半干旱黄土丘陵区纯林土壤腐殖质异化特征及与其他性质的关系
2015-03-11张晓曦刘增文朱博超邴塬皓LucNhuTrung
张晓曦,刘增文,朱博超,邴塬皓,于 齐,Luc Nhu Trung,5
1 西北农林科技大学水土保持研究所, 杨凌 712100 2 西北农林科技大学资源环境学院, 杨凌 712100 3 农业部西北植物营养与农业环境重点实验室, 杨凌 712100 4 西北农林科技大学林学院, 杨凌 712100 5 农业与农村发展厅, 越南老街 330100
半干旱黄土丘陵区纯林土壤腐殖质异化特征及与其他性质的关系
张晓曦1,刘增文2,3,朱博超2,邴塬皓4,于 齐4,Luc Nhu Trung1,5
1 西北农林科技大学水土保持研究所, 杨凌 712100 2 西北农林科技大学资源环境学院, 杨凌 712100 3 农业部西北植物营养与农业环境重点实验室, 杨凌 712100 4 西北农林科技大学林学院, 杨凌 712100 5 农业与农村发展厅, 越南老街 330100
纯林长期生长或多代连栽必然会导致土壤腐殖质含量和构成发生异化,探究这种异化特征及其与土壤其他性质的关系可以为纯林管理或混交改造提供科学依据。通过对半干旱黄土丘陵区南泥湾林场8种典型纯林土壤腐殖质及其他性质进行系统检测,结果表明:(1)侧柏林土壤腐殖质含量最高(34.61 g/kg),腐殖化程度和稳定性一般;白榆和白桦林土壤的腐殖质含量中等(19.69—23.58 g/kg)、腐殖化程度和稳定性最佳;茶条槭和小叶杨林土壤的腐殖质含量(20.59—22.53 g/kg)和构成均为中等水平;油松、沙棘和刺槐林土壤的腐殖质质量较低(11.77—13.81 g/kg),且腐殖化程度较低,稳定性相对最差;(2)与胡敏酸含量存在显著相互促进作用(P<0.05)的土壤性质为CEC、N、微生物量和蛋白酶活性(相关系数0.769—0.926,下同),存在显著相互抑制作用的为有效Cu(-0.793);与富啡酸存在显著相互促进作用的为N、CEC、微生物量、蔗糖酶和磷酸酶活性(0.836—0.955),存在显著相互抑制作用的为有效Cu(-0.822);与胡敏素存在显著相互促进作用的为N、CEC、微生物量、磷酸酶活性和有效Zn(0.766—0.951),存在显著相互抑制作用的为脱氢酶活性(-0.784)。(3)腐殖质构成与其他性质的相关性均不显著(P>0.05),其中相对有利于提高胡敏酸/腐殖酸含量之比的土壤性质为蛋白酶、蔗糖酶和过氧化氢酶活性,而不利的是脱氢酶活性;相对有利于提高胡敏酸/富啡酸含量之比的为速效K、CEC和脲酶活性,而不利的是脱氢酶活性。(4)总体而言土壤腐殖质含量较之腐殖质构成与其他性质之间具有更大的相关性;向土壤增施N肥可以促进腐殖质的形成,增加K肥则有利于腐殖质构成的改善,而通过混交改造或增加林下植被是促进纯林土壤腐殖质化过程和解决土壤退化的根本措施。
纯林; 腐殖质; 异化特征; 土壤性质
腐殖质含量和构成是反映土壤肥力质量的重要指标,所以历来受到土壤和生态学研究的重视[1]。由单一树种组成的纯林,由于受到枯落物分解、根系分泌物、养分循环和群落环境的特殊影响,必然导致其土壤腐殖质在含量和构成上发生明显的异化(即特异变化)并对森林土壤生态过程产生深远的影响。在陕北半干旱黄土丘陵区分布的森林70%以上为纯林,已有调查研究表明,由于立地环境条件相对恶劣,土壤腐殖质化过程缓慢,这些纯林的长期经营和多代连栽出现了生长衰退、土壤极化或退化等现象[2]。分析出现这些问题的原因,是否与纯林土壤腐殖质异化特征有关?森林土壤腐殖质含量和构成特征与其他理化生物学性质之间究竟有何关系?哪些土壤性质能够促进(抑制)腐殖质含量的提高和改善(恶化)腐殖质的构成?所以,搞清楚不同树种纯林土壤的腐殖质分异特征,综合分析土壤腐殖质与其他性质之间关系对解决纯林衰退问题非常必要。
