乌拉盖草原不同群落类型α多样性研究
2015-03-18王君芳白耀华闫晓红
张 健,邢 旗,王君芳,白耀华,闫晓红
(内蒙古和信园蒙草抗旱绿化股份有限公司,呼和浩特 010030)
乌拉盖草原不同群落类型α多样性研究
张 健,邢 旗*,王君芳,白耀华,闫晓红
(内蒙古和信园蒙草抗旱绿化股份有限公司,呼和浩特 010030)
用比较样地法调查了乌拉盖草原9个不同群落类型的α多样性特征。结果表明:(1)物种数随取样面积增加呈对数增加,且与对数模型S=alnA+b能够很好拟合(R2≥0.9715)。(2)不同群落类型的α多样性测度值(丰富度指数、多样性指数、均匀度指数),都表现出山地草甸类“羊草+线叶菊+杂类草”类型、温性草甸草原类“羊草+杂类草”类型、山地草甸类“羊草+杂类草”类型具有较高的多样性指数,而低地草甸类“羊草群落”类型拥有较低多样性指数。(3)群落的多样性更多的受均匀度指数的影响。
群落类型;种-面积关系;α多样性
1 研究背景及意义
丰富多彩的生命世界是地球在浩瀚宇宙中最具魅力之处。各种生命形式息息相关,彼此作用,一起维系着生态系统的结构和功能。然而,由于人类活动的干扰不断加剧,大量生物正在以前所未有的速度灭绝。生物多样性与多种生态系统服务功能密切相关。生物多样性的丧失以及生态系统服务功能和可持续性的下降,将最终威胁到人类的福祉和生存。因此,生物多样性的保护已经刻不容缓。
生物多样性给我们人类带来了巨大的经济效益,如药用植物和工业上的重要原料。同时,生物多样性因其自身的多变性和复杂性,也给我们人类带来许多美学价值,是人类文化不可分割的一部分〔1〕。
生态系统中生物多样性的维持是全球生物多样性与生态系统管理的中心目标之一〔2〕。Yachi&Loreau〔3〕认为物种丰富度对生态系统生产力具有两方面的影响:(1)缓冲效应(buffering effect)即降低生产力随时间的波动;(2)性能增加效应(performance_enhancing effect)提高生产力的平均水平。除此之外,草地植物群落的生物多样性对草地生态系统的稳定性发展和生产力的维持具有很大作用〔4-6〕。
生物多样性包括物种多样性、遗传多样性和生态系统多样性。群落多样性是生态系统能量流动与物质循环的主要提供者,也是维持生态系统及调控全球变化的主要作用因素。一般认为α多样性就是物种多样性,物种多样性是指物种种类和数量的丰富程度〔7〕,是一个区域或一个生态系统可测定的生物学特征〔8〕,反映群落内部物种数和物种相对多的一个指标,只具有数量特征而无方向性。
