京海黄鸡新城疫和传染性支气管炎抗病性状的全基因组关联分析
2015-03-22王文浩王金玉张跟喜樊庆灿陈学森韩鲲鹏王永娟
王文浩,张 涛,王金玉*,张跟喜,樊庆灿,陈学森,韩鲲鹏,王永娟
(1.扬州大学动物科学与技术学院,扬州 225009; 2.江苏省动物遗传繁育与分子设计重点实验室,扬州 225009;3.江苏京海禽业集团,南通 226103)
京海黄鸡新城疫和传染性支气管炎抗病性状的全基因组关联分析
王文浩1,2,张 涛1,2,王金玉1,2*,张跟喜1,2,樊庆灿1,2,陈学森1,2,韩鲲鹏1,2,王永娟3
(1.扬州大学动物科学与技术学院,扬州 225009; 2.江苏省动物遗传繁育与分子设计重点实验室,扬州 225009;3.江苏京海禽业集团,南通 226103)
旨在寻找影响京海黄鸡新城疫和传染性支气管炎抗病性状的SNP位点。本研究采用简化基因组测序的方法,检测京海黄鸡基因组的SNP位点,并对这些位点与新城疫和传染性支气管炎性状进行关联分析。结果表明:在基因组水平上有1个与京海黄鸡新城疫抗病性状显著相关的SNP位点,7个与该性状潜在显著的SNPs位点,并将这些位点定位到5个基因上,分别为EEA1、CARS2、SCML2、GRP20以及TOMIL2基因,这些基因均可能影响或调控机体的抗病及免疫能力,可以考虑作为京海黄鸡新城疫抗病性状的候选基因作为后续研究。没有发现与京海黄鸡传染性支气管炎显著相关的SNP位点,该性状可能是复杂性状,受到多种因素的影响。因此,本研究发现的几个标记位点可能对京海黄鸡的新城疫抗病性状存在一定的影响,可以作为候选基因,为京海黄鸡抗病育种过程中的标记辅助选择提供参考资料。
京海黄鸡;简化基因组测序;抗病性状;SNPs
全基因组关联分析(GWAS)是以遍布于整个基因组的单核苷酸多态性(SNP)为分子标记,以发现影响复杂性状发生的遗传标记和遗传标记的分布特征为目的,对复杂的经济性状进行直接的关联分析[1]。与以往的方法相比,全基因组关联分析最大优点:不需要在研究之前,根据那些并没有完全研究清楚的分子机理来假定某个基因或位点与研究的经济性状相关联。即GWAS不需要构建任何不确定的假设。GWAS是直接在全基因组范围内筛选与重要经济性状相关联的方法[2]。
2005年,《Science》发表了一篇关于年龄相关性视网膜黄斑变性的GWAS文章,引起了生物学界和医学界学者们的广泛关注[3],这是GWAS的首次报道。随后,研究者们开展了一系列关于人类遗传疾病的GWAS研究。2008年,J.C.Barrett[4]使用全基因关联分析发现了多个与克罗恩病关联显著的SNP位点。2009年,L.A.Weiss[5]对人的自闭症进行的全基因组关联分析,发现了相关的染色体区域。在猪上,N.Duijvesteijn 等[6]以 987 头杜洛克为试验动物,使用60K SNP芯片,研究了公猪屠体中的雄烯酮含量,发现了37 个与雄烯酮含量关联显著的SNP,这些SNP主要分布于1号和6号染色体上。在牛上,L.Jiang等[7]以14个半同胞家系组成的2 093中国荷斯坦奶牛为试验动物,使用牛50 kb SNP 芯片,结合混合线性模型和遗传传递不平衡的方法,对产奶性状进行了全基因组关联分析,发现了105个与中国荷斯坦奶牛产奶性状关联显著的SNPs。家禽上,R.Liu等[8]使用鸡Illumina 60K SNP芯片,研究北京油鸡的屠宰和肉质性状,发现了24个与鸡的屠宰性状关联显著和潜在显著的SNP。L.Xie 等[9]使用鸡Illumina60 K SNP芯片,对白洛克和杏花鸡的F2代杂交群体的体重性状和日增重性状进行了全基因组关联分析,发现鸡1号染色体的173.5~175.0区域是与鸡的体重和日增重性状关联显著的SNP集中区域。该区域存在极为明显的连锁现象。