截至目前,国内外关于森林土壤腐殖质的研究已经广泛涉及到腐殖质特性与土壤肥力的关系,森林发育阶段、经营方式和年限对土壤腐殖质影响,以及森林土壤腐殖质对土壤抗蚀性的影响等诸多方面[3-5]。然而,针对半干旱黄土丘陵区不同树种纯林开展土壤腐殖质特征的专门系统研究相对较少。为此,本研究以半干旱黄土丘陵区南泥湾林场常见典型纯林类型为对象,通过对土壤腐殖质及其他性质进行系统检测,分析不同树种纯林土壤腐殖质的异化特征,并对其与其他土壤性质之间的关系进行相关统计分析,旨在分析纯林生长对土壤腐殖质的影响以及由此可能导致的土壤性质的变化,据此提出改善纯林土壤性质的途径和方法,为后期纯林管理或混交改造提供科学依据。
1 材料与方法
1.1 标准地设置、土壤样品采集
以地处黄土高原中部暖温带半干旱丘陵区的延安南泥湾林场为对象,当地海拔900—1 400 m,年均气温8.5—9.5 ℃,无霜期165—190 d,降水量约520—550 mm。森林植被以人工林为主,也有少量天然次生林,土壤类型为黄绵土。本次调查选择了地貌部位(梁峁坡中部)、坡向(阴坡或半阴坡)和坡度(15—25°)等立地条件基本相似、郁闭度大于0.85、未经人为干扰的当地具有典型代表性的8个不同树种成熟纯林(表1)。林下植被主要以羊胡子草Carexrigescens、冰草Agropyroncristatum和蒿类Aremisia等草本植物为主,少有个别的苦糖果Lonicerastandishii、牛奶子Elaeagnusumbellate等灌木,且总盖度均小于15%。
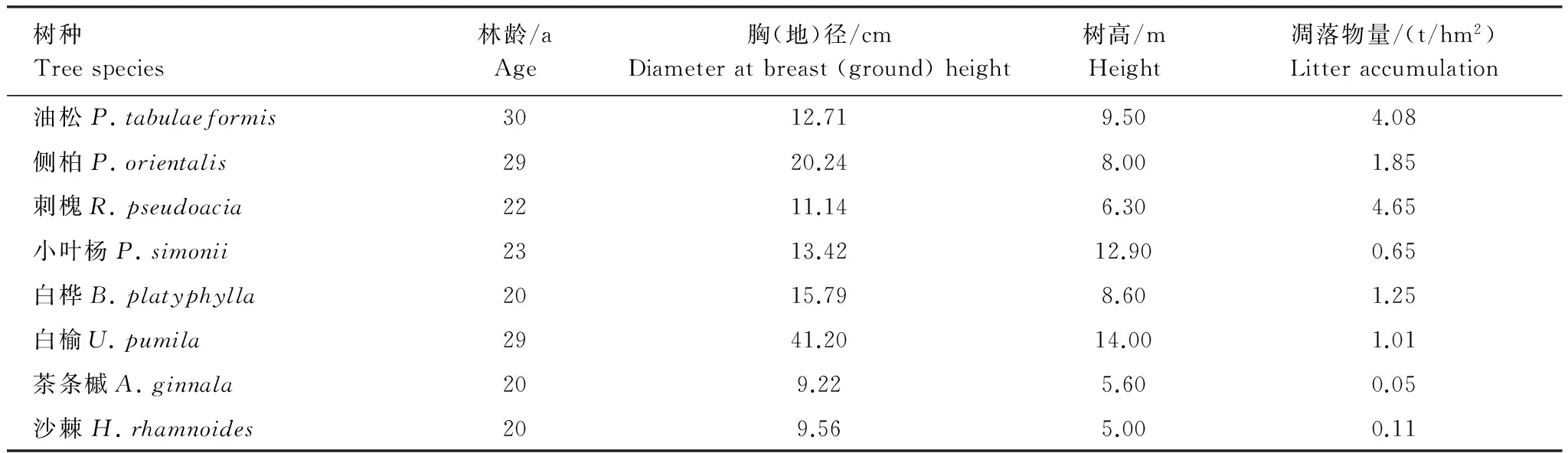
表1 土壤采集地林分平均生长状况Table 1 Introduction of forests for soil collection
具体采样方法为:首先分别每个树种纯林建立3块20 m×20 m的标准地(共计8个树种24块标准地),然后在每个标准地内均匀设置5个1 m×1 m的小样方,以二次混合采样法(即先每个样方混合采样,然后5个样方再次等量混合采样)采集0—20 cm土层的土壤样品约3 kg左右(即每个标准地采集1份二次混合样,每种林地土样3份,共计24份样品),剔除植物根系、未分解枯落物和砾石等杂物后带回室内备测。
1.2 土壤性质测定与数据处理
土壤pH值采用PHS-2型酸度计测定(水土比为2.5∶1);腐殖质含量(胡敏酸、富啡酸和胡敏素)采用焦磷酸钠浸提-重铬酸钾氧化法测定;土壤全N采用半微量凯氏定N法测定;碱解N采用扩散法测定;有效P 采用NaHCO3法测定;速效K采用醋酸铵浸提火焰光度法测定;CEC采用乙酸钠-乙酸铵火焰光度法测定;Cu、Zn、Fe、Mn的全量采用硝酸高氯酸消煮-原子吸收分光光度法(AAS法)测定,有效态含量采用为二乙三胺五乙酸(DTPA)浸提-原子吸收分光光度法(AAS法)测定[6]。微生物量C采用氯仿熏蒸-K2SO4浸提法测定;脲酶活性采用苯酚钠-次氯酸钠比色法测定;蔗糖酶活性采用Na2S2O3滴定法测定;过氧化氢酶活性采用KMnO4滴定法测定;过氧化物酶活性采用滴定法测定;脱氢酶采用三苯基四唑氯化物比色法测定;磷酸酶活性采用磷酸苯二钠比色法测定;蛋白酶活性采用茚三酮比色法测定[7-8]。
测定数据应用Excel 2010进行整理,每个树种林地的以上所有指标测定均采用3块标准地(即3次重复)实测的平均值(每块标准地采用误差不超过5%的3个测次平均),每个林地的每项测定值使用SPSS 19.0软件进行单因素方差分析,多重比较采用最小显著差法(LSD)(α=0.05)。
2 结果与分析
2.1 不同纯林土壤腐殖质异化特征比较
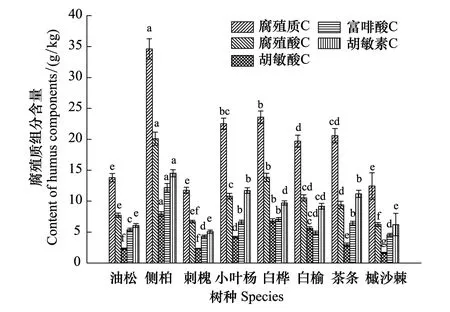
图1 土壤腐殖质含量特征Fig.1 Properties of soil humus contents
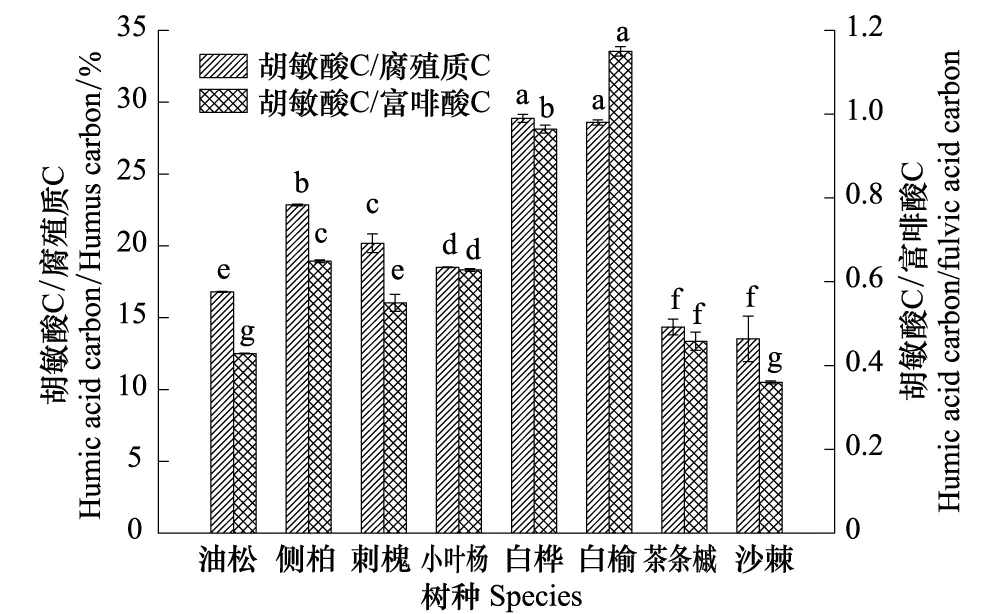
图2 土壤腐殖质构成特征Fig.2 Properties of soil humus composition
