互花米草生物量变化对盐沼沉积物有机碳的影响
2015-03-13冯振兴高建华汪亚平高建慧白凤龙
冯振兴,高建华,2,*,陈 莲,汪亚平,高建慧,白凤龙
1 南京大学, 海岸与海岛开发教育部重点实验室, 南京 210093 2 海洋沉积与环境地质国家海洋局重点实验室, 青岛 266061 3 盐城工学院环境科学与工程学院, 盐城 224051 4 国土资源部海洋油气与环境地质重点实验室,青岛海洋地质研究所, 青岛 266071
互花米草生物量变化对盐沼沉积物有机碳的影响
冯振兴1,高建华1,2,*,陈 莲1,汪亚平1,高建慧3,白凤龙4
1 南京大学, 海岸与海岛开发教育部重点实验室, 南京 210093 2 海洋沉积与环境地质国家海洋局重点实验室, 青岛 266061 3 盐城工学院环境科学与工程学院, 盐城 224051 4 国土资源部海洋油气与环境地质重点实验室,青岛海洋地质研究所, 青岛 266071
以江苏王港典型互花米草(Spartinaalterniflora)盐沼湿地为研究对象,分析光滩及互花米草滩沉积物中有机碳的水平和垂向分布特征,了解互花米草生物量的季节动态变化,探讨二者之间的相互关系,在此基础上研究互花米草生物量分布和季节变化对沉积物中有机碳(TOC)含量的影响。结果表明,互花米草枯落物中的有机碳数量在两个月内衰减了40%,而表层沉积物中TOC含量及其中互花米草来源TOC所占比例的变化,均与互花米草地表枯落物量的季节变化存在两个月的“相位差”,这与枯落物快速分解时间大致吻合,说明枯落物是表层沉积物中TOC的重要来源。高达60%的互花米草地下生物量分布在0—20cm深度内,该深度范围内沉积物中TOC含量较高,且TOC主要来源于互花米草。此外,不同深度TOC含量与地下生物量之间存在良好的正相关关系,说明地下生物量是影响沉积物TOC含量的重要因子。研究区互花米草年固碳能力为2274g m-2a-1,盐沼沉积物中TOC埋藏速率达到了470 g m-2a-1,是地表一个重要的碳汇;同时研究区每年向近岸水域输出大量的TOC,是近岸海域生态系统的一个重要碳源。
有机碳;互花米草;生物量;季节动态
The response of organic carbon content to biomass dynamics inSpartina
湿地是地球上生物多样性最丰富、生产力最高、生态功能最齐全的生态系统,具有极高的初级生产力和强大的固碳能力[1]。尽管湿地面积仅占全球陆地面积的4%—5%,但其碳储量可达陆地生态系统总量的35%,是地球上最大的碳库之一[2]。盐沼湿地是一种重要的湿地类型,大量的外来有机质在此积聚,加之本地较高的初级生产力、持续的沉积物堆积和较低的腐化分解速率,使得盐沼湿地有着很高的碳沉积速率和固碳能力[3- 5]。然而,由于受到自然变化和人类活动的影响,特别是生物入侵导致的生态系统结构和功能的转变,盐沼湿地碳循环又具有明显的时空变化特征,从而导致碳储量存在很大的不确定性。有机碳作为盐沼湿地生态系统物质循环的一个关键成分,是维持湿地生态系统高生产力和高生物量的基础,因而揭示盐沼湿地有机碳存储变化,对全球碳收支及与之有关的全球气候变化研究有着重要意义。
互花米草(SpartinaalternifloraLoisel)于1982年被引种到江苏沿海[6],凭借极强的适应性和繁殖能力,逐渐取代碱蓬(Suaedasalsa)成为当地优势物种,形成大面积的互花米草盐沼湿地,并对盐沼生态系统产生了重要影响[7- 10]。植被种群的改变势必会引起盐沼土壤有机碳库的变化,互花米草引种改变了盐沼有机碳输入模式,极大地增加了盐沼有机碳的累积,对此已有很多研究报道[11- 15]。然而,关于互花米草盐沼有机碳的累积机制,尤其是根、茎、叶及枯落物等不同植物组织,在盐沼有机碳埋藏和分解中的作用,尚缺乏深入研究。本文选择小区域的盐沼湿地作为研究对象,通过分析盐沼沉积物和植被中的TOC含量及其季节变化规律,探讨互花米草对盐沼湿地有机碳含量动态变化的影响,并初步估算苏北互花米草盐沼湿地的固碳能力。
1 研究地区与研究方法
1.1 研究区概况
研究区位于江苏海岸带中部王港河口潮滩湿地(33°11.2′N,120°45.5′E),受东海前进潮波和南黄海旋转潮波系统的控制,以潮汐作用为主,为典型半日潮,平均潮差为3.68m,波浪作用小,发育典型的淤泥质潮滩[16](图1)。该地处于北亚热带季风气候区,夏季盛行东南风,冬季盛行偏北风,年平均气温12.6℃,年均降水量1040mm。王港河河口段受潮汐作用影响强烈,采样点位于口门上游5km处,两岸自河向陆依次分布光滩和互花米草滩。由于围垦后大片土地被开发为鱼塘,河堤被人为填高,芦苇滩(Phragmitesaustralis)只在河堤地势较高处留有斑点状残余。
