黄芩苷对热应激条件下LLC-PK1细胞HSP70 表达的影响
2015-03-18迟世凯孙春玲李华涛姜忠玲曹荣峰田文儒
迟世凯,孙春玲,李华涛,丛 霞,姜忠玲,王 新,曹荣峰,田文儒
(青岛农业大学动物科技学院,山东 青岛 266109)
中兽医认为,高温作为应激因子对机体产生热应激乃至造成热应激综合征,最后往往以肾功能衰竭而死亡[1]。黄芩苷(Baicalin)是从黄芩的干燥根中提取出的一种黄酮类化合物,是黄芩的主要有效成分之一,其解热作用显著[2],Tsai 等[3]研究认为黄芩苷可能通过抑制下丘脑中N-甲基-D-天冬氨酸受体依赖羟基旁路和TNF-α的增加从而发挥其解热作用。目前关于黄芩苷单体解热方面的研究甚少,其解热机制的研究也尚未深入,以黄芩为主药的中药复方解热机制方面的研究也不多,且复方研究结果不能代表单味药。现已证明,多种体外培养的细胞和胚胎受热后均能产生热休克蛋白(HSP),HSP70 是HSPs 家族里含量最多,亦是最重要的一种非特异性细胞保护蛋白,尤其对体外培养的细胞具有显著的抗热保护作用[4-5]。刘萍等[6]研究表明,黄芩苷能够促进大鼠脑缺血再灌注损伤后海马神经元HSP70 的表达,而黄芩苷能否调节热应激条件下的细胞合成HSP70,目前未发现有任何报道。因此,本研究拟探讨不同浓度黄芩苷对热应激条件下猪肾小管上皮(LLC-PK1)细胞HSP70 表达的影响,以期为黄芩苷的解热作用机制提供理论基础。
1 材料与方法
1.1 主要试剂 黄芩苷,购自北纳创联生物技术研究院;高糖DMEM和胰蛋白酶,购自GIBCO公司;胎牛血清,购自HyClone 公司;RNA提取试剂盒,购自原平皓生物有限公司;反转录试剂盒、rTaqDNA聚合酶和DL-1 000 Marker,购自TaKaRa公司;LightCycler®480 SYBR Green I Master,购自Roche 公司;HSP70、GAPDH 抗体和兔抗羊IgGHRP 分别,购自Santa Cruze 公司和Jackson ImmunoResearch 公司;BLUeye Prestained Protein Ladder,购自GeneDireX 公司;Western 及IP 细胞裂解液、一抗二抗去除液和ECL 荧光底物试剂盒,购自碧云天生物制品有限公司。
1.2 引物设计与合成 根据GenBank 中收录号为NM001123127的猪HSP70 基因序列和XM003357928猪β-Actin基因序列设计引物,并由上海生工生物工程技术服务有限公司合成。序列如下:HSP70 Sense primer:5′-GCCCTGAATCCGCAGAATA-3′,Anti-sense primer:5′-TCCCCACGGTAGGAAACG-3′,扩增片段长度为152 bp;β-Actin:Sense primer:5′-CGGGACCTGACCGACTACCT-3′,Anti-sense primer:5′-GGCCGTGATCTCCTTCTGC-3′,扩增片段长度为411 bp。
1.3 细胞培养与试验分组 LLC-PK1 细胞为美国ATCC细胞库第3 代细胞,购自上海复祥生物科技有限公司。细胞用含12%胎牛血清的高糖DMEM在37 ℃、5% CO2的饱和湿度下培养,待细胞覆盖率达到80%~90%后传代,传代细胞用于后续试验。
试验分为Ⅰ组37 ℃常温对照组,Ⅱ组42 ℃单纯热应激1 h 组,而Ⅲ组、Ⅳ组、Ⅴ组、Ⅵ组和Ⅶ组分别添加0.01 μg/mL、0.1 μg/mL、1 μg/mL、10 μg/mL 和100 μg/mL 不同浓度黄芩苷培养24 h 后再将其42 ℃热应激1 h 组。
1.4 cDNA合成与PCR 扩增电泳 按照RNA快速提取试剂盒(原平皓生物有限公司)说明书,提取各组细胞总RNA,按照反转录试剂盒(TaKaRa 公司)说明书,进行RT 反应。将合成的cDNA进行PCR 扩增,产物经2%琼脂糖凝胶(含0.5 g/L 溴化乙锭)电泳,由GDS-8000 凝胶成像分析系统拍照鉴定。
1.5 荧光定量PCR 检测HSP70 基因的表达 将合成的cDNA产物做一系列浓度梯度稀释,使用Roche LightCycler 480Ⅱ荧光定量PCR 仪进行定量分析。反应体系为10 μL,其中包括:ddH2O23.6 μL,SYBR Green Realtime PCR Master Mix 5 μL,上下游引物各0.2 μL,cDNA1 μL。反应条件为95 ℃10 min,95 ℃5 s 和58 ℃30 s,共45 个循环。