每年因为疾病给家禽业带来巨大的损失。对于抗病性能的选择,常规育种具有一定的难度,分子遗传学技术虽然迅猛发展,但在鸡抗病研究未见有实质性的成效。相对于生产性能,家禽抗病性的研究总体偏少[10],对于抗病育种的选择方法、选择性状和选择标记都仍然处于探索中。本研究采用简化基因组测序技术,旨在寻找影响京海黄鸡新城疫和传染性支气管炎抗体水平相关联的SNPs和相关的基因。
1 材料与方法
1.1 试验动物
试验群体为京海黄鸡育种场11世代核心群纯种母鸡400只。所有母鸡同一批次孵化,在同一鸡舍内,以相同条件饲养,其健康状况良好,无患病记录,且均有系谱可查,彼此间亲缘关系清楚。所有个体在1日龄时带翅号,按照既定免疫日程进行免疫(表1)。所有个体于60日龄时采集血液3 mL,其中1.5 mL用于禽血DNA的提取,1.5 mL用于析出血清,测定新城疫和传染性支气管炎抗体滴度。其中,DNA提取采用北京艾德莱生物公司试剂盒,用核酸分析仪分析其浓度和纯度。抗体滴度的测定采用美国AFFINITECH新城疫和传染性支气管炎抗体ELISA检测试剂盒。
1.2 简化基因组测序
提取后质检合格的DNA样品送于北京百迈克生物技术公司进行简化基因组测序。测序原理:利用酶切预测软件对参考基因组的GC含量、重复序列情况和基因特点等信息进行分析,设计标记开发方案。利用上述分析得出的酶切组合对DNA进行酶切打断试验,将酶切产物进行一系列修复、修饰后进行桥式扩增。用琼脂糖凝胶电泳进行片段大小选择,通过PCR扩增,增大文库量,建好的文库用Illumina HiSeqTM2000进行测序(图1)。然后将测序得到的reads(双端序列)比对到参考基因组Gallus_gallus.WASHUC2.59上,通过筛选获得SLAF片段后进行SNP检测。
1.3 统计分析
通过SPSS11.0软件统计所测抗体水平的均值、极值、方差、标准差、偏度等。考虑到群体遗传背景间差异可能导致的群体分层现象,通过主成分分析法对群体分层进行评估,再绘制QQ-plot图来对群体分层进行检测。全基因组关联分析采用GAPIT软件的混合线性模型(Mixed liner model,MLM)完成,公式:
表1 京海黄鸡免疫日程
Table 1 Jinghai Yellow chicken’s immunization schedule
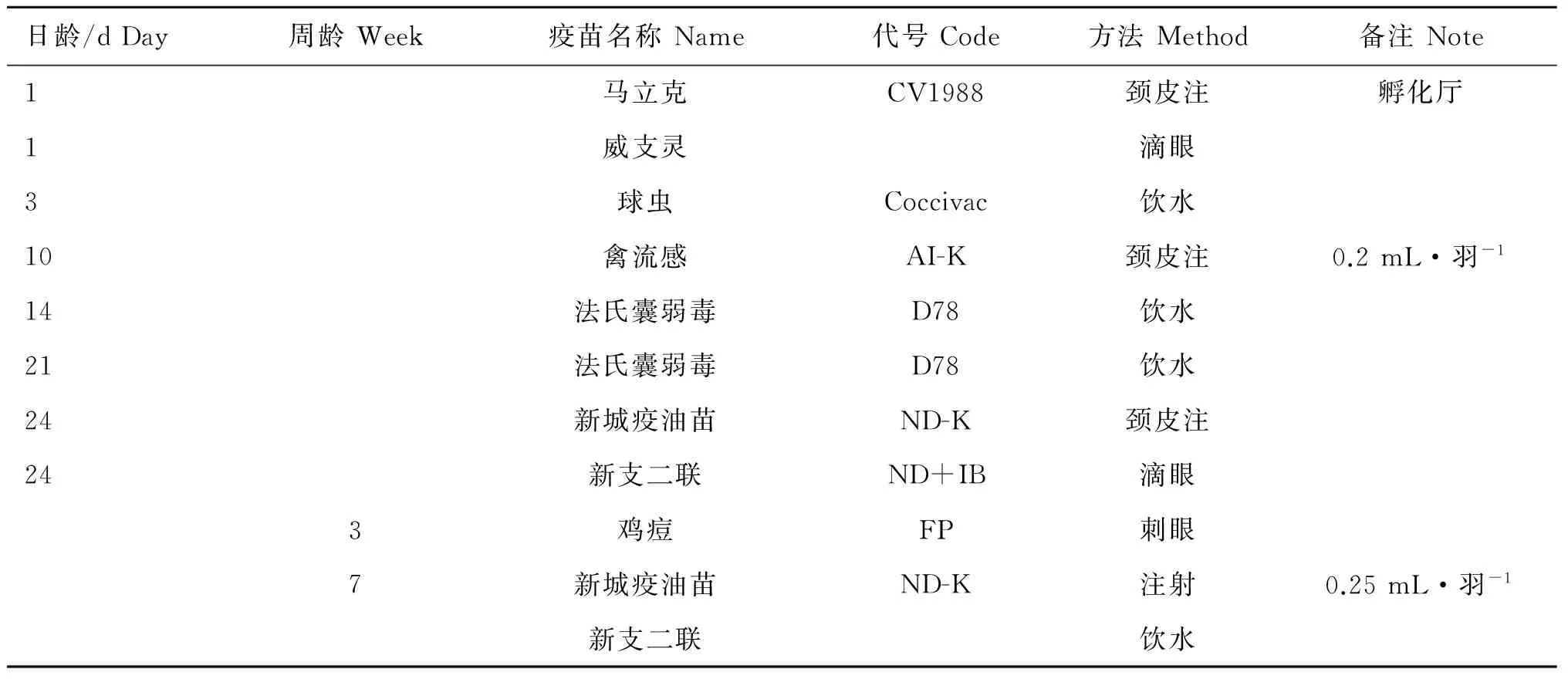
日龄/dDay周龄Week疫苗名称Name代号Code方法Method备注Note1马立克CV1988颈皮注孵化厅1威支灵滴眼3球虫Coccivac饮水10禽流感AI-K颈皮注0.2mL·羽-114法氏囊弱毒D78饮水21法氏囊弱毒D78饮水24新城疫油苗ND-K颈皮注24新支二联ND+IB滴眼3鸡痘FP刺眼7新城疫油苗ND-K注射0.25mL·羽-1新支二联饮水
y=Xα+Qβ+Kμ+e
其中,通过PCA计算样品的聚类主成分,以PC1、PC2定义群体的群体结构Q,通过GAPIT软件计算样品间亲缘关系Kinship(K),X为基因型,y为表型,最终每个SNP位点都能得到一个与相关性状的关联值;用Bonferroni法对混合线性模型计算得出的P值进行多重假设检验校正[10],筛选出全基因组范围内显著、潜在显著的SNP位点。
1.4 功能位点基因注释和生物学功能挖掘
