几率就越大。 许氏平 鲉46对有效扩增的EST-SSR引物中, 其中 18对具有多态性, 多态检测率为 39.13%; An等(2009)开 发的许氏平 鲉Genomic-SSR位点14对有效扩增, 其中 13对多态, 多态检测率高达 92.86%;Yasuike等(2013)设计30对完美型Genomic-SSR引物,其中 17对有效扩增并且显示多态, 多态检测率为100.00%。EST序列中有效扩增的微卫星引物中, 具有多态性的微卫星比例明显低于基因组中的微卫星比例。此外, Bai等(2011)开发的多态Genomic-SSR位点平均观测等位基因数、平均观测杂合度以及平均期望杂合度分别为 5.7、0.4194和 0.5002, 均高于本研究开发的EST-SSR, 同样的结果也存在于An等(2009)和Yasuike等(2013)开发的许氏平鲉Genomic-SSR标记中。上述结果均证实了以往学者有关 EST-SSR位点多态性低于 Genomic-SSR位点的结论(齐晓艳等,2013; 周小龙等, 2013)。一般认为这是由于EST序列来源于基因编码区, 更易受到选择压力的作用从而表现出较高的保守性, 因此多态性要比基因组 SSR低(Chabaneet al, 2005)。 微卫星侧翼序列在近缘物种中比较保守, 根据已有物种的微卫星引物实现跨物种通用是开发微卫星引物的一种有效途径。分子标记可以被分为两类,Ⅰ型分子标记与已知功能的基因相关联, 而Ⅱ型分子标记与基因组未知区域相关(O’Brien, 1991)。EST序列来源于功能基因, 由此开发的 EST-SSR引物属于Ⅰ型微卫星引物, 而Ⅰ型微卫星引物已被证实比未知基因背景的Ⅱ型引物通用性更强(Holtonet al,2002)。Ma等(2010, 2011)开发了拟穴青蟹Ⅱ型和Ⅰ型微卫星引物, 并分别检验其在同属物种中的通用情况, 结果显示紫螯青蟹(Scylla tranquebarica)对拟穴青蟹(Scylla paramamosain)Ⅰ型微卫星引物的通用性更强。本研究中开发的18对多态EST-SSR引物对褐菖 鲉的通用率和多态率分别达到100%和88.89%, 显著优于An等(2009)开发的14对Genomic-SSR位点,后者的相关数据分别为78.57%、78.57%。 一般认为, 对于微卫星引物, 亲缘关系越近的物种实现扩种扩增的可能性越大, 而在本研究中, 与许氏平鲉亲缘关系较远的褐菖鲉 (Sebastiscus marmoratus)却表现出了更好的通用性。此外, 通过分析 An等(2009)有关许氏平 鲉14对微卫星引物在4个近缘种中的通用性实验结果得知, 与许氏平 鲉亲缘关系较近的Sebastes inermis通用情况(通用率 10/14, 多态率10/14)却没有Sebastes marmoratus(通用率 11/14, 多态率11/14)好 。同样的结果都与褐菖 鲉(Sebastiscus marmoratus)这个物种有关, 具体原因有待进一步研究分析。另外, 上述结果与赵丽丽(2008)在青石斑鱼和美洲黑石斑鱼的跨种扩增结果类似, 后者认为这可能与跨种扩增研究中使用的样品数量有限、DNA复制时错配而产生假阳性带、副产物带过多、多态位点判断等原因有关。此外, 通用性实验结果显示, 位点HJ606和HJ4183 在许氏平 鲉 和褐菖 鲉中可以有效扩增而在朝鲜平 鲉中不能有效扩增, 这一结果得到进一步确认后可以应用于从上述三个物种中鉴定朝鲜平 鲉的工作。 本研究结果显示, 许氏平 鲉EST-SSR位点检出率较高, 具有较好的开发潜力, 实验开发的多态EST-SSR 位点可用于许氏平 鲉群体遗传多样性分析、系统进化分析和近缘种通用性检测等研究, 为今后许氏平 鲉的良种选育工作奠定基础。 王国良, 2010. 海带EST-SSR标记开发及TPS基因的克隆和比较遗传学研究. 青岛: 中国海洋大学硕士学位论文, 31—34 乔洪金, 刘相全, 孙国华等, 2012. 大竹蛏(Solen grandis)cDNA文库中微卫星标记的筛选. 海洋与湖沼,43(6): 1128—1133 齐晓艳, 董迎辉, 姚韩韩等, 2013. 文蛤30个微卫星标记的开发及在斧文蛤和帘文蛤中的通用性检测. 水产学报, 37(8):1147—1154 许晓军, 张海琪, 张 超等, 2013. 中华鳖表达序列标签资源中的微卫星信息分析. 经济动物学报, 17(1): 15—18 李 霞, 白俊杰, 吴淑勤等, 2004. 剑尾鱼微卫星DNA的筛选.中国水产科学, 11(3): 196—201 杨 惠, 茅云翔, 孔凡娜等, 2009. 坛紫菜EST-SSR筛选及其在遗传多样性分析中的实用性. 中国海洋大学学报, 39(2):265—270 邱 樱, 2013. 马氏珠母贝EST-SSR和SNP标记开发及其与珍珠层性状的关联分析. 海口: 海南大学硕士学位论文, 3—4 张云武, 张亚平, Ryder O A, 2001. 微卫星及其应用. 动物学研究, 22(4): 315—320 陈松波, 龚 丽, 刘海金, 2010. 牙鲆EST资源的SSR信息分析. 东北农业大学学报, 41(10): 82—86 易少奎, 高泽霞, 罗 伟等, 2013. 团头鲂 EST-SSR在厚颌鲂中的跨种扩增及杂交F1的鉴定. 水产学报, 37(7): 970—977 周小龙, 朱靖华, 董迎辉等, 2013. 泥蚶(Tegillarca granosa)基因组 SSR和 EST-SSR的开发及比较研究. 海洋与湖沼,44(2): 467—475 赵丽丽, 2008. 3种石斑鱼微卫星标记的开发及跨种扩增. 南昌:南昌大学硕士学位论文, 32—34 侯战辉, 王 嫣, 石耀华等, 2008. 马氏珠母贝(Pinctada martensii)2个不同地理种群遗传变异的EST-SSR分析. 海洋与湖沼, 39(2): 178—183 董迎辉, 吴国兴, 姚韩韩等, 2013. 泥蚶34个EST-SSR标记的开发及在格粗饰蚶中的通用性检测. 