根据对8种纯林土壤腐殖质特征测定的结果表明(图1),总腐殖质含量以侧柏林土壤显著最高(34.61 g/kg),其次为白桦和小叶杨(22.53—23.58 g/kg),再次为白榆和茶条槭(19.70—20.59 g/kg),油松、沙棘和刺槐林显著最低(11.77—13.81 g/kg)。总腐殖酸含量以侧柏林土壤显著最高(20.10 g/kg),其次为白榆、小叶杨、白桦和茶条槭(9.41—13.87 g/kg),油松、刺槐和沙棘显著最低(6.23—7.73 g/kg)。其中,胡敏酸含量以侧柏林地土壤显著最高(7.91 g/kg),其次为白榆、白桦和小叶杨(4.17—6.81 g/kg),茶条槭、刺槐、油松和沙棘较低(1.65—2.96 g/kg);富啡酸含量以侧柏林地土壤显著最高(12.19 g/kg),其次为白榆、小叶杨和茶条槭(6.45—7.06 g/kg),再次为油松和白桦(4.90—5.41 g/kg)、沙棘和刺槐显著最低(4.32—4.58 g/kg);胡敏素含量以侧柏林地土壤显著最高(14.52 g/kg),其次为小叶杨和茶条槭(11.18—11.72 g/kg),再次为白榆和白桦(9.17—9.71 g/kg),沙棘、油松和刺槐显著最低(5.07—6.22 g/kg)。
腐殖质的构成以胡敏酸在腐殖质中所占比例以及胡敏酸与富啡酸比值来反映(图2)。结果表明,胡敏酸/腐殖质比例以白榆和白桦林地土壤显著最高(28.59%—28.88%),其次为侧柏、刺槐、小叶杨和油松(16.80%—22.85%),茶条槭和沙棘显著最低(13.26%—14.35%);胡敏酸/富啡酸比值以白榆显著最高(1.15),其后依次为白桦、侧柏、小叶杨和刺槐(0.55—0.95),再次为茶条槭和油松(0.43—0.46),沙棘显著最低(0.36),所有林地土壤在胡敏酸/富啡酸比值这一指标上均存在显著差异。
综合以上分析可知,8种纯林土壤比较,侧柏林土壤腐殖质含量最高但腐殖化程度和稳定性一般;白榆和白桦林土壤的腐殖质含量中等、腐殖化程度最高,稳定性最好;茶条槭和小叶杨林土壤的腐殖质含量和构成均为中等水平;油松、刺槐和沙棘林土壤的腐殖质质量较低且腐殖化程度低、稳定性差。
2.2 森林土壤腐殖质特征与微生物及土壤酶活性的关系分析
土壤腐殖质的形成是各种有机物质在微生物活动和一系列酶促反应下逐步降解和转化的结果,同时腐殖质化过程又会对微生物和土壤酶活性产生反馈作用。但是,针对半干旱黄土丘陵区森林土壤来说,土壤腐殖质含量及构成与微生物量和土壤酶活性之间的相互关系和相互作用强度究竟如何?现以24块标准地的数据(表2)进行统计相关分析(表3),结果表明:与胡敏酸含量呈显著正相关的指标为微生物量C和蛋白酶;与富啡酸含量呈显著正相关作用的指标为微生物量C、磷酸酶和蔗糖酶;与胡敏素含量呈显著正相关的指标为:微生物量C和磷酸酶,呈显著负相关的为脱氢酶;可见,微生物、蛋白酶、磷酸酶和蔗糖酶与腐殖质的形成具有显著的相互促进作用,脱氢酶则表现出一定的相互抑制作用。
微生物量和酶活性与腐殖质构成的相关性均未达到显著水平,相较之下较为有利于提高胡敏酸/腐殖质比值和胡敏酸/富啡酸比值的指标为脲酶和蛋白酶,而较为不利的指标为脱氢酶。可见,酶活性对腐殖质构成的影响不明显,蛋白酶和脲酶相对最有利于腐殖质良好结构的形成,而脱氢酶最为不利。

表2 不同林地土壤生物学性质Table 2 Biological properties of soil from different forests
表中数值表示平均值(标准差); 同列中不同字母代表具有显著性差异
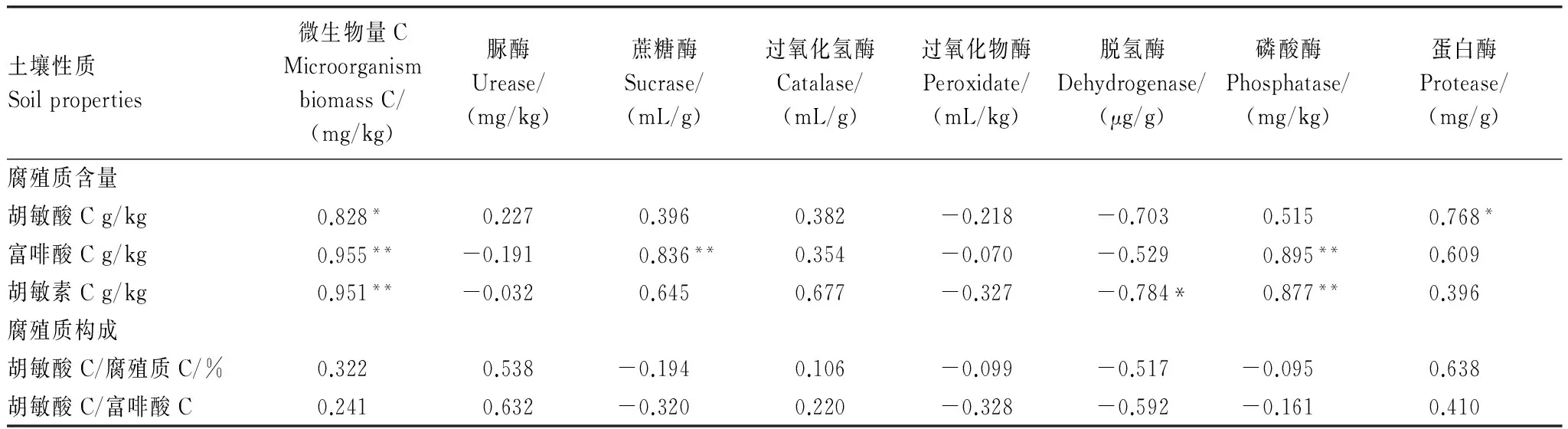
表3 森林土壤腐殖质特征与其他生物学性质的相关系数(n=24)Table 3 Correlation coefficients of soil humus with other biological properties
腐殖质含量 Humus contents(胡敏酸 Humic acid;富啡酸 Fulvic acid;胡敏素 Humin;腐殖酸 Humic acids;腐殖质 Humus);腐殖质构成Humus composition(胡敏酸C/腐殖质C Humic acid carbon/Humus carbon;胡敏酸C/富啡酸C Humic acid carbon/Fulvic acid carbon); *P<0.05,**P<0.01
