秋季东海溶解态和颗粒态氨基酸的组成与分布*
2015-03-08张鹏燕杨桂朋
张鹏燕 陈 岩 杨桂朋
(中国海洋大学 化学化工学院 青岛 266100)
氨基酸是有机氮主要的存在形式, 同时也是大部分生物体有机碳的重要成分之一(Degens et al,1976)。颗粒态氨基酸含量占总颗粒有机碳的 3%—50%, 占总颗粒氮含量的 9.3%—104%(Ditmar et al,2001)。不稳定的有机物为异养生物提供食物和能量,也为河流、河口和沿海水域的初级生产提供营养物质(Mayer et al, 1998)。对控制氨基酸丰度和组成上有更深入的了解, 可以为不稳定有机物的来源和生物化学循环提供有效途径(Aufdenkampe et al, 2001)。溶解态氨基酸(THAA)包括溶解游离氨基酸(DFAA)和溶解结合态氨基酸(DCAA), 其中DCAA的来源和去除过程受浮游植物和细菌的影响(Burdige, 1991)。DFAA为细菌提供氮源和碳源, 同时在无机氮浓度低的环境下为微藻提供营养物质(Linares, 2006)。颗粒态氨基酸(PAA)是颗粒有机物的重要组成部分, 为浮游生物和底栖生物提供生源要素和氮。海水中溶解态氨基酸和颗粒态氨基酸的来源和去除与浮游植物、浮游生物、细菌和底栖生物的生长有紧密关系, 因此了解氨基酸的组成、浓度变化以及与环境因子之间的关系,对认识海洋生物地球化学循环有着重要的意义。
东海是中国海洋生产力最高的海区, 也是我国海洋研究的重点海域。东海沿岸的上升流是近岸环流的重要组成部分, 它能把低温、高盐和富含营养盐的深底层海水带到真光层, 为浮游生物的光合作用提供充足的养料, 从而影响海区的初级生产力的分布和生物资源量(王甜甜等, 2006)。影响东海的陆源输入和水团比较复杂, 主要河流是长江、钱塘江、闽江等, 黑潮水系、沿岸水系和混合水系是东海的主要水团。
目前, 国外对溶解态氨基酸和颗粒态氨基酸的研究已广泛展开(Spitzy et al, 1991; Rosenstock et al,1993; Svensson et al, 2004), 国内对河流和邻近海域溶解态氨基酸的调查不断完善(Yang et al, 2009; 陈岩等, 2010), 但对我国近海 PAA的研究却还只是起步阶段, 关于东海海域PAA的研究报道甚少(刘宗广等, 2013; Zhu et al, 2014)。因此, 本文选取东海海域为研究目标, 对东海海域的溶解态和颗粒态氨基酸的浓度分布和组成进行了分析, 并进一步讨论了其与环境因子特别是叶绿素a(chl a)之间的相关情况。
1 材料与方法
1.1 样品采集和保存