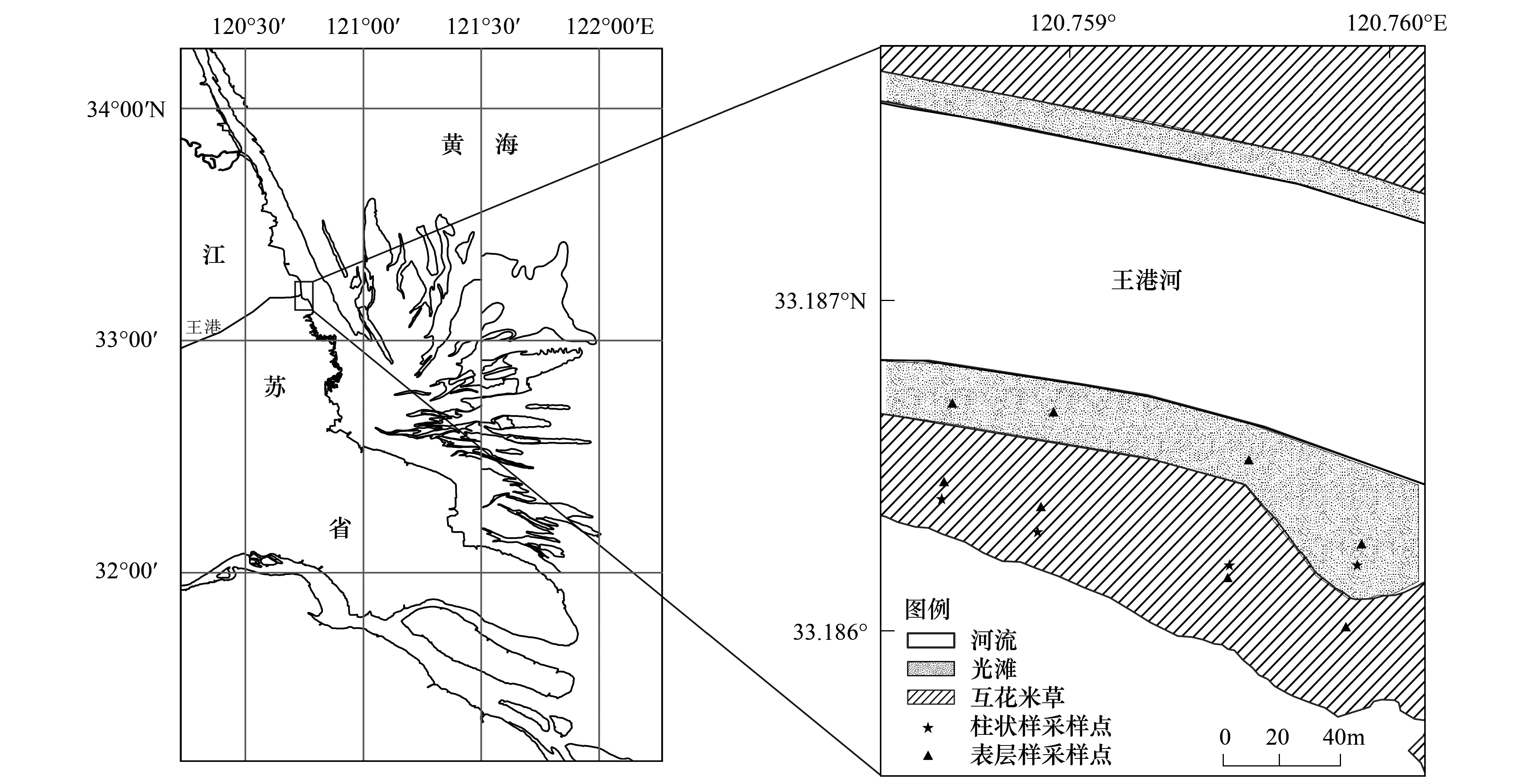
图1 研究区和采样点分布图Fig.1 Location of study area and sampling sites
1.2 研究方法
1.2.1 数据采集
自2012年8月至2013年6月(每两个月1次,分6次),在研究区采集沉积物表层样(图1)。共4条断面,每条断面每次分别在光滩和互花米草滩定点采集表层2cm样品各1个,共8个。沉积物样品带回实验室后,立即冷冻保存。互花米草植被生物量调查:在采集沉积物的同时,每次随机选择3个1m×1m的样方,采用收割法获取地上茎叶样品,作为地上生物量;在样方内,收集所有的互花米草地表凋落物,作为样方的互花米草枯落物量;同时挖取部分地下根系。2013年6月于河口段20km内均匀设置6个点,分别采集涨潮、落潮水样,现场过滤后冷冻保存,带回实验室分析。
由于潮滩工作环境艰难复杂,且相对地上部分,地下生物量年内变化不大[17],因此本文使用8月份数据代表年平均地下生物量。2012年8月,在采集地上生物量的样方点,按照1m×1m×1m规格,采集地下五层根系(0—20,20—40,40—60,60—80和80—100cm),获取地下生物量的分布特征。采用Wider和Lang提出的“Litterbag method”[18]测量植物残体腐烂分解速率:将死亡的地下根装入尼龙袋,袋子大小为20cm×20cm,网眼为100目,每袋装入15g(干重),共6袋,2012年8月将尼龙袋埋于盐沼湿地表层,以后每两个月取出1袋,带回实验室处理。
为了调查滩面沉积物的垂向变化规律,于2012年8月采集了4根柱状样(WG1—WG4),其中WG1—WG3分别位于断面01—03的互花米草滩,柱长为93、103cm和97cm;WG4位于断面04的光滩,柱长82cm。沉积速率采用“双桩法”[19]测得,具体操作即在潮滩上插入两根细木桩,间距1m,两桩均高出滩面相同高度,测量时在两桩上架一根横梁,测量横梁中心点到滩面的垂直距离,该距离的变化即可记录滩面的沉积速率。
1.2.2 样品处理
植物样品在50℃下烘干至恒重,称重法测定互花米草生物量,植物根、茎、叶、枯落物于ALPHA2124型冷冻干燥机冻干后,分别用球磨仪(RestchRM200)研磨过100目筛,测定有机碳(TOC)和总氮(TN)含量。此外,植株根、茎、叶同时测定同位素碳(δ13C)含量。自尼龙袋中取出的死亡根和其他植物样品一样,先称得质量变化后,再测试TOC含量。
沉积物样品,先用10%的稀盐酸去除样品中的碳酸盐,再用去离子水反复洗致中性后冻干。取适量样品,研磨至100目以下,立即上机测定TOC、TN和δ13C含量。水样同上。
实验中TOC、TN含量采用Leeman CE440元素分析仪测定,测试精度为0.1%。同位素δ13C采用DELTAplus Advantage同位素质谱仪测定,分析精度为0.1‰。沉积物粒度数据则由英国Malvern公司生产的Mastersizer 2000型激光粒度仪测得。
1.2.3 不同来源有机碳计算
盐沼沉积物中的有机碳除了受到本地互花米草生长的影响,还与外源有机碳输入有关。由于所有外源有机碳均是潮水涨落所带来的,因此本文假定,以水样中颗粒态有机碳δ13C值(≈-27.97‰)作为外源有机碳代表值,以根、茎、叶中δ13C平均值-13.55‰作为δ13Cspar代表值。依据端元线性混合模型[20- 21](式1),可计算出盐沼沉积物中互花米草来源TOC含量及相对比例:
δ13Csedi=f×δ13Cspar+(1-f)×δ13Cwater
(1)
式中,δ13Csedi和δ13Cwater分别代表样品和外源有机碳的δ13C值,f为互花米草来源有机碳所占比例。
2 结果与讨论
2.1 互花米草生物量季节动态变化

图2 互花米草地上生物量季节分配Fig.2 Seasonal variation of the aboveground biomass allocation of S. alterniflora (Mean ± SD)