1.6 Western Blot 检测HSP70 蛋白的表达 提取细胞总蛋白,混合2×SDS样品缓冲液加热变性后进行SDS-PAGE 凝胶电泳,4℃条件下220 mA恒流湿转2 h,再封闭2 h,按顺序加入一抗二抗孵育后,用ECL 试剂盒进行化学发光,最后用UVP 凝胶成像系统拍照检测,用AlphaView对试验结果进行灰度值分析。
1.7 数据统计分析 采用SPSS16.0 软件做统计学处理,各组数据进行LSD多重比较,均数间进行t检验。
2 结果
2.1 PCR 扩增 利用设计的HSP70 和β-Actin 引物进行PCR 扩增,在152 bp 处扩增到了一条HSP70 的条带,在411 bp 处扩增到了一条β-Actin的条带,与预期值相符合,经基因测序证实其为目的基因产物。
2.2 HSP70 mRNAReal-time PCR 结果 各组细胞以β-Actin 做内参基因,用2-△△Ct法进行相对定量分析后发现(图1),与37 ℃常温对照组相比,其他各组LLC-PK1 细胞HSP70 mRNA的表达量均升高,其中Ⅵ组和Ⅴ组差异极显著(P<0.01),其余各组差异显著(P<0.05);与42 ℃单纯热应激组相比,除Ⅰ组外,其余各组LLC-PK1 细胞HSP70 mRNA的表达量均升高,其中Ⅴ组差异极显著(P<0.01),Ⅲ,Ⅳ,Ⅵ租和Ⅶ组差异显著(P<0.05)。
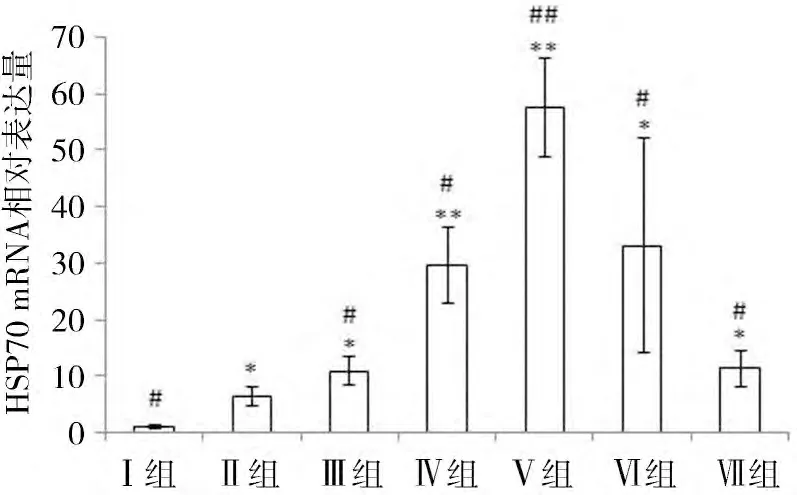
图1 各组LLC-PK1细胞HSP70 m RNA相对表达量

图2 HSP70和GAPDH蛋白电泳

图3 各组LLC-PK1细胞HSP70 蛋白相对表达量
2.3 HSP70 蛋白Western Blot 结果 Western Blot检测蛋白表达的结果(图2)用AlphaView软件扫描其灰度,并对数据进行统计分析发现(图3),与37 ℃常温对照组相比,其他各组LLC-PK1 细胞HSP70 蛋白的表达量均升高,其中Ⅴ组差异极显著(P<0.01),其余各组差异显著(P<0.05);与42℃单纯热应激组相比,除Ⅰ组外,其余各组LLCPK1 细胞HSP70 蛋白的表达量均升高,但Ⅲ组和Ⅶ组差异不显著,Ⅴ组差异极显著(P<0.01),Ⅳ组和Ⅵ组差异显著(P<0.05)。
3 讨论
细胞在受到高温、缺氧及重金属等应激源刺激时,细胞过表达HSP70,而HSP70 能够增加细胞自身对损害的耐受力,维持细胞正常功能代谢,提高其存活率[7]。因此,很多研究者尝试过许多方法来增加细胞中HSP70 的表达[8-9],从而提高细胞抗应激的能力,但在培养液添加中草药的单一成分来提高细胞的HSP70 表达的报道甚少,添加黄芩苷的尚无报道。
据报道黄芩苷具有清热解毒、抗菌消炎、抗氧化、降压,及镇静等作用[10]。而李倩楠[11]曾经报道,高剂量黄芩苷(160 mg/kg)灌服能明显降低发热大鼠的体温,并认为黄芩苷可能通过抑制TNFA和IL-1B的产生或释放,继而引起IL-6 的改变,再使PGE2 合成减少,从而发挥解热作用。赵红艳[12]等研究表明,黄芩苷可以通过降低下丘脑中PGE2 和cAMP 的含量来起到解热作用。
本试验42 ℃单纯热应激1 h 能显著诱导LLC-PK1 细胞HSP70 的表达,说明42 ℃的温度能够引起细胞的热休克反应。与37 ℃常温对照组相比,在LLC-PK1 细胞培养液中添加的黄芩苷在1 μg/mL 以 下(0.01~1 μg/mL)时,细胞表达HSP70 的量随黄芩苷浓度增加而升高,而当黄芩苷浓度在1 μg/mL 以上(1~100 μg/mL)时,虽然细胞HSP70 表达呈降低趋势,但仍显著高于对照组,这说明黄芩苷(0.01~100 μg/mL)能够上调热应激条件下LLC-PK1 细胞HSP70 的表达。与42 ℃单纯热应激组相比,黄芩苷(0.01~100 μg/mL)均能显著增加细胞HSP70 mRNA的表达,但0.01 μg/mL 和100 μg/mL 黄芩苷对细胞HSP70 的蛋白表达无显著性影响,首先,有研究表明[13],真核基因表达的转录和翻译发生的时间和位点存在时空间隔,转录和翻译还受到多种因素调控,因此mRNA的表达水平并不一定代表蛋白的表达水平;其次,极有可能因为0.01 μg/mL 的黄芩苷浓度太小,不足以诱导热应激条件下细胞HSP70 蛋白的表达,而100 μg/mL 的黄芩苷的浓度又过大,抑制了热应激条件下细胞HSP70 蛋白的持续高表达,所以,试验成功筛选出黄芩苷诱导热应激条件下LLCPK1 细胞HSP70 表达的最佳浓度范围,即0.1~10 μg/mL,且1 μg/mL 的黄芩苷诱导效果最显著。