2.3 森林土壤腐殖质特征与化学性质的关系分析
土壤腐殖质含量及构成特征同样与其他化学性质密切相关。为了探明半干旱黄土丘陵区的森林土壤腐殖质特征与其他化学性质(表4)之间的相互关系及其强度,做统计相关分析(表5)。
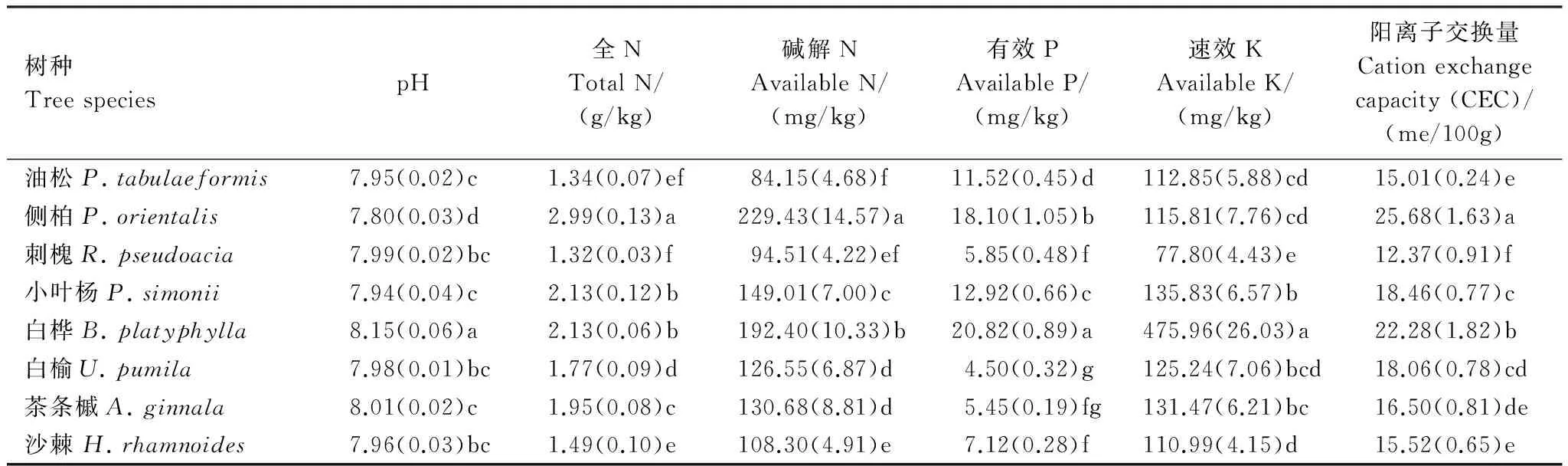
表4 不同林地土壤化学性质Table 4 Chemical properties of soil from different forests
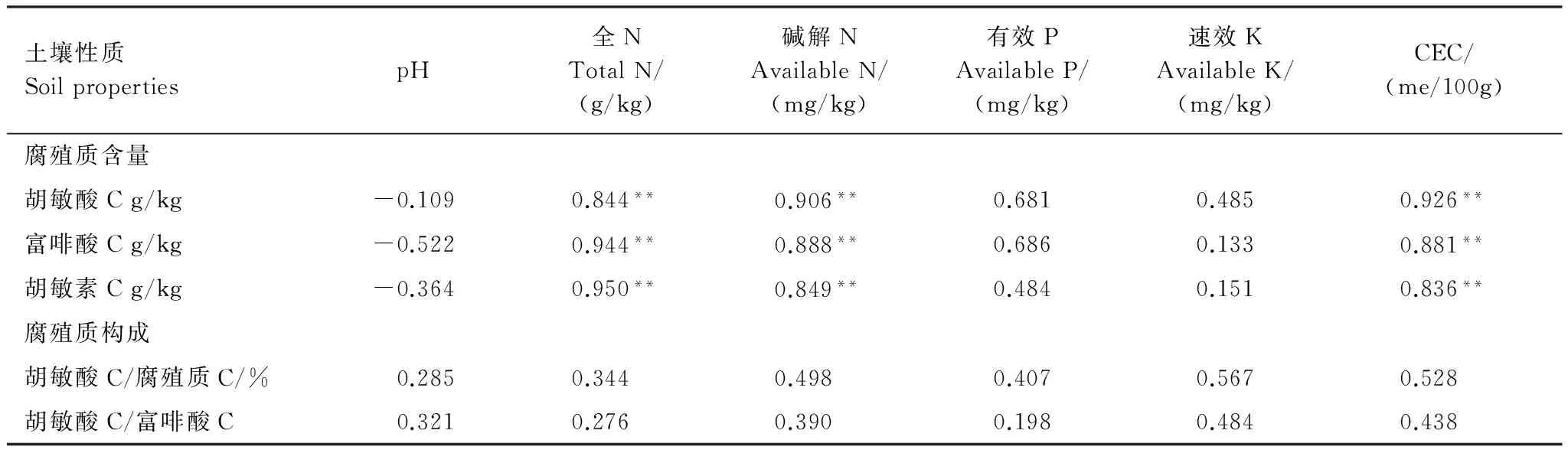
表5 森林土壤腐殖质特征与其他化学性质的相关系数Table 5 Correlation coefficients of soil humus with other chemical properties
与胡敏酸、富啡酸和胡敏酸含量均呈显著正相关作用的指标为CEC、碱解N和全N;pH值与3种腐殖质组分的含量均表现出不显著的负相关关系;可见,腐殖质各组分的形成与土壤N素含量存在显著相互促进作用,能显著的提高土壤的保肥能力,但有导致土壤出现酸化的趋势。
土壤化学性质与腐殖质构成的相关性均未达到显著水平,相较之下较为有利于明显有利于提高胡敏酸/腐殖质比值和提高胡敏酸/富啡酸比值的指标依次为速效K、CEC和碱解N;良好的腐殖质构成与土壤pH值则具有一定的正相关关系。可见,腐殖质构成与速效养分仅存在一定的相互促进作用,而较好的腐殖质构成可能部分抵消其含量增加导致的土壤酸化趋势。
2.4 森林土壤腐殖质特征与微量元素的关系分析
根据腐殖质含量及构成与土壤微量元素含量(表6)的统计相关分析表明(表7):
与胡敏酸和富啡酸含量呈显著负相关的指标为有效Cu,与胡敏素含量呈显著正相关的指标为有效Zn。此外,有效Cu含量与胡敏素含量与也呈现出一定的负相关,而有效Zn含量则与胡敏酸和富啡酸含量呈一定正相关关系。由此表明,腐殖质能提高土壤Zn的有效性而降低Cu的有效性。
微量元素与腐殖质构成特征的关系均未达显著水平,其中较为有利于提高胡敏酸/腐殖质比值的指标为有效Zn,而较为不利的依次为全Fe、有效Mn和有效Cu。较为有利于提高胡敏酸/富啡酸比值的指标为有效Zn,而较为不利的指标主要为有效Mn、全Fe和有效Cu。表明土壤中Zn的适当积累以及Zn有效性的提高可能有利于腐殖质腐殖化程度的提高和稳定性的增加,而Fe的积累和Cu、Mn有效性的提高则相反。
微量元素含量较之大量元素含量和生物学性质与腐殖质各组分含量及构成特征的相互作用更为微弱,均未达到显著水平,且相关的正负性相对复杂,这可能与不同微量元素本身特殊的化学性质有关。
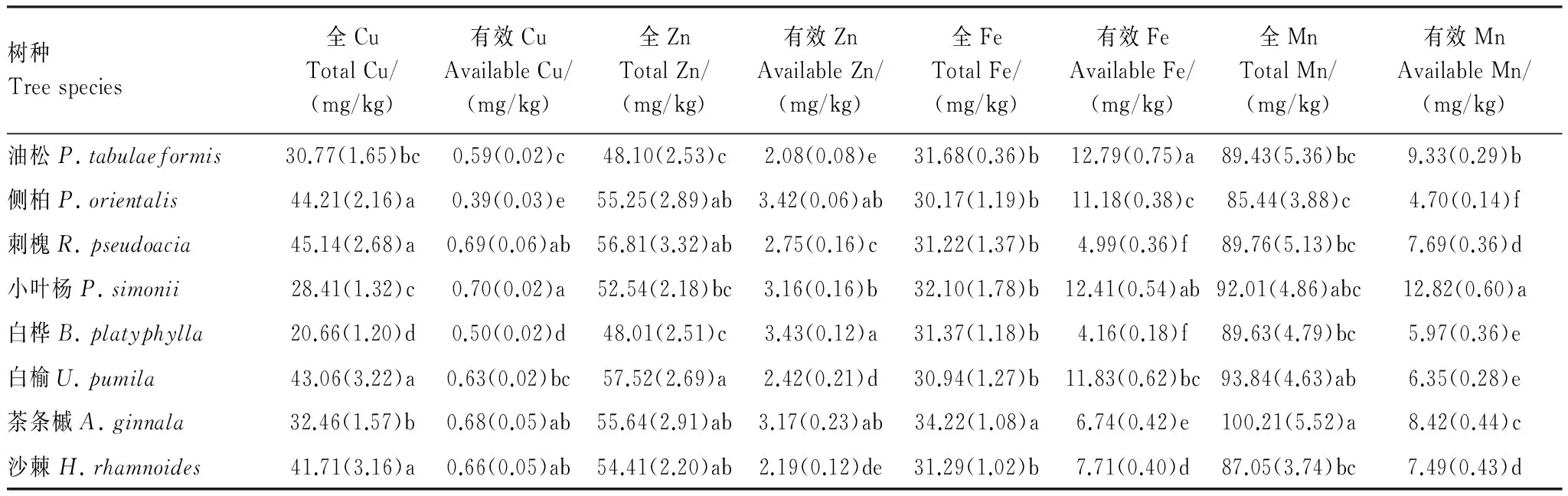
表6 不同林地微量元素含量Table 6 Microelements content of soil from different forests
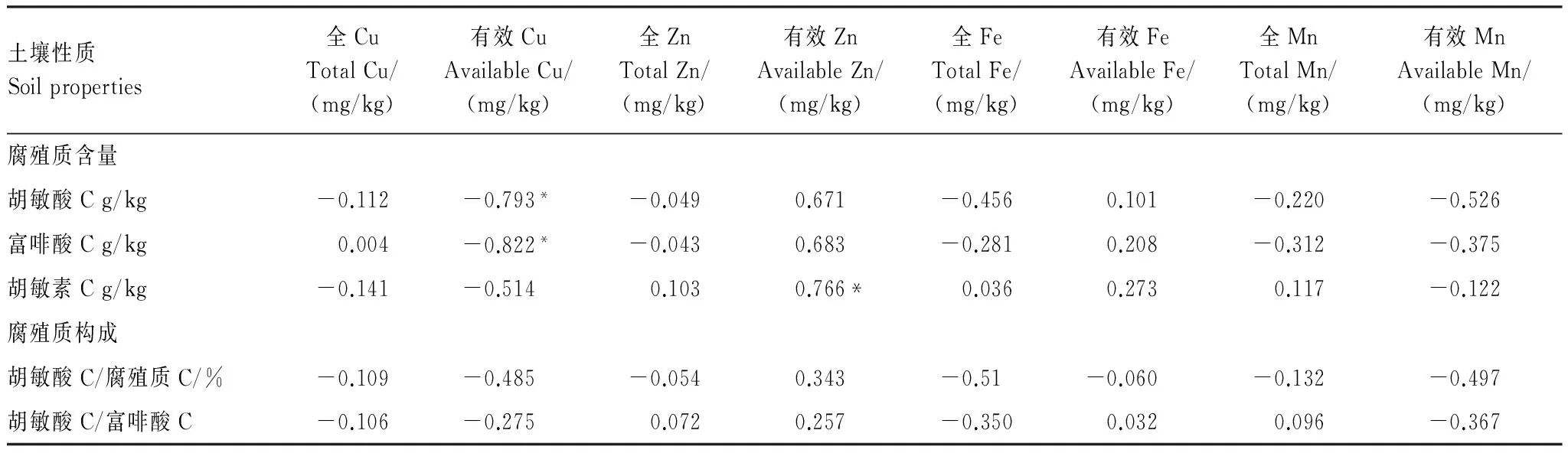
表7 森林土壤腐殖质性质与微量元素含量的相关系数Table 7 Correlation coefficients among soil humus properties and microelements content
3 讨论
3.1 关于不同纯林土壤的腐殖质含量及构成特征