内蒙古高原的草原植被是欧亚大陆草原区亚洲中部亚区的重要组成部分,而乌拉盖管理区位于锡林郭盟东北部,属草甸草原向典型草原过渡区,有“受人类活动干扰最少的锡盟天然草原”之美誉,研究其生物多样性对乌拉盖草地生态系统的稳定性发展和生产力的维持、保护乌拉盖草原植物的药用资源和生态系统功能都具有重要意义。
2 研究区概况与研究方法
2.1 研究区概况
试验示范区设在锡林郭勒盟乌拉盖管理区。乌拉盖管理区位于锡林郭盟东北部,大兴安岭山脉西麓中端。东经115°10′至120°7′,北纬44°40′至46°40′。气候属于半干旱半湿润大陆性气候,多年平均降水量322.3mm,年际变化较大,年内分布不均,降水量70%集中在6-8月份,且由东北向西南递减。无霜期90-110天。年平均蒸发量为1552.1mm,年平均气温-0.9℃,极端最低温度-42.1℃(发生于1987年1月9日),极端最高气温38.4℃(发生于1968年7月22日)。平均风速每秒4.1米。全年平均日照实数2659.8小时。海拔高度864米到1280米之间,平均海拔980米。土地总面积5013km2,其中草场占总面积的92.1%。
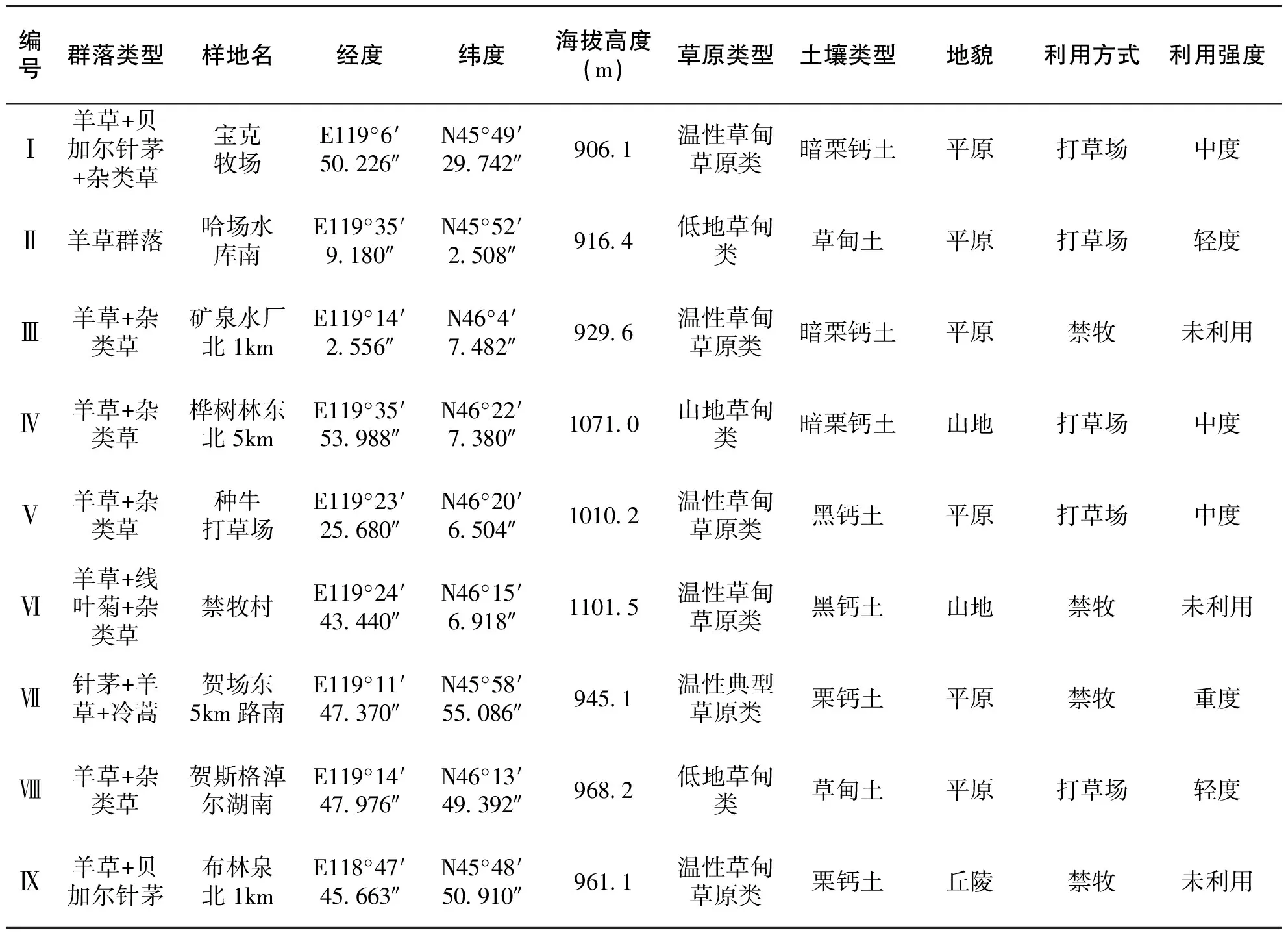
表1 观测样地基本情况
乌拉盖草原属草甸草原向典型草原过渡区,土壤条件好,主要以栗钙土、黑钙土为主。植物种类多样,是锡林郭勒盟地区产量较高、物种较丰富的草原,近年平均干草产量为115kg/亩,有501种野生植物,分属62科256属,种数最多的是菊科,其次是禾本科,再次为豆科等。

图1 试验样地在遥感影像分布图
本研究于2015年8月份选择了乌拉盖管理区不同群落类型的草原代表性群落典型地段,设置9个观测样地(样地基本情况见表1),各样地在遥感影像上的分布情况如图1。
2.2 观测取样方法
观测方法采用中国科学院植物研究所植被与环境变化重点实验室草原生态系统功能生态学研究组2012年对草地植物多样性野外调查的方法〔9〕。样线、样方、种-面积记录如图2所示。
(1)按事先规划好的路径寻找和定位监测样地。
(2)根据样地的地形、植被和土地利用情况,确定中心点,用标杆进行标记,并尽量保证500m×500m调查样地在空间上是连续分布的,且100m×100m样地位于所调查植被的代表性地段通过。