[1]刘钟杰,许剑琴.中兽医学[M].北京:中国农业出版社,2002:453-455.
[2]梁英,韩鲁佳.黄芩中黄酮类化合物药理学作用研究进展[J].中国农业大学学报,2003,8(6):9.
[3]Tsai CC,Lin MT,Wang JJ,et al.The antipyretic effects of baicalin in lipopolysaccharide-evoked fever in rabbits[J].Neuropharmacology,2006,51(4):709-717.
[4]Aréchiga CF,Ealy AD,Hansen P J.Evidence that glutathione is involved in thermotolerance of preimplantation murine embryos[J].Biology of Reproduction,1995,52:1296-1301.
[5]Gifondorwa DJ,Robinson MB,Hayes CD,et al.Exogenous delivery of heat shock protein 70 increases lifespan in a mouse model of amyotrophic lateral sclerosis[J].The Journal of Neuroscience,2007,27(48):13173-13180.
[6]刘萍,菊英,李倩,等.黄芩苷对大鼠脑缺血再灌注损伤后海马神经元HSP70 表达的影响[J].药学学报,2006,41(7):619-624.
[7]Radons I,Multhoff G .Immunostimulatory functions of membrane-bound and exported heat shock protein 70[J].Exerc Immu-nolRev,2005,11:17-33.
[8]Roesslein M1,Schibilsky D,Muller L,et al.Thiopental protects human T lymphocytes from apoptosis in vitro via the expression of heat shock protein 70[J].Journal of Pharmacology and Experimental Therapeutics,2008,325(1):217-225.
[9]Chu E K,Ribeiro SP,Slutsky AS.Heat stress increases survival rates in lipopolysaccharide-stimulated rats[J].Critical Care Medicine,1997,25:1727-1732.
[10]孙先枝,程建波,卜登攀,等.黄芩苷的生物学功能和黄芩及其提取物在畜禽生产中的应用研究进展[J].动物营养学报,2013,25(7):1459-1464.
[11]李倩楠,葛晓群.黄芩苷的解热作用及对细胞因子的影响[J].中国中药杂志,2010,35(4):1068-1072.
[12]赵红艳.黄芩苷对发热大鼠下丘脑PGE2 和cAMP 含量的影响[J].中国应用生理学杂志,2002,18(2):139-141.
[13]de Sousa Abreu R,Penalva L O,Marcotte E M,et al.Global signatures of protein and mRNAexpression levels[J].Molecular Bio-Systems,2009,5(12):1512-1526.