使用BLAST软件将关联位点前后50 kb范围内的基因序列与参考基因组Gallus_gallus.WASHUC2.59进行对照,找出该位点在基因组上所在位置。并将此结果与 nr、SwissProt、GO、COG、KEGG等数据库进行比对,获得多变区基因的注释信息,进而探索基因与性状之间的关系。
2 结 果
2.1 DNA质检及抗体水平测定
提取的DNA使用核酸蛋白分析仪测定基因组DNA的浓度和纯度,确保所有基因组浓度大于100 ng·mL-1,OD260 nm/OD280 nm值为1.7~1.8。使用 1%的琼脂糖凝胶电泳检测基因组DNA有无拖尾现象(图2)。新城疫和传染性支气管炎抗体滴度的描述性统计见表2。

图1 简化基因组测序原理Fig.1 The theory of SLAF-seq

图2 凝胶电泳检测图Fig.2 Gel electrophoresis of each sample
表2 凝抗体滴度描述性统计分析
Table 2 Antibody’s descriptive statistical analysis
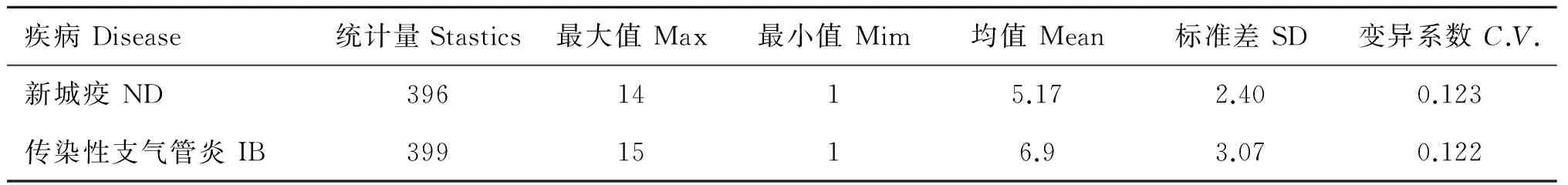
疾病Disease统计量Stastics最大值Max最小值Mim均值Mean标准差SD变异系数C.V.新城疫ND3961415.172.400.123传染性支气管炎IB3991516.93.070.122
2.2 简化基因组测序
根据简化基因组测序技术,首先进行酶切,然后对酶切片段双端测序后,所有样品共得到329 314 696个测序reads,保留测序单碱基错误率占总碱基数目的比率在1/100以下的resds,再使用SOAP软件将各个样品数据比对到参考基因组Gallus_gallus.WASHUC2.59,选取双端reads都比对到基因组上的reads,Pair-end和Single-end效率越高,测序质量越好。得到整体的比对效率统计结果见表3。再根据比对纠错结果,选取样品平均深度在2以上的group,定义SLAF标记,得到94 926个SLAF标记以及在每个染色体上的分布(表4),再根据定义好的SLAF标记,进行基于样品的群体内部SNP检测(表5),再对测得的SNPs做质量控制:剔除基因型检出率小于80%、剔除最小基因频率小于3%的个体,最终得到符合条件的可用于后续分析的306 322个SNPs。
表3 获得的reads与参考基因组对比后结果

Table 3 The comparison result between obtained reads number and reference genome sequence %
Pair-end percentage为双端都比对到基因组上的reads占总reads的比例;Single-end percentage为一端比对到基因组上而另一端没比对到基因组上的reads占总reads的比例;Unmap percentage为未比对到基因组上的reads占总reads的比例
Pair-end percentage means the ratio of double end sequence to the original sequence;Single-end percentage means only one end can comparise to the original sequence;Unmap percentage means no end can comparise to the original sequence
表4 SLAF标记统结果
Table 4 SLAF marker’s statistical result