于2012年10月20日至26日(秋季)随“科学三号”调查船对东海海域进行了取样(如图1), 共4个断面。表层海水用12L Niskin Rosette采水器进行采集, 同时现场的温度、盐度和深度等相关参数由温盐深度仪(SBE-911CTD)同步测定, 采集后立即低压过滤, 所用的滤膜是已经在 450°C箱式电阻炉(SX-12-10型)中高温灼烧过 4个小时的玻璃纤维滤膜(Whatman GF/F), 过滤后将滤液装进样品瓶, 滤膜用锡纸包好,将所得的水样和滤膜放在-20°C的环境下冷冻保存。此外, 采集海水样品用Whatman GF/F玻璃纤维滤膜过滤, 滤膜包裹于锡纸内, 冷冻保存, 用于测定海水中chl a的含量。

图1 2012年秋季东海调查站位示意图Fig.1 Locations of sampling stations in the East China Sea in October 2012
1.2 样品分析
1.2.1 DFAA、THAA、DCAA和 PAA浓度的测定仪器: 美国 Waters Alliance ® HPLC液相色谱仪,Waters e2695系统, e2475荧光检测器(检测条件λex=337nm, λem=454nm), Agilent ZORBAX Eclipse 氨基酸分析柱。
流动相: A是0.05mol/L的醋酸钠缓冲溶液(pH=7.2±0.05), 醋酸钠缓冲溶液∶四氢呋喃(V/V)=100∶1;B是甲醇; C是Milli-Q水(由S-A10 Milli-Q超纯水系统制得); D是乙腈∶甲醇∶Milli-Q水(V/V/V)=1∶1∶1; 流动相均经过0.2μm PALL Supor-450滤膜过滤。
分析方法: OPA-MPA柱前衍生高效液相色谱法(对Lindroth和Mopper等人确定的方法进行调整)。
取 760μL海水样品加入有 240μL衍生化试剂的进样瓶中, 衍生化10min后自动进样(v=20μL)用流动相梯度洗脱分离方法进行测定。由此得到的是DFAA的浓度。
取 2mL海水样品, 40μL 0.2% (v/v)的抗坏血酸,2mL浓盐酸(12mol/L)于5mL安瓿瓶中, 冲入N2吹扫2min, 用酒精喷灯封口, 在 110°C环境下水解 22h。水解完成后, 敲开安瓿瓶瓶口, 放置到 65°C的真空干燥箱中干燥, 等干燥完成后, 取 2mL Milli-Q水溶解, 然后按照测定DFAA浓度的方法进行测定。扣去空白, 由此得到的是THAA的浓度。THAA的浓度减去DFAA的浓度得到DCAA的浓度。
滤膜冻干后, 将滤膜、5mL Milli-Q水、100μL抗坏血酸和5mL (6mol/L)盐酸加入到10mL的安瓿瓶中,同DCAA一样水解干燥后, 取10mL Milli-Q水加入安瓿瓶中将固体溶解后, 移入离心管中, 离心10min,立刻吸取760μL上清液(防止膜的吸附), 如同DFAA浓度的测定。扣去空白, 由此得到PAA的浓度。