水产学报, 37(1): 70—77 An H S, Park J Y, Kim M-Jet al, 2009. Isolation and characterization of microsatellite markers for the heavily exploited rockfishSebastes schlegeli, and cross-species amplification in four relatedSebastesspp.. Conservation Genetics, 10(6): 1969—1972 Bai C C, Liu S F, Zhuang Z Met al, 2011. Isolation and characterization of microsatellite markers for the Korean rockfish,Sebastes schlegeli. Genetics and Molecular Research, 10(3): 2065—2068 Chabane K, Ablett G A, Cordeiro G Met al, 2005. EST versus genomic derived microsatellite markers for genotyping wild and cultivated barley. Genetic Resources and Crop Evolution,52(7): 903—909 Chistiakov D A, Hellemans B, Volckaert F A, 2006.Microsatellites and their genomic distribution, evolution,function and applications: A review with special reference to fish genetics. Aquaculture, 255(1—4): 1—29 Holton T A, Christopher J T, McClure Let al, 2002.Identification and mapping of polymorphic SSR markers from expressed gene sequences of barley and wheat.Molecular Breeding, 9(2): 63—71 Ma H Y, Ma C Y, Ma L Bet al, 2010. Novel polymorphic microsatellite markers inScylla paramamosainand cross-species amplification in related crab species. Journal of Crustacean Biology, 30(3): 441—444 Ma H Y, Ma C Y, Ma L B, 2011. Identification of type I microsatellite markers associated with genes and ESTs inScylla paramamosain. Biochemical Systematics and Ecology,39(4—6): 371—376 Marshall T C, Slate J, Kruuk L E Bet al, 1998. Statistical confidence for likelihood-based paternity inference in natural populations. Molecular Ecology, 7(5): 639—655 O’Brien S J, 1991. Mammalian genome mapping: lessons and prospects. Current Opinion in Genetics & Development,1(1): 105—111 Rousset F, 2008. GENEPOP’007: a complete re-implementation of the GENEPOP software for Windows and Linux.Molecular Ecology Resources, 8(1): 103—106 Schug M D, Wetterstrand K A, Gaudette M Set al, 1998. The distribution and frequency of microsatellite loci inDrosophila melanogaster. Molecular Ecology, 7(1): 57—70 Tautz D, Renz M, 1984. Simple sequences are ubiquitous repetitive components of eukaryotic genomes. Nucleic Acids Research, 12(10): 4127—4138 Weber J L, 1990. Informativeness of human (dC-dA)n·(dG-dT)npolymorphisms. Genomics, 7(4): 524—530 Yasuike M, Noda T, Fujinami Yet al, 2013. Tri-, tetra- and pentanucleotide-repeat microsatellite markers for the Schlegel’s black rockfishSebastes schlegelii: the potential for reconstructing parentages. Conservation Genetics Resources, 5(2): 577—581 Yeh F C, Yang R, Boyle T, 1999. POPGENE version 1.31:Microsoft Window Based Freeware for Genetic Analysis.Alberta, Canada: University of Alberta and Centre for International Forestry Research Yoshida K, Nakagawa M, Wada S, 2005. Multiplex PCR system applied for analysing microsatellite loci of Schlegel’s black rock fi sh,Sebastes schlegeli. Molecular Ecology Notes, 5(2): 416—418

2.4 许氏平鲉多态性微卫星引物的通用性检测
3 讨论