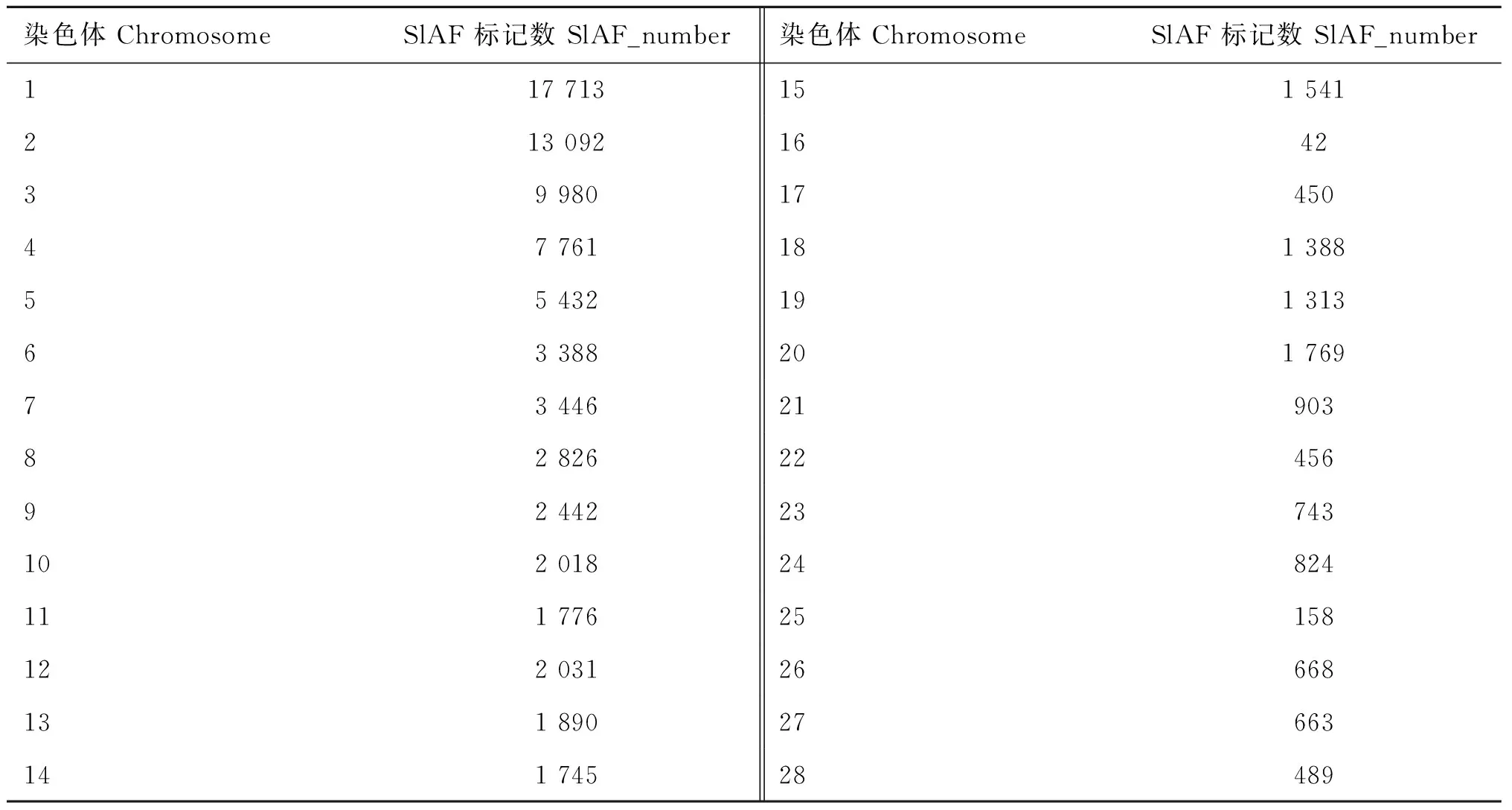
染色体ChromosomeSlAF标记数SlAF_number染色体ChromosomeSlAF标记数SlAF_number117713151541213092164239980174504776118138855432191313633882017697344621903828262245692442237431020182482411177625158122031266681318902766314174528489
表5 SLAF多态性统计结果
Table5 SLAF’s polymorphism statistical result

SLAF数量SLAFnumber总深度Totaldepth平均深度AveragedepthSLAFpolyPoly率/%Polyratio949261586974055.189492099.99
2.3 统计分析
基于SNP,通过admisture软件,计算样品的群体结构,做样品的群体结构聚类分析,分别假设400个样品的分群数(K值)为1~10,进行聚类,聚类结果如图3;再充分考虑个体间亲缘关系可能导致的群体分层的影响,做出2个性状的QQ-plot图(图4),如图4所示,通过SNP关联分析计算的观测值(纵坐标)在期望值(横坐标)上方,说明并无群体分层现象,基于混合线性模型的关联分析结果是比较可靠的,可以进行后续分析;将聚类分析结果带入GAPIT混合线性模型分析软件,将简化基因组测序得出的SNPs分别与京海黄鸡的新城疫、传染性支气管炎抗体滴度指标进行关联分析,每个SNP都对应一个与相关性状进行关联分析后得到的P值,再用Bonferroni法对该混合线性模型计算得出的P值进行多重假设检验校正,得出在全基因组水平上显著关联的P值为3.3×10-7,在全基因组水平上潜在显著关联的P值为3.3×10-6。
根据上述分析模型以及多重假设检验校正,在全基因组水平上找到了1个与新城疫抗病性状显著相关的SNP位点,找到了7个与新城疫抗病性状潜在显著的SNPs位点,每个位点序列号、所在染色体上位置、对应P值以及距离最近基因如表6所示;而对于传染性支气管炎抗病性状,未找到在全基因组水平上显著或者潜在显著的SNP位点。

A图中每种颜色代表一个群,每行代表一个分群值的情况;图中展示了400个样品分群值为1~10的聚类情况,B图中为每个K值对应的C.V.值,K为1的时候最小In the figure A,every colour stands for a group,each line means a situation of a cluster;Figure B shows clustering situation that the cluster number varies from 1 to 10图3 群体结构聚类图Fig.3 Clustering figure of group structure

上图通过SNP关联分析计算的观测值(纵坐标)均在期望值(横坐标)上方,说明并无群体分层现象Both of the observed values are higher than expected values,shows no population stratification图4 两个性状的QQ-plot图Fig.4 QQ-plot pictures of two traits
2.4 功能位点的基因注释和生物学功能挖掘