胡敏酸是土壤腐殖质最重要的组成成分,由于其具有大量边缘官能团和活跃的化学性质,能与土壤养分和矿物产生作用,所以常用来表征土壤腐殖质的品质。土壤的腐殖化程度一般采用胡敏酸/富啡酸的比值表示,可反映腐殖质的芳构化和聚合程度以及稳定性[9]。然而,也有一些研究表明,由于受到计算方法的限制可能导致胡敏酸/富啡酸比值的误差较大,所以认为胡敏酸/总腐殖质比值更适于作为腐殖化程度的表征指标[10]。考虑到所研究土壤的差异和为了更加全面、客观,本研究选取了上述两种指标并结合腐殖质各组分的含量等来综合考察8种纯林土壤的腐殖质特征,结果表明不同树种林地土壤的腐殖质含量和构成均存在明显的异化现象,这主要是不同树种林木枯落物的性质差异所致[11]。Trap等[12]对枯落物基质质量与腐殖质形式之间的关系进行分析,证明枯落物基质质量是影响森林熟化过程中腐殖质形成和形式变化的主要因子,并指出枯落物的C/N、木质素/N比值和N、K的含量均对腐殖质形式的变化存在显著影响。本文与Trap等的研究结论相似:白榆和白桦等枯落物含N高或C/N较低的阔叶乔木树种(具体数值见参考文献[13])形成的腐殖质质量也较好,而枯落物基质质量较差的油松分解过程受到限制,转化的腐殖质也就相对较少。这是由于木质素是腐殖质形成的重要碳源,其分解对腐殖质形成具有重要的意义[14-15],而基质质量差的枯落物含N量相对低,不能满足微生物(尤其是在木质素分解过程中具有重要地位的真菌)的生活需要[16],从而影响到腐殖质的累积量和构成优劣。同时,本研究的结果表明,枯落物某些微量元素含量的差异也对腐殖质的形成和组成存在一定影响,这可能是由于这些微量元素参与了某些酶类的形成或者是其发挥活性的关键因子。例如本研究发现Mn的有效性提高不利于腐殖质的形成,与Trap等[12]的结论相似。这是由于Mn是Mn-过氧化物酶的重要组成部分,而后者直接影响了木质素的分解[17],从而影响到腐殖化过程。沙棘的枯落物数量较少,即使具有较低的C/N比和相对高N含量,其形成的腐殖质数量也较少。
总之,腐殖质形成的数量和构成受到枯落物类型、数量和质量的制约,其中枯落物基质质量的作用最为重要。当然,其他限制因子如树种特殊化学成分(如多酚等影响分解的物质)[18]、微量元素(如Mn)、叶生理结构(如角质层、蜡质厚度等)[19]等均可能对腐殖质形成过程产生影响,其规律有待更为深入的探究。
3.2 关于腐殖质特征与其他土壤性质的关系
林地土壤腐殖质的各组分含量均与全N、碱解N和CEC存在紧密的正相关关系,与党亚爱和Fontana等[20-21]研究结果相同。表明土壤N含量与枯落物基质质量共同决定了土壤微生物的养分供应,进而影响到其对腐殖质合成和分解的方向与速率。一些研究表明[22],P的供应也会限制土壤微生物对有机质的利用与转化,故土壤中P的含量也可能会对腐殖质的形成和积累产生一定影响,本研究的结果支持了上述观点。胡敏酸和富啡酸分别呈弱酸性和强酸性,其含量是影响土壤pH的重要因子,本研究的结果表明,腐殖酸含量总体与土壤酸碱性表现为相对明显的负相关,这与Andersson等[23]的结果相似,与张林海等[24]的研究结果相反,证明pH对腐殖质的各组分存在一定的反作用,影响其积累与分解的同时也影响其化学性质的变化[25-26]。腐殖质组分含量与微量元素含量的相关性方向多变,这与何蓉等[4]的研究结果相似,其原因则尚需进一步的研究加以解释。
前人研究发现,在林分发育过程中微生物数量会出现先升后降的现象,与此同时,随森林经营时间延长和林分更新,森林土壤腐殖质的质量会有所下降[2]。本研究表明,微生物量C与腐殖质组分间存在紧密的正相关,而与腐殖质构成指标间存在较弱正相关,表明腐殖质(含量和构成)会随微生物量的变化做出响应,为上述发现提供了一定的机理解释。
本研究结果发现蔗糖酶、蛋白酶和磷酸酶活性与腐殖质某些组分呈显著正相关关系,这与闫德仁[27]的结果相反。而脱氢酶与胡敏素显著负相关、过氧化物酶与腐殖质组分含量呈现一定的正相关,与闫德仁的结果相同。表明前三者的活性提高可能提供了腐殖质形成的一些前体物质(如加速蛋白质水解提供氨基酸)或者提供了有机质分解的养分条件(如磷酸酶为微生物提供P),且蔗糖等低分子物质确实参与了腐殖质的形成过程,这与窦森等的总结相似[16]。另有研究认为,酶在土壤中以酶-腐殖质复合体的形式存在[28],从而保持其活性,但针对不同酶类的研究结果存在差异,其机理有待进一步深入探讨和验证。