(3)以中心点标杆为依据,通过罗盘和米绳确定东南西北4条250m的样线(每25m插1根旗杆),以及位于东北和西南对角线上距离中心点45m的两个种-面积和点格局考察点的位置,并用旗杆作标记;同时,进行种-面积曲线的调查和记录。
(4)调查人员从中心点沿样线开始样带记录;同时,进行样地中心点地标的埋设、样地位置及天气信息的记录、景观和工作照片的拍摄。
(5)调查的数据、照片和标本进行整理存档,并进行室内测定。
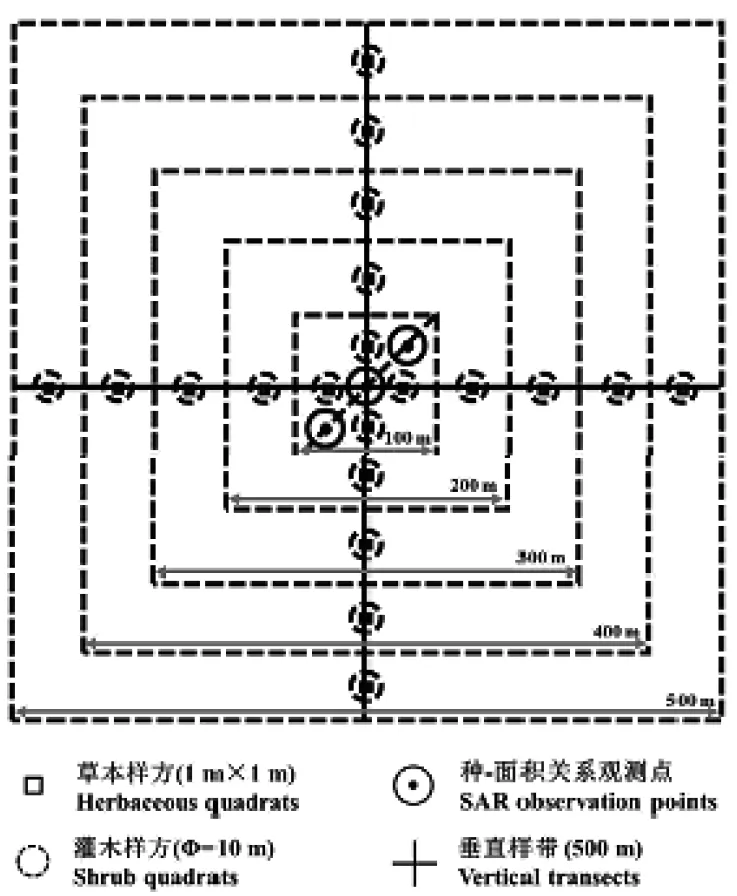
图2 单个监测样地上样线、样方、种-面积关系及
2.3 α多样性测度方法
本项研究采用了三类常用多样性的指数,即丰富度指数(richness index)、均匀度指数(evenness index)和多样性指数(diversity index)。其测度公式如下:
(1)物种丰富度指数采用Margalef(1958)指数:
式中,S为物种数目,N为所有物种个体总数。
(2)多样性指数采用Shannon-Wienner:H′=-∑PilnPi
式中,Pi是一个个体属于第i种的概率。
(3)均匀度指数
Pielou(1975,1977)均匀度指数(E1):
2.4 数据处理
本文数据使用spss17.0和excel2003进行统计分析。
3 结果与分析
3.1 种-面积关系
通过对图3中9个样地种-面积曲线图的分析比较,结果表明:一定区域内物种数量随面积的增加呈对数型增加,当面积达到一定程度时,取样面积再增加时,物种数量趋于平缓。从图中可以看出群落内的物种数与取样面积关系与对数模型S=alnA+b(式中,S为物种数目,A为取样面积,a、b为模型中的参数)能够很好拟合,并且对数模型中R2值≥0.9715。结果与白永飞、许志信〔10〕等关于内蒙古高原针茅草原群落α多样性研究结果一致。
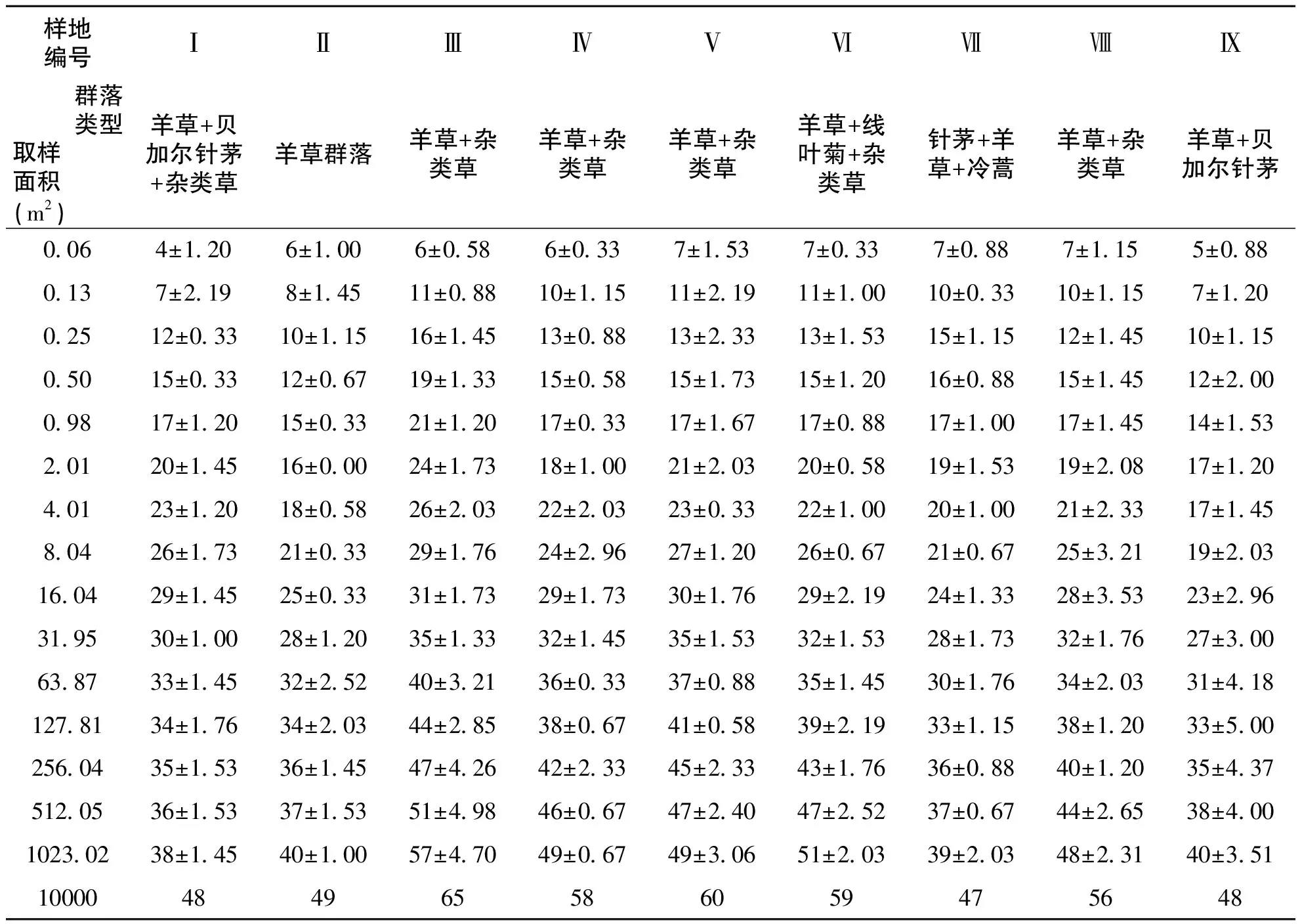
表2 群落物种数目与取样面积的关系
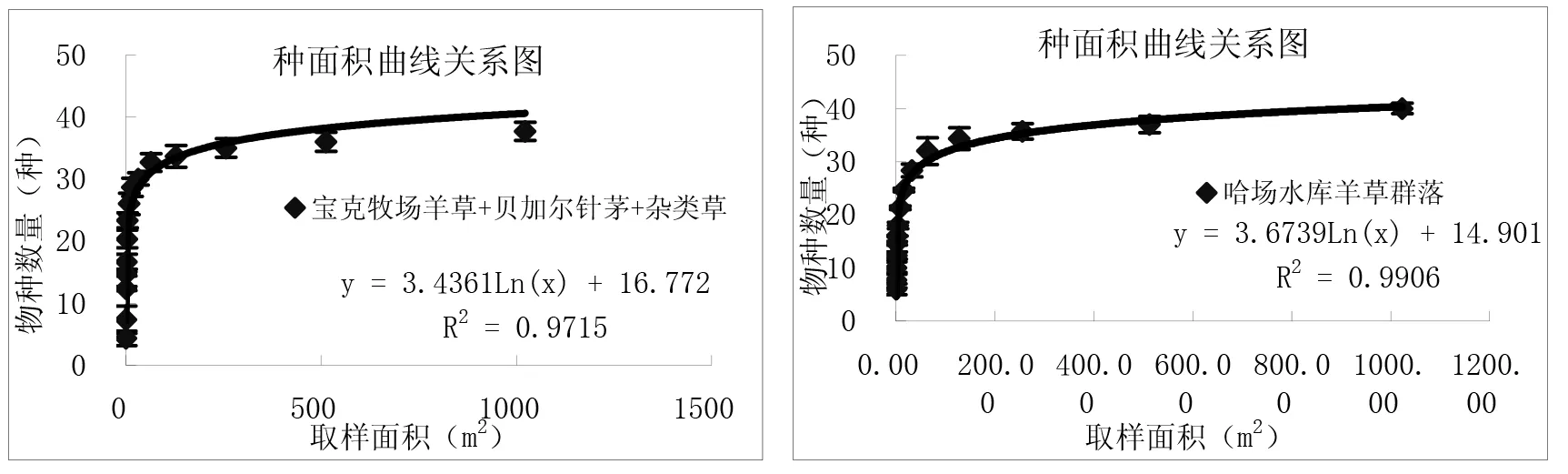
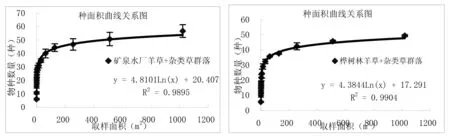
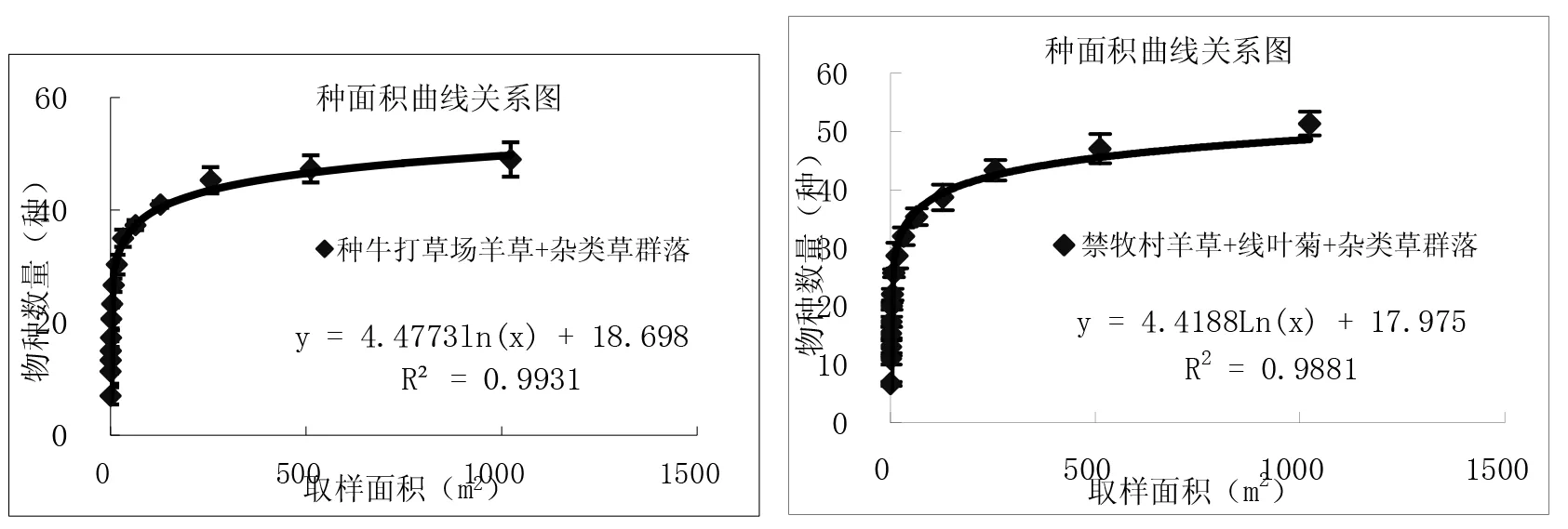
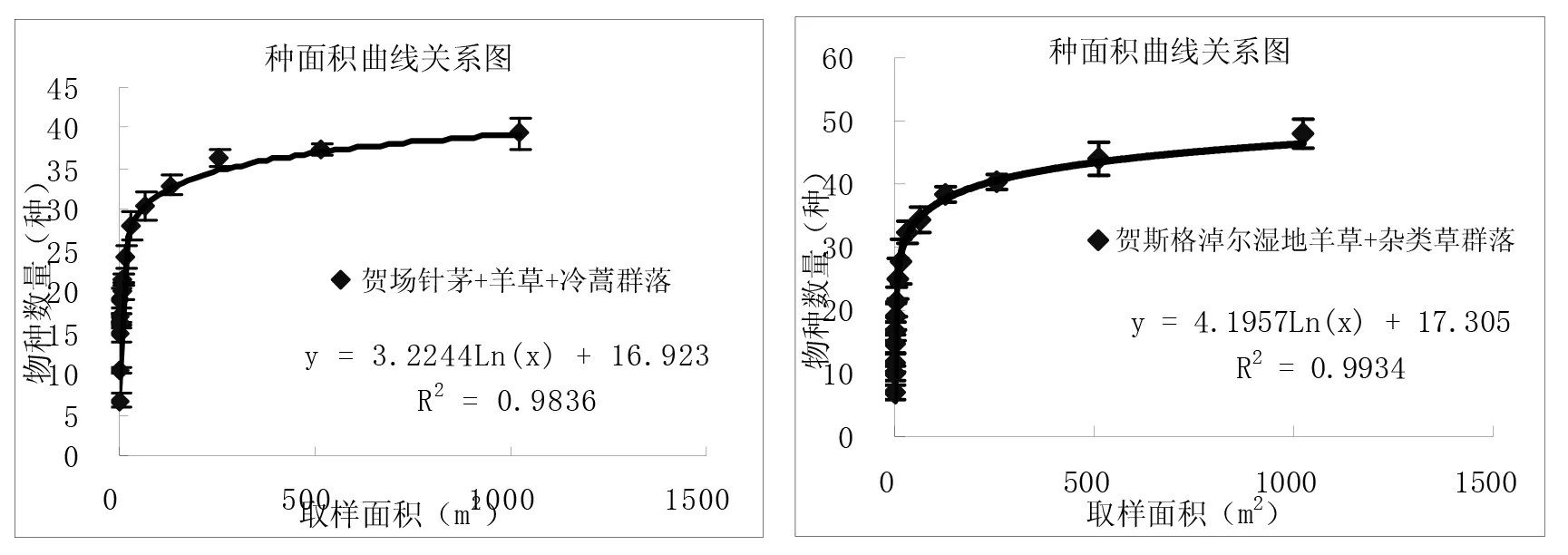
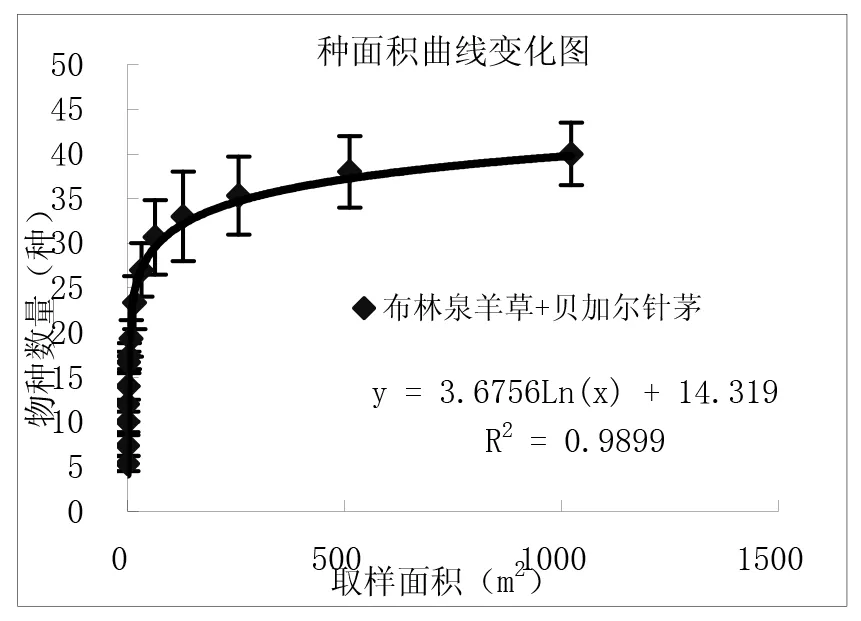
图3 各群落类型种-面积曲线关系图
3.2 不同群落类型及1m2物种数