基于鸡基因组Gallus_gallus.WASHUC2.59的基因组信息对GIWAS分析结果进行了注释,并一一列出(表6),同时用nr、SwissProt、GO、COG、KEGG等数据库对上述5个基因进行了注释,探索其生物学功能以及与性状之间可能的关系。
早内体抗原1(Early endosome antigen 1,EEA1)。上述数据库注释后得出该蛋白细胞分布为膜片段、核、早内体、胞浆、外部质膜;该蛋白能具有保守的结构域,如核酸结合域、钙调蛋白结合域、锌离子结合域等,参与膜泡结合、突触膜泡内体融合、早内体晚内体运输等一系列生命活动。
半胱氨酸-转移核糖核酸酶2(Cysteinyl-tRNA synthetase 2,CARS2),根据上述数据库的注释,得出该蛋白细胞分布为线粒体、胞浆;分子功能包括调控半胱氨酸-转移核糖核酸连接酶活性,也具有保守的结构域,如核苷酸结合域、ATP结合域以及金属离子结合域;参与的生物学过程包括半胱氨酸-转移核糖核酸的氨酰化作用。
中足性梳样2(Sex comb on midleg-like 2,SCML2),根据上述数据库的注释,得出其细胞分布为PcG蛋白质复合体、核腔、染色质核仁;分子功能包括调控序列特异性DNA结合转录因子活性、也具有核小体结合域、组蛋白结合域、DNA结合域;参与的生物进程包括染色质重组、DNA模板化转录的正负调控、精子的形成、胚胎的形态发生、造血作用、细胞分化、细胞周期的调控、多细胞动物有机体发育的调控等。
G蛋白偶联受体20(G protein-coupled receptor 20,GPR20),属于与G蛋白有信号连接的一大类受体家族的一员,根据上述数据库的注释,得出其为质膜的有机组成部分;分子功能包括调控G蛋白偶联受体的活性、具有特定的蛋白结合域、小分子结合域;参与的生物进程包括参与免疫系统内淋巴细胞的活化、白血球的分化、正调控白血球的活化作用和免疫效应的过程、调控骨髓白细胞介导的免疫、参与肾的系统进程包括全身动脉血压的调控、G蛋白偶联受体的信号通路、血液凝固、负调控信号转导、正调控细胞的代谢过程、粒细胞的激活、正调控细胞因子的分泌等。
myb1样靶标2(Target of myb1-like 2,TOM1L2),根据上述数据库的注释,得出其存在于胞质、胞膜、早期内体、部分细胞器;分子功能包括调控蛋白激酶活化剂的活性、有特定的蛋白质结合域;参与的生物进程包括细胞内蛋白质的转运、内吞作用、跨膜受体蛋白酪氨酸激酶信号通路、内小体转运。
表6 每个SNP位点在染色体上位置以及对应P值
Table 6 The chromosome location of every SNP and itsPvaule
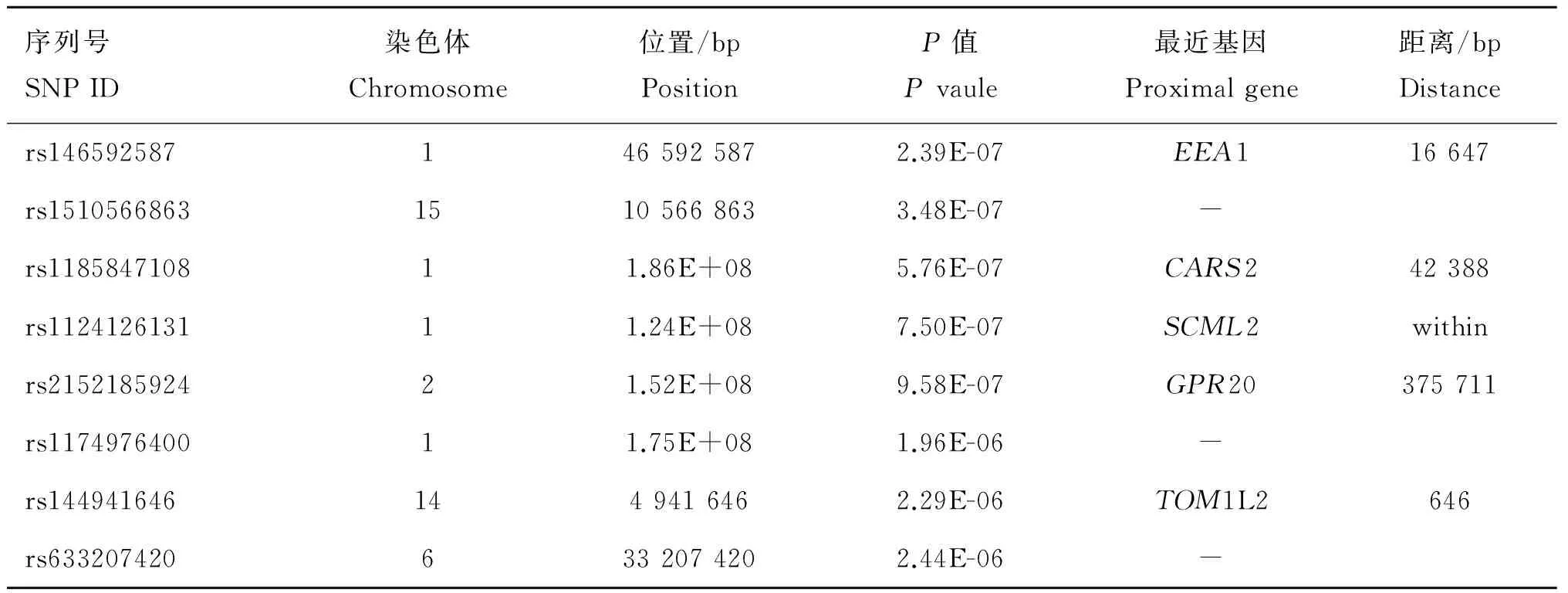
序列号SNPID染色体Chromosome位置/bpPositionP值Pvaule最近基因Proximalgene距离/bpDistancers1465925871465925872.39E-07EEA116647rs151056686315105668633.48E-07-rs118584710811.86E+085.76E-07CARS242388rs112412613111.24E+087.50E-07SCML2withinrs215218592421.52E+089.58E-07GPR20375711rs117497640011.75E+081.96E-06-rs1449416461449416462.29E-06TOM1L2646rs6332074206332074202.44E-06-