夏季互花米草生长旺盛,地上生物量不断增大,至10月达到2596g/m2,为全年最高(图2)。此后逐渐下降,来年4月降至全年最低值,为1442g/m2。4—6月气温回升,新生互花米草快速生长,地上生物量又开始增大。从植株不同部位数据来看,茎、叶也表现出相同的变化规律。8—12月地表枯落物量逐渐下降,12月份为全年最低,2月份植株已全部枯黄,但仍然挺立,枯落物量与12月份基本相当,约170g/m2(图3)。此后枯立物开始大量凋落,到6月份达到最大值,是12月份的1.25倍。
互花米草具有发达的地下根系,地下生物量随着深度的增加呈指数衰减(图3),其中0—20cm和20—40cm深度内的生物量分别占地下生物量的60%和33%,依次达到2896g/m2和1312g/m2。由此可见,地下生物量主要分布在表层40cm深度内,尤其是0—20cm。
2.2 互花米草枯落物有机碳衰减规律
分解实验表明,枯落物中剩余有机碳数量与分解时间存在较好的指数衰减关系(图4)。最初的2个月内(8—10月),埋藏地下根茎体内有机碳数量减少了40%。此后,随着时间的逐渐延长,有机碳数量缓慢减少,基本保持稳定。

图3 地表枯落物量季节分配与地下生物量分布Fig.3 Seasonal variation of surface litter mass of S. alterniflora and Distribution of root biomass within a depth of 100cm for S. alterniflora

图4 有机碳分解曲线和不同深度生物量与有机碳含量的关系Fig.4 The content of TOC loss for litter bags and Relationship between belowground biomass and the content of TOC in different depth
2.3 表层沉积物有机碳季节变化及其与枯落物的关系
2.3.1 表层沉积物中TOC含量的季节变化
互花米草滩和光滩不同月份表层沉积物中TOC含量表明,两者均存在明显的季节变化(表1)。

表1 光滩和互花米草滩表层沉积物有机碳含量季节动态/%Table 1 Seasonal variation of the content of Total Organic Carbon (TOC) in the surface sediments of the bare and S.alterniflora flat
其中,互花米草滩TOC最大含量出现在8月份,平均值为1.31%,2月份TOC含量出现最小值,其他时间内含量变化不大。光滩TOC在6月份含量最高,平均含量为0.66%,12月份含量较低。互花米草滩TOC含量在各个月份均高于光滩,年平均含量为1.08%,为光滩的2.19倍。
2.3.2 表层沉积物中TOC含量与枯落物量的关系

图5 地表枯落物、表层沉积物有机碳含量及其中互花米草贡献比例三者的对照Fig.5 Comparison between surface litter mass, the content of TOC in surface sediments and the percentage of TOC derived from S. alterniflora
植物枯落物及其分解是维持盐沼生态系统营养元素循环的重要环节,也是沉积物中有机碳最重要的来源之一[22]。前人研究表明,湿地枯落物的分解最初是可溶性成分的淋溶过程,碳和氮在数周内即可大量损失[23- 24]。而本文的研究结果也表明,研究区互花米草枯落物的质量在两个月内即减少了44%,有机碳数量衰减了40%。因此,对短时间尺度而言,枯落物快速分解释放的有机碳,必然会引起表层沉积物中TOC含量的变化。本文研究发现,互花米草滩表层沉积物中TOC含量与地表枯落物量季节变化之间呈现出较好的一致性,但二者存在明显的“相位差”(图5)。4—8月地表枯落物量较高,而TOC含量相对较高的时间是6—10月,其中6月枯落物量达到最大,但表层沉积物中TOC含量最高值出现在8月,TOC含量变化比枯落物量变化延迟两个月,这恰好与枯落物快速分解时间一致。说明枯落物量的变化,显著地影响了表层沉积物TOC含量。此外,研究区表层沉积物粒度季节变化不大,且与TOC含量之间无明显线性关系(P>0.05)。另外,光滩表层TOC含量季节变化与互花米草滩明显不同,表明了由枯落物引起的有机碳来源变化是导致互花米草滩表层TOC含量变化的主要原因。同时,表层沉积物中互花米草来源TOC所占比例,其全年变化曲线与枯落物量变化走向一致,但同样存在两个月的时间滞后现象。说明枯落物是表层沉积物有机碳的重要来源,其分解产生的有机碳输入,是影响表层沉积物中TOC含量的重要因子。