不同群落类型的9个样地的单位面积内物种丰富度变化较明显,介于12-23种之间;其中位于禁牧村Ⅵ号样地的山地草甸类“羊草+线叶菊+杂类草”群落类型物种数最多,达23种,并且除了与Ⅳ号样地的山地草甸类“羊草+杂类草”群落类型无差异外与其它群落类型的样地都存在明显差异;位于哈场水库南的Ⅱ号样地低地草甸类“羊草群落”类型物种数最少为12种,除了与Ⅲ、Ⅶ、Ⅸ样地无差异外与其它群落类型样地都存在明显差异。这说明样地内1m2物种数量随草地类和群落类型的不同而存在差异(图4)。
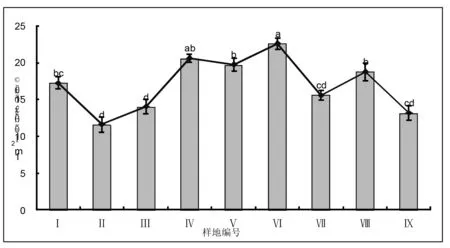
图4 不同群落类型样地单位面积内物种数
另外,调查中发现,样地中有草原类主要有“温性草甸草原类、低地草甸类、山地草甸类、温性典型草原类”;群落类型主要有“羊草+贝加尔针茅+杂类草、羊草+杂类草、针茅+羊草+冷蒿、羊草+线叶菊+杂类草、羊草+贝加尔针茅”。群落中的优势植物大多以禾草为主,禾草主要有羊草(Leymuschinensis(Trin.)Tzvel.)、贝加尔针茅(StipaBaicalensisRoshev.)、大针茅(StipagrandisP.Smirn.)、糙隐子草(Cleistogenessquarrosa(Trin.)Keng)、硬质早熟禾(PoasphondylodesTrin.)、披碱草(ElymusdahuricusTurcz.)等;另外一些亚优势种及伴生种主要为豆科、菊科和杂类草等,其中,豆科牧草主要斜茎黄芪(AstragalusadsurgensPall.)、胡枝子(LespedezabicolorTurcz.)、扁蓿豆(Medicagoruthenica(L.)Sojak.)、广布野豌豆(ViciacraccaL.)、草木樨状黄芪(AstragalusmelilotoidesPall.)、歪头菜(ViciaunijugaA.Br.);莎草科主要有寸草苔(CarexduriusculaC.AMey.)等;杂类草有线叶菊(Filifoliumsibiricum(L.)Kitam.)、芍药(PaeonialactifloraPall.)、防风(Saposhnikoviadivaricata(Trucz.)Schischk.)、红柴胡(BupleurumscorzonerifoliumWilld.)、石竹属(Dianthus)、委陵菜属(Potentilla)、麻花头(SerratulacentauroidesL.)