3 讨 论
本研究采用了简化基因组测序的方法来寻找全基因组范围内的SNP,简化基因组测序(Reduced-representation sequencing)技术即是指利用生物信息学方法,设计标记开发方案,筛选特异性长度片段,应用高通量测序技术获得海量标签序列来充分代表目标物种全基因组信息的测序方法,是将整个基因组进行“简化”后进行的测序。它有着通量高、准确率高、数据利用率高、性价比高以及不受参考基因组限制,对没有参考基因组的物种也可以进行大规模筛查SNP位点的优点[11-12],与传统的芯片技术相比,它更有助于一些未知SNP位点的发现。后续的多重假设检验校正时,公式为P=ɑ/n,其中n为质控后所有用于后续分析的SNP数,鸡上较常用的芯片一般找出的SNPs数约几万个,而本试验经质控合格后用于后续分析的SNPs超过30万个,因此为了降低假阴性结果的出现,适当提高了ɑ值(由0.05提高到0.1),最终得出在全基因水平上显著关联的P值。
目前在鸡上对于遗传相关性的研究主要集中在生长、繁殖等性状,对于抗病性状的研究并不多,究其原因是现在的抗病性状界限难以区分,还存在一个确定的抗病性状到底能不能代表抗病能力的问题,以及抗病性状较难检测、检测费用较高等,熟知的抗病性状大多数是抗体[10]。本研究中,所有京海黄鸡均确保同批次孵化、健康状况良好、饲喂及外周环境保持一致、同时段免疫、免疫后同一日程采血,取血清检测其抗体滴度,力求将可能影响结果的负面影响减到最小。
本研究发现,1号染色体上的EEA1基因与京海黄鸡的新城疫抗体水平在基因组水平上显著相关。最早从人体内克隆出了一段编码一种进化上保守的180 ku的蛋白的cDNA,并命名为EEA1[11],在人上,EEA1是一种与早期核内体胞质面瞬发性相关联的亲水性外周膜蛋白,同时EEAI自身抗体被认为与神经系统疾病、亚急性皮肤红斑狼疮以及韦格纳肉芽肿病有关[13],同时根据数据库注释结果表明,该基因还具有特异性的核苷酸结合域,而根据已有文献植物抗病基因编码的蛋白大多富含氨基酸重复单位和核苷酸结合位点[14-15],在动物上未见该基因的相关报道,该基因可作为影响京海黄鸡新城疫抗体水平的候选基因进行后续研究。同时鉴定出4个基因(CARS2、SCML2、GPR20、TOMIL2)与京海黄鸡新城疫抗体滴度存在潜在关联。其中GPR20是G蛋白偶联受体家族的一员,与核苷酸和脂类受体密切相关,并且已经证明GPR20在没有配合基的情况下能够保持持续的活性来确保与它偶联的G蛋白持续的活性[16];CARS2是一种半胱氨酸-转移核糖核酸酶,目前已经在染色体上靠近CARS基因的地方发现了与糖尿病性肾病显著相关的位点,并证明了这个I型糖尿病重要的晚期并发症之前发病机理的通路研究[17];SCML2属于多梳家族的一员,该家族被认为参与控制哺乳动物的细胞增殖以及肿瘤发生[18];TOMIL2基因目前未见相关报道。根据文献和基因注释结果综合来看最终得到的5个基因均可能影响机体的抗病及免疫能力,可以考虑作为候选基因进行后续研究。本研究未发现与京海黄鸡传染性支气管炎抗病性状显著相关的SNP位点,可能是由于该性状是一个复杂性状,受到多重因素的影响相互抵消的结果。
4 结 论
本研究采用简化基因组测序的方法在全基因组范围内寻找影响京海黄鸡新城疫和传染性支气管炎抗病性状的SNP位点,最终在全基因范围内找到了1个与京海黄鸡新城疫抗病性状显著相关的SNP位点,找到了7个与京海黄鸡新城疫抗病性状潜在显著相关的SNPs位点,最终共定位到5个基因上,这5个基因均可能影响机体的抗病及免疫能力;全基因组范围内未发现与传染性支气管炎显著相关的SNP位点。该结果能够为京海黄鸡及其他品种的研究提供一定的参考。