2.4 沉积物有机碳的垂向分布及其与地下生物量的关系
2.4.1 沉积物中TOC、TN和δ13C的垂向分布特征
柱样WG1—WG4中的TOC和TN含量,以及C/N、δ13C、平均粒径和互花米草来源有机碳所占比例的垂向变化如图6所示。其中WG1—WG3柱样各指标数值差异不大,TOC含量依次介于0.12%—1.70%、0.09%—1.76%和0.09%—1.56%,TN含量分别为0.014%—0.148%、0.011%—0.157%和0.012—0.129%。TOC和TN含量的峰值一般出现在亚表层(2.5—5cm)处,亚表层向下至20cm深度内TOC和TN含量下降较快,20cm深度以下,TOC和TN含量较低且变化较小。光滩柱样WG4的TOC和TN含量表现出了与互花米草滩不同的垂向变化特征:自表层至30cm,TOC和TN含量大致稳定,平均值分别为0.49%和0.058%,但从30cm向下,TOC和TN含量逐渐降低。另外,柱样WG1—WG3的C/N值也呈现出沿深度先减小后稳定的趋势,WG4柱样C/N值自上而下变化不大,基本维持在8.37左右。
柱样WG1—WG3的δ13C值分别为-24.23‰—-18.03‰、-23.12‰—-18.36‰和-23.85‰—-18.42‰,δ13C值大致随深度呈减小趋势,主要集中在0—20cm深度内,20cm以下,除去个别波动,基本稳定在平均值-22.02‰左右。WG4整柱δ13C值变化不大,平均值为-23.54‰。
互花米草滩表层沉积物有粗化现象,WG1柱在5—40cm深度内变化不大,40cm以深逐渐变粗。WG2整柱波动较大,在30cm和60cm上下较小范围内有明显细化的波动,10cm以下总体呈现变粗趋势。WG3柱15cm以下大致逐渐变粗。WG4粒径波动主要集中在表层20cm内,下部变化不大。相比互花米草滩,光滩沉积物更细。
2.4.2 沉积物TOC垂向分布与互花米草地下生物量的关系
沉积物中TOC含量是有机碳输入与微生物分解输出相平衡的结果,有机碳的输入是影响TOC含量垂向变化的主要因素之一。地表枯落物量相对地下生物量小很多,且主要影响表层沉积物,因此沉积物TOC含量垂向变化更多地取决于互花米草地下生物量的分布。本文互花米草滩沉积物中TOC含量与互花米草地下生物量垂向分布的相互关系可以看出:首先,二者在变化形态上表现出了良好的一致性(图3,图6);其次,0—20cm深度内TOC含量较高,且随着深度的减小,互花米草来源TOC含量不断增大,互花米草来源TOC的贡献逐渐提高;第三,对不同深度地下生物量与沉积物中TOC含量的回归分析也表明,二者存在显著正相关(图4)。由此可见,地下生物量对沉积物TOC含量变化有着重要影响。
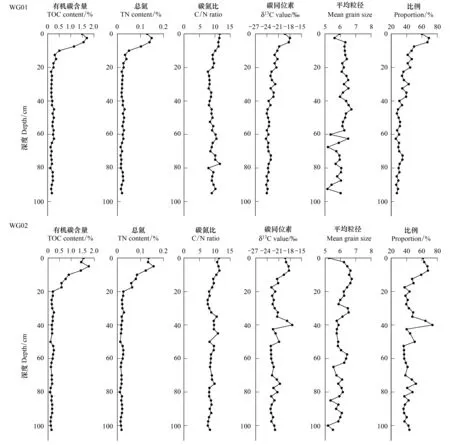

图6 柱状WG1—WG4中有机碳含量、总氮、碳氮比、碳同位素值、平均粒径和比例(互花米草来源有机碳所占比例)的垂向变化Fig.6 Vertical variations in TOC,TN content, C/N ratio,δ13C value, mean grain size and proportion (the proportion of TOC derived from S. alterniflora) in the cores WG1—WG4
另外,沉积物中的δ13C和C/N值的垂向变化也表明了互花米草对沉积物有机碳的巨大贡献。C/N可用来判断沉积物有机质来源变化,通常认为陆源有机质的C/N>12,而海源的为6—9[25]。互花米草滩柱样0—20cm深度内C/N介于9.30—12.56,来源显著偏向于陆地高等植物。且相应柱样中的δ13C值分布也表明,δ13C值主要在0—20cm范围内迅速减小,之后基本保持稳定。本研究测得互花米草根、茎、叶中的δ13C值介于-13.89‰—-13.27‰,而0—20cm内的δ13C平均值为-19.90‰,显著偏重,明显受到互花米草地下根茎的影响。因此,互花米草是沉积物有机碳最主要的来源。
除了有机质来源的影响外,沉积物TOC含量还受到粒度控制作用[20,26]。与开敞潮滩沉积环境不同,本地区光滩相对较窄,且受到互花米草对粗颗粒物质截留作用,光滩沉积物主要来自于退潮时的滩面细流,因此光滩有着比互花米草滩更细的沉积物富集[27- 28]。但是,0—20cm深度内光滩沉积物TOC含量显著低于互花米草滩。说明粒控效应不是本研究区的主要控制因子,沉积物TOC含量更多的受到互花米草来源有机碳的影响。