、风毛菊属(Saussurea)、达乌里龙胆(GentianadahuricaFisch)、沙参(AdenophorastrictaMiq.)、地榆(SanguisorbaofficinalisL.)、华北蓝盆花(ScabiosatschiliensisGrun.)、小黄花菜(HemerocallisminorMill.)等,共调查记录了物种283种,隶属41科,152属,其中很多具有药用价值。
3.3 不同尺度下各样地内物种数
不同尺度下各样地物种数变化较明显,物种变化趋势呈现出基本是500×5m2>500×500m2>100×100m2>100×5m2>1m2的规律。说明物种数随着取样面积的增大而增大(图5)。
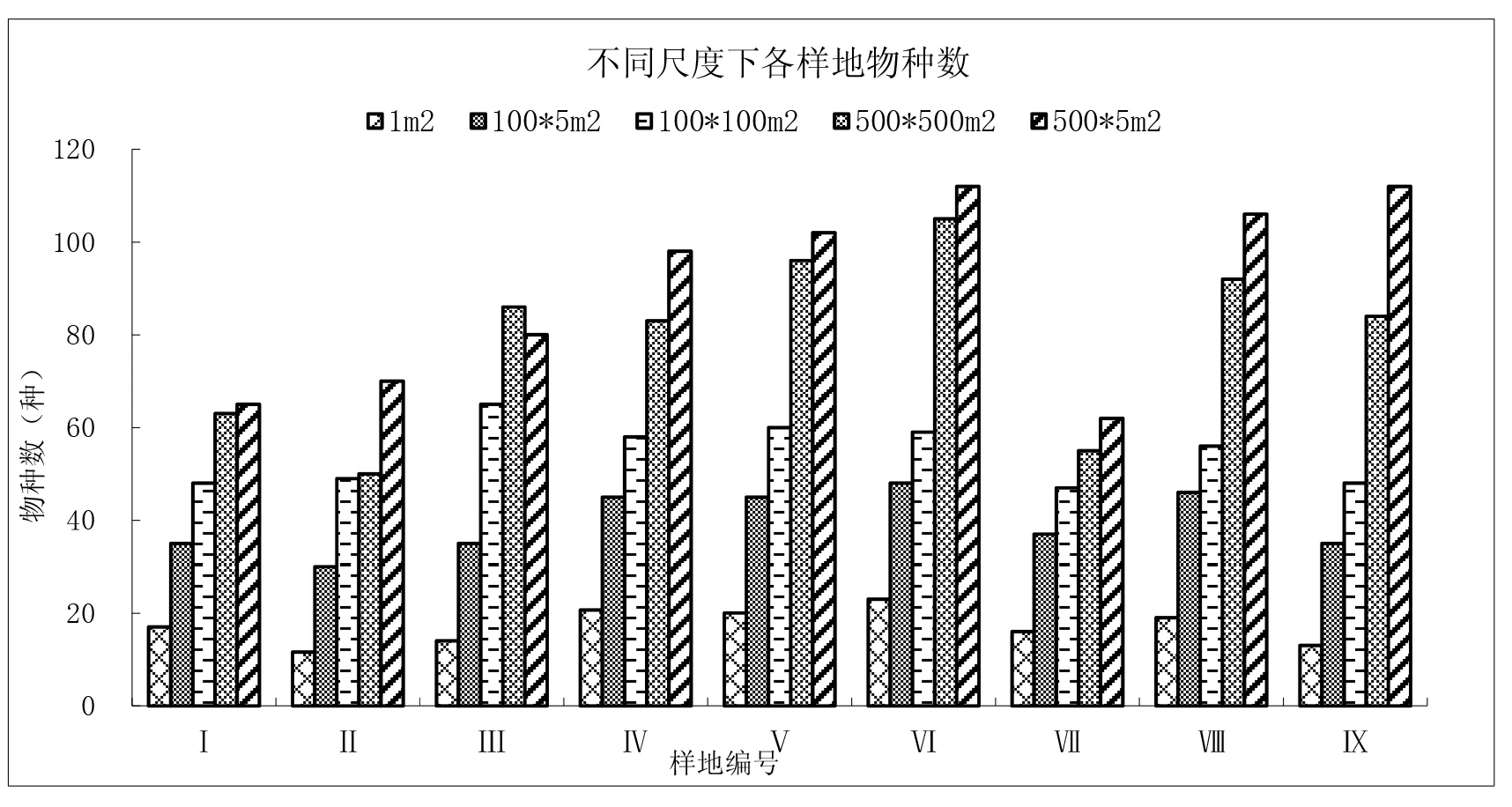
图5 不同尺度下各样地物种数
3.4 不同生境群落类型的α多样性特征
不同群落类型多样性指数见表3,样地Ⅴ的温性草甸草原类“羊草+线叶菊+杂类草”群落类型丰富度指数最高,达11.4959;样地Ⅵ的山地草甸类“羊草+杂类草”类型多样性指数最高达2.6098;样地Ⅱ的低地草甸类“羊草群落”类型的丰富度指数和多样性指数都最低分别为7.7446、1.5495;丰富度指数大小顺序为:Ⅴ>Ⅲ>Ⅵ>Ⅳ>Ⅷ>Ⅸ>Ⅰ>Ⅶ>Ⅱ;多样性指数大小顺序为:Ⅵ>Ⅴ>Ⅳ>Ⅷ>Ⅸ>Ⅰ>Ⅶ>Ⅲ>Ⅱ(表3)。
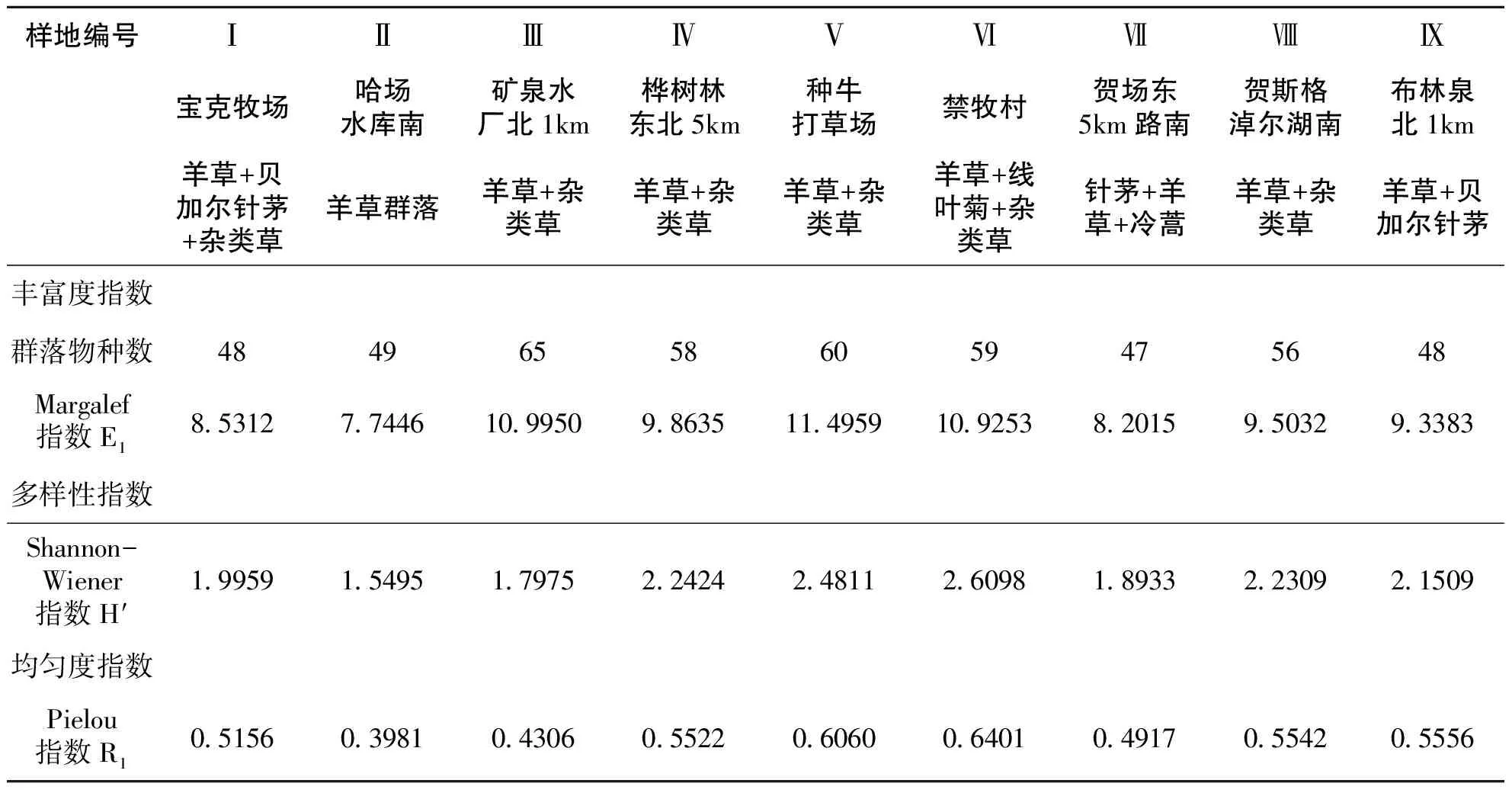
表3 各群落丰富度指数、多样性指数和均匀度指数
均匀度指数的结果表明,样地Ⅵ的山地草甸类“羊草+线叶菊+杂类草”类型、样地Ⅵ的山地草甸类“羊草+杂类草”类型、样地Ⅴ的温性草甸草原类“羊草+杂类草”类型、样地Ⅷ的低地草甸类“羊草+杂类草”类型、样地Ⅸ的温性草甸草原类“羊草+贝加尔针茅”类型具有较高的均匀度,其均匀度指数高于样地Ⅰ的“羊草+贝加尔针茅+杂类草”、样地Ⅶ的温性典型草原类“针茅+羊草+冷蒿”类型、样地Ⅲ的温性草甸草原类“羊草+杂类草”类型、样地Ⅱ的低地草甸类“羊草+杂类草”类型。其中位于样地Ⅵ的山地草甸类“羊草+线叶菊+杂类草”类型,均匀度指数最高,表明在所有这些样地中该样地个体数量分配较为均匀,而样地Ⅱ的低地草甸类“羊草+杂类草”群落均匀度指数最低,说明其个体数量分配不均,较其它样地稍差。具体大小顺序为:Ⅵ>Ⅴ>Ⅸ>Ⅷ>Ⅳ>Ⅰ>Ⅶ>Ⅲ>Ⅱ(表3)。
3.5 α多样性指数的相关性分析