不同有效态微量元素与腐殖质的关系表现出较大差异,这可能与特定元素与腐殖物质的络合物稳定程度有关。本研究结果表明,有效Cu与腐殖酸含量呈现显著负相关,有效Zn则与胡敏素含量呈显著正相关。这与吴炳孙等[29]的部分研究结果不符。其原因可能在于研究土壤的腐殖质含量和结构有所差异,且腐殖质各组分含量不同情况下,对微量元素起主要“活化”或者“钝化”作用的腐殖质组分也有所变化[29]。此外,在高离子强度下,被土壤矿质吸附的腐殖质将因静电作用被压缩,其分子结构发生变化[30]。这种作用也会影响微量元素及其他土壤性质与腐殖质特性之间的关系。
与多数研究不同,本研究中腐殖质构成指标与土壤性质的相关性总体上弱于腐殖质含量与相应土壤性质的相关性,证明在研究区土壤条件下,与土壤理化生物学性质关系更紧密的主要为腐殖质各组分的绝对含量而非其组成结构。此外,从本研究结果对生产实践的指导意义来看,向土壤增施N肥可以促进腐殖质的形成,增加K肥则有利于腐殖质构成的改善,而通过选择适宜树种混交以调节枯落物腐解和养分释放过程[18,31],从而改善归还土壤的有机质结构[32],或者通过增加林下植被以提高森林土壤的腐殖质含量可能是解决研究区纯林土壤退化现象的根本措施之一。
4 结论
(1)侧柏林土壤腐殖质含量最高,但腐殖化程度和稳定性一般;白榆和白桦林土壤的腐殖质含量中等、腐殖化程度和稳定性最佳;茶条槭和小叶杨林土壤的腐殖质含量和构成均为中等水平;油松、刺槐和沙棘林土壤的腐殖质质量较低,且腐殖化程度较低,稳定性相对最差。
(2)与胡敏酸含量存在显著相互促进作用的土壤性质为CEC、N(全N和碱解N)、微生物量和蛋白酶活性,存在显著相互抑制作用的为有效Cu;与富啡酸存在显著相互促进作用的为N、CEC、微生物量、蔗糖酶和磷酸酶活性,存在显著相互抑制作用的为有效Cu;与胡敏素存在显著相互促进作用的为N、CEC、微生物量、磷酸酶活性和有效Zn,存在显著相互抑制作用的为脱氢酶活性。
(3)腐殖质构成与其他性质的相关性均不显著,其中,相对有利于提高胡敏酸/腐殖酸含量之比的土壤性质为蛋白酶、蔗糖酶和过氧化氢酶活性;相对有利于提高胡敏酸/富啡酸含量之比的为速效K、CEC和脲酶活性。脱氢酶活性的提高最不利于腐殖质良好构成的形成。
(4)总体而言,土壤腐殖质含量较之腐殖质构成与其他性质之间具有更大的相关性,在实践中向土壤增施N肥可以促进腐殖质的形成,增加K肥则有利于腐殖质构成的改善,而通过混交改造或增加林下植被是促进纯林土壤腐殖质化过程和解决土壤退化的根本措施。
[1] 窦森, 肖彦春, 张晋京. 土壤胡敏素各组分数量及结构特征初步研究. 土壤学报, 2006, 43(6): 934-940.
[2] 刘增文, 段而军, 刘卓玛姐, 冯顺煜. 黄土高原半干旱丘陵区不同树种纯林土壤性质极化研究. 土壤学报, 2009, 46(6): 1110-1120.
[3] 赵兰坡, 王杰, 刘景双, 刘淑霞, 王艳玲, 王鸿斌, 张志丹. 不同肥力条件下黑土及其有机无机复合体的腐殖质组成. 应用生态学报, 2005, 16(1): 93-99.
[4] 何蓉, 张培芬, 杨卫. 高黎贡山自然保护区土壤腐殖质组成与微量元素含量的研究. 西部林业科学, 2006, 35(2): 48-52.
[5] 王鑫, 刘建新. 黄土高原沟壑区烟草连作对土壤腐殖质性质的影响. 水土保持学报, 2007, 21(6): 126-129.
[6] 鲁如坤. 土壤农业化学分析方法. 北京: 中国农业科技出版社, 2000. 22, 28, 50-57, 106-109, 205-214.
[7] 姚槐应, 黄昌勇. 土壤微生物生态学及其实验技术. 北京: 科学出版社, 2006. 139.
[8] 关松荫. 土壤酶及其研究法. 北京: 农业出版社, 1986. 274-320.
[9] Anderson D W, Paul E A. Organ-mineral complexes and their study by radiocarbon dating. Soil Science Society of America Journal, 1984, 48(2): 298-301.
[10] 张甲坤, 陶澍, 曹军. 中国东部土壤水溶性有机物含量与地域分异. 土壤学报, 2001, 38(3): 308-314.