2.5 互花米草盐沼有机碳储量变化及固碳能力
由于互花米草各部位生物量和TOC含量的季节动态,单位面积互花米草植被有机碳储量全年变化显著(表2)。其中,10月生物量最大时,有机碳储量达到峰值,为1140g/m2。地下部分4月储量最低,2月储量最高,其他时间变化不明显,且地下储量可达地上部分的1.71—2.74倍。

表2 互花米草不同部位有机碳储量的季节动态/(g/m2)Table 2 Seasonal variation of organic carbon storage in different parts of S. alterniflora
对湿地植物的研究发现,植物体内的氮、磷等元素具有随季节而向上输送蓄积或向下回流的特点[29- 31],本文研究表明,有机碳也存在着类似的运移规律。从春季生长期开始,地上部分有机碳储量增长较快,而地下部分略有下降。4月生长初期,光合作用器官不发达,主要依靠吸收地下根茎中储藏的养分,有机碳转移至茎叶中,导致了4月份地下部分有机碳储量为全年最低。此后,为维系地上部分的快速生长,地下部分生长缓慢或略有下降。10月以后地上储量开始下降,向地下转移积聚,使得地下有机碳储量持续增长,且地上部分减少的储量远大于地下增加的储量。
互花米草盐沼湿地的固碳作用主要体现在两个方面。一方面,湿地植物通过自身生长固定了大量的有机碳。互花米草地下根茎为多年生,年更新率约为0.57[32],用“最大现存法”估算互花米草群落年净初级生产力,依据光合作用反应方程推算,互花米草年固碳能力为2274g m-2a-1,是中国植被平均固碳能力(494 g m-2a-1)的4.60倍[33];另一方面,茂盛的植物可以捕获更多的沉积物,提高了沉积物有机碳埋藏速率[34]。根据沉积物TOC含量和沉积速率,估算出当地盐沼有机碳埋藏速率为470 g m-2a-1,远高于中国内陆盐沼(67g m-2a-1)和沿海滩涂湿地有机碳埋藏速率的平均值(236 g m-2a-1)[35]。本文中,互花米草生物量最大和最小时期有机碳储量的差值为655g/m2,而通过矿化作用埋藏到沉积物中的有机碳为470 g/m2,其余部分有机碳则以不同的形态和枯落物的形式被潮水带走进入沿岸水体[36- 37]。由此可见,互花米草不但在盐沼湿地沉积物有机碳累积中发挥重要作用,对沿岸水体的生态系统也有着重要影响。
3 结论
(1)互花米草的地上生物量、枯落物量以及表层沉积物中的TOC含量有着明显的季节变化。互花米草枯落物衰减实验表明,枯落物的质量在两个月内即减少了44%,有机碳数量衰减了40%。
(2)互花米草滩表层沉积物TOC含量及其中互花米草来源TOC所占比例,均与地表枯落物量之间呈现出相似的季节变化趋势,但存在两个月的“相位差”。这与枯落物分解的时间相吻合,表明枯落物分解释放的有机碳,是影响表层沉积物TOC含量变化的重要因素。
(3)地下生物量随着深度的增加呈指数衰减,超过50%的互花米草地下生物量分布在0—20cm深度内,该深度范围内的沉积物TOC含量较高,且TOC主要来源于互花米草。此外,研究发现垂向TOC含量与地下生物量间存在较好的正相关关系,说明地下生物量对沉积物TOC含量变化有着重要影响。
(4)研究区互花米草年固碳能力为2274g m-2a-1,而互花米草盐沼湿地沉积物中的TOC埋藏速率达到了470 g m-2a-1。除此之外,互花米草盐沼湿地还向近岸水域输出了大量有机碳,对沿岸生态系统产生了重要影响。
致谢:朱庆光、徐夏楠、徐粲、吴丹丹参与野外数据采集工作,赵秧秧、倪文斐、石勇帮助写作,特此致谢。
[1] Brix H, Sorrell B K, Lorenzen B. Are phragmites-dominated wetlands a net source or net sink of greenhouse gases? Aquatic Botany, 2001, 69(2): 313- 324.
[2] 韩冰, 王效科, 欧阳志云. 中国农田生态系统土壤碳库的饱和水平及其固碳潜力. 农村生态环境, 2005, 21(4): 6- 11.