相关分析结果表明,各种α多样性指数之间具有一定的正相关性(表4)
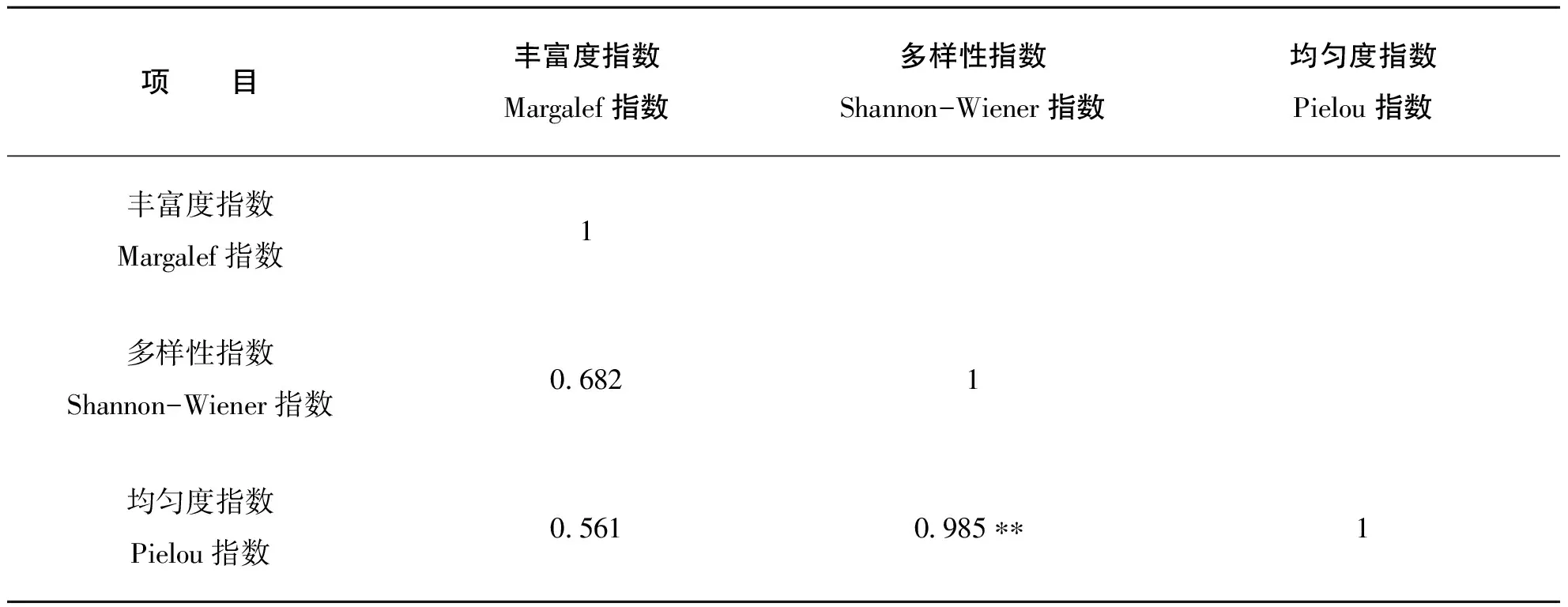
表4 α多样性指数间的相关分析
均匀度指数和其它多样性指数成正相关,说明均匀度是增加群落多样性的重要因子(物种数在一定范围的前提下)。Shannon-Wiener多样性指数与Pielou均匀度指数相关系数达到0.985,说明两者在描述群落多样性时有较高的一致性。Pielou均匀度指数与Shannon-Wiener多样性指数的相关性较高,而与Margalef丰富度指数相关系数相对较低,说明群落的多样性更多的受均匀度指数的影响,这与邱波,任青吉(2004)〔11〕和徐广平,张德罡等〔12〕研究结果一致(表4)。
4 结论
(1)群落内的物种数随取样面积的增大呈对数趋势增加,物种数量与取样面积关系与对数模型S=alnA+b(S为物种数目,A为取样面积,a、b为模型中的参数)能够很好拟合,并且对数模型中R2值≥0.9715。9个试验样地结果表明,在一定尺度范围内各群落的物种数目均随取样面积增加呈对数型增加。
(2)无论是用物种多样性指数(Shannon-Wiener指数),还是用物种丰富度指数或均匀度指数(Pielon)测度乌拉盖草原不同群落类型的α多样性,都反映出相似的结果,即温性草甸草原类的“羊草+线叶菊+杂类草”类型、温性草甸草原类的“羊草+杂类草”类型、山地草甸类“羊草+杂类草”类型具有较高的多样性指数,而低地草甸类的“羊草群落”类型拥有较低多样性指数。
(3)均匀度指数和丰富度指数和多样性指数成正相关,是增加群落多样性的重要因子(物种数在一定范围的前提下)。Margalef丰富度指数与Pielou均匀度指数在描述群落多样性时有较高的一致性,并且群落的多样性更多的受均匀度指数的影响。
〔1〕王小明,李明,盛和林.生物多样性与其保护意义〔J〕.大自然探索,1995,14(3)
〔2〕Tilman D,Knops J,Wedin D and Reich Petal .1997.The influence of functional diversity and composition on ecosystem processes.Science,277:1300~1302.