[11] Descheemaeker K, Muys B, Nyssen J, Sauwens W, Haile M, Poesen J, Raes D, Deckers J. Humus form development during forest restoration in exclosures of the Tigray highlands, Northern Ethiopia. Restoration Ecology, 2009, 17(2): 280-289.
[12] Trap J, Bureau F, Perez G, Aubert M. PLS-regressions highlight litter quality as the major predictor of humus form shift along forest maturation. Soil Biology and Biochemistry, 2013, 57(2): 969-971.
[13] 祝振华, 刘增文, 袁娜, 杜良贞. 黄土高原小叶杨与其他树种枯落叶混合分解对养分释放的影响. 西北林学院学报, 2012, 27(2): 1-5.
[14] Bahri H, Rasse D P, Rumpel C, Dignac M F, Bardoux G, Mariotti A. Lignin degradation during a laboratory incubation followed by13C isotope analysis. Soil Biology and Biochemistry, 2008, 40(7): 1916-1922.
[15] Balaria A, Johnson C E. Compositional characterization of soil organic matter and hot-water-extractable organic matter in organic horizons using a molecular mixing model. Journal of Soils and Sediments, 2013, 13(6): 1032-1042.
[16] 窦森, 王帅. 不同微生物对形成不同腐殖质组分的差异性研究进展. 吉林农业大学学报, 2011, 33(2): 119-125.
[17] Grinhut T, Hadar Y, Chen Y. Degradation and transformation of humic sub-stances by saprotrophic fungi, processes and mechanisms. Fungal Biology Reviews, 2007, 21(4): 179-189.
[18] Gnankambary Z, Bayala J, Malmer A, Nyberg G, Hien V. Decomposition and nutrient release from mixed plant litters of contrasting quality in an agroforestry parkland in the south-Sudanese zone of West Africa. Nutrient Cycling in Agroecosystems, 2008, 82(1): 1-13.
[19] Gallardo A, Merino J. Leaf decomposition in two Mediterranean ecosystems of Southwest Spain: influence of substrate quality. Ecology, 1993, 74(1): 152-161.
[20] 党亚爱, 李世清, 王国栋. 黄土高原典型区域土壤腐殖酸组分剖面分布特征. 生态学报, 2012, 32(6): 1820-1829.