[3] Chmura G L, Anisfeld S C, Cahoon D R, Lynch J C. Global carbon sequestration in tidal, saline wetland soils. Global Biogeochemical Cycles, 2003, 17(4): doi: 10.1029/2002GB001917.
[4] Berg B, McClaugherty C. Plant Litter Decomposition, Humus Formation, Carbon Sequestration. New York: Springer, 2003.
[5] 曹磊, 宋金明, 李学刚, 袁华茂, 李宁, 段丽琴. 滨海盐沼湿地有机碳的沉积与埋藏研究进展. 应用生态学报, 2013, 24(7): 2040- 2048.
[6] Chung C H. Thirty years of ecological engineering withSpartinaplantations in China. Ecological Engineering, 1993, 2(3): 261- 289.
[7] Zhang R S, Shen Y M, Lu L Y, Yan S G, Wang Y H, Li J L, Zhang Z L. Formation ofSpartinaalterniflorasalt marshes on the coast of Jiangsu Province, China. Ecological Engineering, 2004, 23(2): 95- 105.
[8] Chen Z Y, Li B, Zhong Y, Chen J K. Local competitive effects of introducedSpartinaalternifloraonScirpusmariqueterat Dongtan of Chongming Island, the Yangtze River estuary and their potential ecological consequences. Hydrobiologia, 2004, 528(1/3): 99- 106.
[9] Li B, Liao C H, Zhang X D, Chen H L, Wang Q, Chen Z Y, Gan X J, Wu J H, Zhao B, Ma Z J, Cheng X L, Jiang L F, Chen J K.Spartinaalterniflorainvasions in the Yangtze River estuary, China: An overview of current status and ecosystem effects. Ecological Engineering, 2009, 35(4): 511- 520.
[10] Wan S W, Qin P, Liu J, Zhou H X. The positive and negative effects of exoticSpartinaalterniflorain China. Ecological Engineering, 2009, 35(4): 444- 452
[11] Cheng X L, Luo Y Q, Chen J Q, Lin G H, Chen J K, Li B. Short-term C4plantSpartinaalterniflorainvasions change the soil carbon in C3plant-dominated tidal wetlands on a growing estuarine Island. Soil Biology & Biochemistry, 2006, 38(12): 3380- 3386.
[12] Liao C Z, Luo Y Q, Jiang LF, Zhou X H, Wu X W, Fang C M, Chen J K, Li B. Invasion ofSpartinaalternifloraenhanced ecosystem carbon and nitrogen stocks in the Yangtze Estuary, China. Ecosystems, 2007, 10(8): 1351- 1361.
[13] 高建华, 杨桂山, 欧维新. 互花米草引种对苏北潮滩湿地TOC、TN和TP分布的影响. 地理研究, 2007, 26(4): 799-808.