〔3〕Yachi S,Loreau M, 1999. Biodiversity and ecosystem productivity in a fluctuating environment: the insurance hypothesis. Proceedings of the National Academy of Sciences USA ,96: 1463-1468.
〔4〕Tilaman D.Downing JA.Bildibersity and stability in grasslands Nature 1994,367:363-365.
〔5〕Tilaman Wed in D.Knops J.Producitvity and sustalinability influenced by biodiversity in grassland ciosystems Nature,1996,379,718-720.
〔6〕Tilaman D,Reich P B.Knops J.Wedin D.Mielke T.Lwhman C.Diversity and productivity in a long-term grassland experiment Science,2001,294,843-845.
〔7〕Whittaker,R H Evolution of species diversity in land communities〔J〕.Evolutionary biology,1977,(10):167.
〔8〕王伯荪,彭少麟.植被生态学-群落与生态系统〔M〕.北京:中国环境科学技术出版社,1997,5-12.
〔9〕万宏伟,潘庆明,白永飞.中国草地生物多样性监测网络的指标体系及实施方案〔J〕.生物多样性,2013,21(6):639-650.
〔10〕白永飞,许志信,李德新.内蒙古高原针茅草原群落α多样性研究〔J〕.生物多样性,2000,8(4):353-360.
〔11〕邱波,任青吉,罗燕江,杜国祯.高寒草甸不同生境类型植物群落的α及β多样性研究〔J〕.西北植物学报,2004,24(4):655-661.
〔12〕徐广平,张德罡,徐长林,蒲小鹏,陈建纲.东祁连山高寒草地不同生境类型植物群落α及β多样性的初步研究〔J〕.草业科学,2006,23(6).
The α diversity research about different community types of WuLaGai steppe
Zhang Jian Xing qi Wang Jun-Fang Bai Yao-Hua Yao Xiao-Hong
(Monsod Drought-Resistance Co.,Ltd.Hohhot 010030)
The traits of α diversity of nine different plant communities in WuLaGai using comparable plot methods. The results showed that: (1) The number of species increase with the sampling area increases logarithmically, and with the logarithmic model S =alnA+bcan be a good fit (R2≥0.9715). (2) The α diversity index values of different community types(richness index, diversity index, evenness index), showed that mountain steppe“ Leymus chinensis(Trin.)Tzvel.) +Filifoliumsibiricum(L.)Kitam. + forbs”type,Warmsteppe“Leymuschinensis(Trin.)Tzvel.) + forbs” type、 mountain steppe “Leymuschinensis(Trin.)Tzvel.) + forbs” type, low plains steppe “Leymuschinensis(Trin.)Tzvel.) + forbs” type has a high diversity index, low plains steppe“Leymuschinensis(Trin.)Tzvel.) ”community types has lower diversity index. (3) community diversity influence more by evenness index.
community type;species - area relationships;α diversity
2015-05-08
朝克图(1964-),男,蒙古族,内蒙古通辽市人,高级畜牧师,从事草原保护建设及草原技术推广工作。
S812
A
2095—5952(2015)04—0009—16