[21] Fontana A, Pereira M G, Cunha dos Anjos L H, de Melo Benites V. Distribution of organic carbon in the humic fractions of diagnostic horizons from Brazilian soils. Communications in Soil Science and Plant Analysis, 2008, 39(7/8): 951-971.
[22] Güsewell S, Gessner M O. N: P ratios influence litter decomposition and colonization by fungi and bacteria in microcosms. Functional Ecology, 2009, 23(1): 211-219.
[23] Andersson S, Nilsson S I, Saetre P. Leaching of dissolved organic carbon (DOC) and dissolved organic nitrogen (DON) in more humus as affected by temperature and pH. Soil Biology and Biochemistry, 2000, 32(1): 1-10.
[24] 张林海, 王瑞申. 闽江河口秋茄湿地土壤腐殖质组成及剖面分布特征. 云南地理环境研究, 2011, 23(1): 5-10.
[25] Xu D, Zhou X, Wang X K. Adsorption and desorption of Ni2+on Na-montmorillonite: Effect of pH, ionic strength, fulvic acid, humic acid and addition sequences. Applied Clay Science, 2008, 39(3/4): 133-141.
[26] Feng K, Wang X L, Wang X Z, Tang Y. Relationship between 2:1 mineral structure and the fixation and release of cations. Pedosphere, 2003, 13(1): 81-86.
[27] 闫德仁. 人工林土壤腐殖质特性和土壤酶活性的研究. 林业科技, 1997, 22(5): 12-14.
[28] Tabatabai M A, García-Manzanedo A M, Acosta-Martínez V. Sub-strate specificity of arylamidase in soils. Soil Biology and Biochemistry, 2002, 34(1): 103-110.
[29] 吴炳孙, 吴敏, 韦家少, 何鹏, 佘贵连, 张焱华. 橡胶人工林地土壤腐殖质组成与有效态微量元素含量的研究. 热带农业科学, 2009, 29(3): 4-9.
[30] Peng X J, Luan Z K, Zhang H M. Montmorillonite-Cu (II)/Fe (III) oxides magnetic material as adsorbent for removal of humic acid and its thermal regeneration. Chemosphere, 2006, 63(2): 300-306.
[31] Bonanomi G, Incerti G, Antignani V, Capodilupo M, Mazzoleni S. Decomposition and nutrient dynamics in mixed litter of Mediterranean species. Plant and Soil, 2010, 331(1/2): 481-496.
[32] 周桂玉, 窦森, 刘世杰. 生物质炭结构性质及其对土壤有效养分和腐殖质组成的影响. 农业环境科学学报, 2011, 30(10): 2075-2080.
Humus differentiation and correlation with other soil properties under pure forest in semi-arid loess hilly area
ZHANG Xiaoxi1, LIU Zengwen2,3, ZHU Bochao2, BING Yuanhao4, YU Qi4, Luc Nhu Trung1,5
1InstituteofSoilandWaterConservation,NorthwestAgricultuceandForestryUniversity,Yangling712100,China2CollegeofNaturalResourcesandEnvironment,NorthwestAgricultuceandForestryUniversity,Yangling712100,China3KeyLaboratoryforAgriculturalResourcesandEnvironmentalRemediationinLoessPlateauofAgricultureMinistryofChina,Yangling712100,China4CollegeofForestry,NorthwestAgricultuceandForestryUniversity,Yangling712100,China5DepartmentofAgricultureRuralDevelopmentofLaoCai,LaoCaiCity330100,Vietnam
Long-term growth or continued planting of pure forests which composed of one single species would cause soil humus differentiation both in content and composition, so studying the characteristics of humus differentiation and its correlation with other soil properties may provide scientific basis for the management and reformation of pure forests. In this study, 8 typical pure forest soils in Nanniwan forest station (which located at the semi-arid loess hill area) were sampled from 24 plots and their humus and other soil properties were measured systematically. The results showed that: (1) the soil fromPlatycladusorientalisforest had the highest humus content (34.61 g/kg) but moderate humification degree and stability; the soils fromUlmuspumilaandBetulaplatyphyllahad moderate humus content (19.69—23.58 g/kg) but highest humification degree and stability; the soils fromAcerginnalaandPopulussimoniiforestshad both moderate humus content (20.59—22.53 g/kg) and composition; the soils fromPinustabulaeformis,HippophaerhamnoidesandRobiniapseudoaciaforests had both low humus content (11.77—13.81 g/kg) and lowest humification degree and stability. (2) According to correlation analysis among humus content and other soil properties, humic acid had significant (P<0.05) positive interaction with CEC, N (total and available N), microbe biomass and protease activity (correlation coefficient 0.769—0.926, the same below), but significant negative with available Cu (-0.793); fulvic acid had significant positive interaction with N, CEC, microbe biomass, sucrase and phosphatase activities (0.836—0.955), but significant negative with available Cu (-0.822); humin had significant positive interaction with N, CEC, microbe biomass and phosphatase activity and available Zn (0.766—0.951), but significant negative with dehydrogenase activity (-0.784). (3) According to correlation analysis among humus composition and other soil properties, no significant correlation (P>0.05) among humus composition and other soil properties was observed. However, the ratio of humic acid/total humus acid would be enhanced by protease, sucrase and peroxidase to some extent, but restrained by dehydrogenase; the ratio of humic acid/fulvic acid would be enhanced by available K, CEC and urease to some extent, but restrained by dehydrogenase. (4) In general, humus content had more observable correlation with other soil properties than humus composition Increasing N fertilize and increasing K fertilize were helpful for the improvement of humus content and composition, respectively. Forming mixed forests or increasing the undergrowth was the essential measure to promote the soil humification process and to solve the problem of soil degradation.
pure forest; humus; differentiation characteristic; soil properties
国家自然科学基金(30471376)
2013-05-20;
日期:2014-04-25
10.5846/stxb201305201116
*通讯作者Corresponding author.E-mail: zengwenliu2003@aliyun.com.cn
张晓曦,刘增文,朱博超,邴塬皓,于齐,Luc Nhu Trung.半干旱黄土丘陵区纯林土壤腐殖质异化特征及与其他性质的关系.生态学报,2015,35(6):1779-1787.
Zhang X X, Liu Z W, Zhu B C, Bing Y H, Yu Q, Luc Nhu Trung.Humus differentiation and correlation with other soil properties under pure forest in semi-arid Loess Hilly Area.Acta Ecologica Sinica,2015,35(6):1779-1787.