[14] Zhang Y H, Ding W X, Luo J F, Donnison A. Changes in soil organic carbon dynamics in an Eastern Chinese coastal wetland following invasion by a C4 plantSpartinaalterniflora. Soil Biology & Biochemistry, 2010, 42(10): 1712- 1720.
[15] Gao J H, Bai F L, Yang Y, Gao S, Liu Z Y, Li J. Influence ofSpartinacolonization on the supply and accumulation of organic carbon in Tidal salt marshes of Northern Jiangsu province, China. Journal of Coastal Research, 2012, 28(2): 486- 498.
[16] 任美锷. 江苏省海岸带和海涂资源综合调查. 北京: 海洋出版社, 1986.
[17] 钦佩, 马连琨, 谢民, 仲崇信. Fe、Cu、Mn、Zn在互花米草(Spartinaalterniflora)初级生产中的动态研究. 生态学报, 1993, 13(1): 67- 74.
[18] Wider R K, Lang G E. A critique of the analytical methods used in examining decomposition data obtained from litter bags. Ecology, 1982, 63(6): 1636- 1642.
[19] Yang S L, Friedrichs C T, Shi Z, Ding P X, Zhu J, Zhao Q Y. Morphological response of tidal marshes, flats and channels of the outer Yangtze River mouth to a major storm. Estuaries, 2003, 26(6): 1416- 1425.
[20] Andrews J E, Greenaway A M, Dennis P F. Combined carbon isotope and C/N ratios as indicators of source and fate of organic matter in a poorly flushed, tropical estuary: Hunts Bay, Kingston Harbour, Jamaica. Estuarine, Coastal and Shelf Science, 1998, 46(5): 743- 756.
[21] 刘敏, 侯立军, 许世远, 欧冬妮, 蒋海燕, 余婕, Gardner WayneS. 长江口潮滩有机质来源的C、N稳定同位素示踪. 地理学报, 2004, 59(6): 918- 926.
[22] Liu J E, Zhou H X, Qin P, Zhou J. Effects ofSpartinaalterniflorasalt marshes on organic carbon acquisition in intertidal zones of Jiangsu Province, China. Ecological Engineering, 2007, 30(3): 240- 249.
[23] France R, Culbert H, Freeborough C, Peters, R. Leaching and early mass loss of boreal leaves and wood in oligotrophic water. Hydrobiologia, 1997, 345(2/3): 209- 214.
[24] Wang X C, Chen R F, Berry A. Sources and preservation of organic matter in Plum Island salt marsh sediments (MA, USA): long-chainn-alkanes and stable carbon isotope compositions. Estuarine, Coastal and Shelf Science, 2003, 58(4): 917- 928.
[25] Cifuentes L A, Coffins R B, Solorzano L, Cardenas W, Espinoza J, Twilley R R. Isotopic and elemental variations of carbon and nitrogen in a mangrove estuary. Estuarine, Coastal and Shelf Science, 1996, 43(6): 781- 800.
[26] Mayer L M. Surface area control of organic carbon accumulation in continental shelf sediments. Geochimica et Cosmochimica Acta, 1994, 58(4): 1271- 1284.
[27] 王爱军, 汪亚平, 杨旸. 江苏王港潮间带表层沉积物特征及输运趋势. 沉积学报, 2004, 22(1): 124- 129.
[28] Wang Y P, Gao S, Jia J J, Thompson C E L, Gao J H, Yang Y. Sediment transport over an accretional intertidal flat with influences of reclamation, Jiangsu coast, China. Marine Geology, 2012, 291- 294: 147- 161.
[29] Quan W M, Han J D, Shen A L, Ping X Y, Qian P L, Li C J, Shi L Y, Chen Y Q. Uptake and distribution of N, P and heavy metals in three dominant salt marsh macrophytes from Yangtze River estuary, China. Marine Environmental Research, 2007, 64(1): 21- 37.
[30] Neves J P, Ferreira L F, Simões M P, Gazarini L C. Primary production and nutrient content in two salt marsh species,AtriplexportulacoidesL. andLimoniastrummonopetalumL., in Southern Portugal. Estuaries and Coasts, 2007, 30(3): 459- 468.
[31] 郭长城, 胡洪营, 李锋民, 下橋雅樹, 迫田章義. 湿地植物香蒲体内氮、磷含量的季节变化及适宜收割期. 生态环境学报, 2009, 18(3): 1020- 1025.
[32] Schubauer J P, Hopkinson C S. Above- and belowground emergent macrophyte production and turnover in a coastal marsh ecosystem, Georgia. Limnology and Oceanography, 1984, 29(5): 1052- 1065.
[33] 何浩, 潘耀忠, 朱文泉, 刘旭拢, 张晴, 朱秀芳. 中国陆地生态系统服务价值测量. 应用生态学报, 2005, 16(6): 1122- 1127.
[34] Mayer L M, Macko S A, Cammen L. Provenance, concentrations and nature of sedimentary organic nitrogen in the Gulf of Maine. Marine Chemistry, 1988, 25(3): 291- 304.