[1] HARDY J,SINGLETON A.Genomewide association studies and human disease [J].NewEnglandJMed,2009,360(17):1759-1768.
[2] LIU Y Z,PEI Y F,LIU J F,et al.Powerful bivariate genome-wide association analyses suggest the SOX6 gene influencing both obesity and osteoporosis phenotypes in males [J].PLoSONE,2009,4(8):e6827.
[3] KLEIN R J,ZEISS C,CHEW E Y,et al.Complement factor H polymorphism in age-related macular degeneration [J].Science,2005,308(5720):385-389.
[4] BARRETT J C,HANSOUL S,NICOLAE D L,et al.Genome-wide association defines more than 30 distinct susceptibility loci for Crohn’s disease [J].NatGenet,2008,40(8):955-962.
[5] WEISS L A.Autism genetics:emerging data from genome-wide copy-number and single nucleotide polymorphism scans [J].ExpertRevMolDiagnost,2009,9(8):795-803.
[6] DUIJVESTEIJN N,KNOL E F,MERKS J W,et al.A genome-wide association study on androstenone levels in pigs reveals a cluster of candidate genes on chromosome 6 [J].BMCGenet,2010,11(1):42.
[7] JIANG L,LIU J,SUN D,et al.Genome wide association studies for milk production traits in Chinese Holstein population [J].PLoSONE,2010,5(10):e13661.
[8] LIU R,SUN Y,ZHAO G,et al.Genome-wide association study identifies loci and candidate genes for body composition and meat quality traits in Beijing-You chickens [J].PLoSONE,2013,8(4):e61172.
[9] XIE L,LUO C,ZHANG C,et al.Genome-wide association study identified a narrow chromosome 1 region associated with chicken growth traits [J].PLoSONE,2012,7(2):e30910.