[35] 段晓男, 王效科, 逯非, 欧阳志云. 中国湿地生态系统固碳现状和潜力. 生态学报, 2008, 28(2): 463- 469.
[36] Odum E P. The status of three ecosystem-level hypotheses regarding salt marsh estuaries: tidal subsidy, outwelling, and detritus-based food chains // Estuarine Perspectives. New York: Academic Press, 1984: 485- 495.
[37] Childers D L, Day J W Jr, McKellar H N Jr. Twenty more years of marsh and estuarine flux studies: revisiting Nixon (1980) // Concepts and Controversies in Tidal Marsh Ecology. Netherlands: Springer, 2000: 391- 423.
alternifloramarsh
FENG Zhenxing1, GAO Jianhua1,2,*, CHEN Lian1, WANG Yaping1, GAO Jianhui3, BAI Fenglong4
1KeyLaboratoryofCoastandIslandDevelopment,MinistryofEducation,NanjingUniversity,Nanjing210093,China2KeyLaboratoryofMarineSedimentologyandEnvironmentalGeology,StateOceanicAdministration,Qingdao266061,China3SchoolofEnvironmentalScienceandEngineering,YanchengInstituteofTechnology,Yancheng224051,China4TheKeyLaboratoryofMarineHydrocarbonResourcesandEnvironmentalGeology,MinistryofLandandResourcesQingdaoInstituteofMarineGeology,Qingdao266071,China
Spartinaalterniflora(C4) was introduced to the coastal region of Jiangsu Province as an exotic species in 1982. This perennial grass then gradually invaded regions previously occupied by native C3plants. Much research has focused on variation of the ecosystem carbon cycle and has demonstrated that thisS.alterniflorainvasion has increased the carbon accumulation rate. However, the impact of plant biomass (both aboveground and belowground) on the organic carbon content of salt marsh sediments requires further study. To explore the response of organic carbon content to the biomass dynamics in anS.alternifloramarsh, the contents of total organic carbon (TOC), total nitrogen (TN), and stable carbon isotope composition (δ13C) of sediments, in anS.alternifloramarsh as well as the bare flat, were investigated by analysis of coral and surface sediments, together with the biomass variations ofS.alterniflora. The sediment water content, grain size, andδ13C of coastal water andS.alterniflorawere also determined. The Litterbag method was used to measure theS.alternifloralitter decomposition rate. The results indicate that the organic carbon of newer litter rapidly decomposed in the first 2 months, and then remained stable. The seasonal variation of the TOC content of surface sediments in theS.alternifloramarsh was significant, and a 2-month phase shift was not only observed in the TOC content variations and surface litter mass but was also recorded for the proportion of organic carbon derived fromS.alternifloraand surface litter mass. However, this phase-shift phenomenon did not exist on the bare flat, indicating that the litter is a dominant source of the organic carbon in surface sediments. The results show that 60% and 33% of belowground biomass was observed in the top 20 cm and at 20—40 cm depths, respectively. Compared with the bare flat, the vertical changes of TOC content in theS.alternifloramarsh showed a different distribution pattern. That is to say, the TOC content of sediments at depths < 20 cm was significantly higher than those at depths > 20 cm, and had an increasing trend from depth 20 cm to the surface. A similar variation was observed for the proportion of organic carbon derived fromS.alterniflorain the top 20 cm of sediments. Furthermore, there was a remarkably positive relationship between the belowground biomass ofS.alternifloraand TOC content. This suggests that belowground biomass plays an important role in TOC variation along the profiles. The greatest and least plant organic carbon storages in theS.alternifloramarsh were observed in October and April, respectively. The average carbon fixation ofS.alterniflora, with quantity 2274 g·m-2·a-1, was 460% that of terrestrial vegetation in China. Similarly, the burial rate of organic carbon ofS.alternifloramarsh sediments, with quantity 470 g·m-2·a-1, was significantly greater than the average value of coastal salt marshes in China. Thus, theS.alterniflorasalt marsh fixed large amounts of carbon dioxide from the air, and enhanced the organic carbon accumulation of the sediment.
organic carbon;Spartinaalterniflora; biomass; dynamics
江苏省自然科学基金重点研究专项(BK2011012);海洋公益性行业科研专项经费项目(2010418006);海洋沉积与环境地质国家海洋局重点实验室开放基金(MASEG201205);国土资源部海洋油气资源和环境地质重点实验室基金项目(MRE201408)
2014- 04- 09;
2014- 11- 21
10.5846/stxb201404090685
*通讯作者Corresponding author.E-mail: jhgao@nju.edu.cn
冯振兴,高建华,陈莲,汪亚平,高建慧,白凤龙.互花米草生物量变化对盐沼沉积物有机碳的影响.生态学报,2015,35(7):2038- 2047.
Feng Z X, Gao J H, Chen L, Wang Y P, Gao J H, Bai F L.The response of organic carbon content to biomass dynamics inSpartinaalternifloramarsh.Acta Ecologica Sinica,2015,35(7):2038- 2047.