[10] 胡耀东.鸡 H/L 和 SRBC 遗传选择及部分抗病性状全基因组关联分析 [D].雅安:四川农业大学,2012. HU Y D.The analysis of genetic selection of disease-resistant trait (H/L and SRBC) in chicken and genome-wide association study for part of disease-resistant traits [D].Ya’an:Sichuan Agricultural University,2012.(in Chinese)
[11] MU F T,JUDY M,STEELE-MORTIMER C O,et al.EEA1,an early endosome-associated protein[J].JBiolChem,1995,6(2):503-511
[12] SUN X W,LIU D Y,ZHANG X F,et al.SLAF-seq:An efficient method of large-scale De-Novo SNP discovery and genotyping using high-throughput sequencing[J].PLoSONE,2013,3(8):e58700.
[13] SELAK S,WOODMAN R C,FRITZLER M J.Autoantibodies to early endosome antigen (EEA1) produce a staining pattern resembling cytoplasmic anti‐neutrophil cytoplasmic antibodies[J].ClinExpImmunol,2000,(122):493-498.
[14] 倪小勋,杨文才.番茄核苷酸位点(NBS)-亮氨酸富集重复(LRR)型抗病基因同源序列的分离与鉴定[J].农业生物技术学报,2013,(10):1145-1151. NI X X,YANG W C.Isolation and analysis of nucleotide binding site (NBS)- leucine- rich- repeat (LRR) type resistance gene analogs in tomato (SolanumlycopersciumL.) [J].JournalofAgriculturalBiotechnology,2013,(10):1145-1151.(in Chinese)
[15] 谢 华,陈绪清,朱 郎,等.白菜类富含亮氨酸重复(LRR)抗病蛋白基因BcLRR cDNA克隆及其介导的植物软腐病抗性分析[J].农业生物技术学报,2010,(3):416-423. XIE H,CHEN X Q,ZHU L,et al.cDNA cloning of an leucine-rich repeats (LRR)- like disease resistance protein gene BcLRR from Chinese cabbage and its functional analysis in mediating plant resistance to Soft- rot disease [J].JournalofAgriculturalBiotechnology,2010,(2):416-423.(in Chinese)
[16] MEMOKO H,TAKEHIKO Y,TAKAO S.Characterization of an orphan G protein-coupled receptor,GPR20,that constitutively activates Giproteins[J].JBiolChem,2008,(283):12747-12755.
[17] PEZZOLESI M G,POZNIK G D,MYCHALECKYJ J C,et al.Genomy-wide association scan for diabetic nephropathy susceptibility genes in type 1 diabetes[J].Diabetes,2009,(6):1403-1410.
[18] 吴海辉.灵长类scml1基因分子进化的初步研究 [D].北京:中国科学院研究生院,2009. WU H H.The preliminary research on the primate scml1 gene’s molecular evolution [D].Beijing:Graduate School of Chinese Academy of Sciences,2009.(in Chinese)
(编辑 郭云雁)
Genome-wide Association Studies for Jinghai Yellow Chicken’s ND and IB Disease Resistance Character
WANG Wen-hao1,2,ZHANG Tao1,2,WANG Jin-yu1,2*,ZHANG Gen-xi1,2, FAN Qing-can1,2,CHEN Xue-sen1,2,HAN Kun-peng1,2,WANG Yong-juan3
(1.CollegeofAnimalScienceandTechnology,YangzhouUniversity,Yangzhou225009,China; 2.KeyLaboratoryforAnimalGenetics,Breeding,ReproductionandMolecularDesignofJiangsuProvince,Yangzhou225009,China;3.JiangsuJinghaiPoultryIndustryGroupCo.,Ltd.,Nantong226103,China)
The objective of this study was to seek SNPs that influence Jinghai Yellow chicken’s ND and IB disease resistance character.The SNPs in Jinghai Yellow chicken were detected by SLAF-seq method and then the association of SNP sites with ND and IB disease resistance character were analyzed.The result showed that one SNP was significantly associated with ND disease resistance character,7 SNPs were potential significantly associated with ND disease resistance,and they were located in 5 genes:EEA1,CARS2,SCML2,GRP20 andTOMIL2;All of the 5 genes might influence or regulate organism’s disease resistance and immune ability and could be the candidate genes for Jinghai Yellow chicken’s disease resistance character.No SNP was found that was significantly associated with IB disease resistance character,this trait was complex and was rugulated by many factors.The results showed the 5 genes had influence on Jinghai Yellow chicken’s ND disease resistance characeter,and could provide the reference data for marker assisted selection of Jinghai Yellow chicken’s disease resistant breeding process.
Jinghai Yellow chiken;SLAF-seq;disease resistance character;SNPs
10.11843/j.issn.0366-6964.2015.02.004
2014-05-09
江苏省高校自然科学基金(12KJB230003);国家自然科学基金( 31201793);国家肉鸡产业技术体系(nycytx-42-G1-05);江苏高校优势学科建设工程;江苏省动物遗传繁育与分子设计重点实验室
王文浩(1988-),男,江苏射阳人,博士生,主要从事动物遗传育种与繁殖研究,E-mail:691042495@qq.com
*通信作者:王金玉,E-mail:jywang@yzu.edu.cn
S831.2
A
0366-6964(2015)02-0196-08