浓度为 0.25μmol/L标准样品的 HPLC色谱图如图2所示, 每种氨基酸利用标准样品的保留时间来定性, 外标法定量, 每种氨基酸的标准曲线相关系数均在 0.995以上, 回收率采用空白加标工作曲线法, 所测的 14种氨基酸中, 大部分氨基酸的回收率在 75%—98%之间, 除了蛋氨酸和苯丙氨酸的回收率较低, 分别为59.7%、52.3%, 这两种氨基酸的回收率偏低可能是在处理样品的过程中发生了转化, 所以该方法对蛋氨酸和苯丙氨酸的定量测定还有待进一步探讨。
1.2.2 chl a浓度的测定 用90%(v/v)丙酮水溶液在暗处萃取冷冻保存的滤膜, 用SC-3610低速离心机离心, 离心完成后取上清液用日立 F-4500荧光分光光度计测定。
1.2.3 溶解有机碳(DOC)和溶解无机氮(DIN)测定方法 DOC测定方法为所取样品用岛津总有机碳分析仪TOC-VCPH(刘莉等, 2013), 在680°C高温催化氧化条件下测定, 用邻苯二甲酸氢钾配制好标准溶液得到标准曲线, 样品的浓度根据标准曲线得到, 其相对标准偏差<2%。
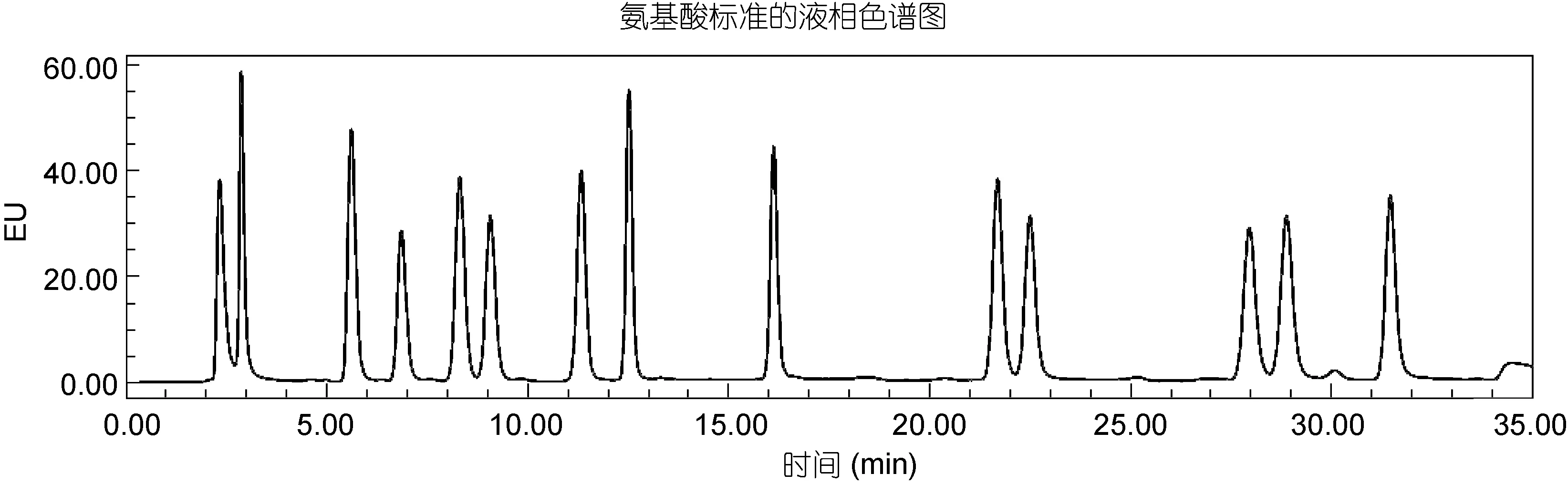
图2 氨基酸标准的液相色谱图Fig.2 Chramatogram of the amino acid standards
采用Seal Analytical AA3营养盐自动分析仪进行测定, 测得总氮(TN)、NO3–-N、NO2–-N 与 NH4+-N 的浓度, 其中 NO3–-N、NO2–-N 与 NH4+-N 的浓度之和为DIN的浓度, 溶解有机氮(DON)的浓度为TN浓度与DIN浓度之差。
2 结果与讨论
2.1 THAA、DFAA和DCAA浓度的水平分布特征
在所调查海域, THAA 的浓度在 0.21—1.98μmol/L, 平均浓度为 0.73μmol/L; 与伊势湾海水中氨基酸的浓度相似(Garrasi et al, 1979)。DFAA的浓度较低在 0.06—0.19μmol/L 之间, 平均浓度为0.12μmol/L; DCAA的浓度为0.15—1.79μmol/L, 平均浓度为0.61μmol/L。DFAA占THAA含量不到20%,而 DCAA的含量占 THAA达 80%以上。由此可见DCAA是THAA中的主要成分。THAA中N的摩尔数与DIN中N的摩尔数的比值范围为0.025—0.84(平均值为0.28)。
由图 3可知, DFAA的水平分布规律不明显。DFAA和 DCAA浓度的最大值均出现在台湾海峡北部, 此海域虽然叶绿素平均值为 0.64μg/L高于平均值 0.47μg/L, 但此处 DIN 的平均值仅为 6.33μmol/L低于东海整体的平均值8.11μmol/L, DON的平均值为64.31μmol/L高于东海整体的平均值46.74μmol/L, 这是由于夏季此处受台湾暖流影响最为明显, 由台湾暖流带来的高温高盐海水为浮游植物的生长提供所需的营养物质, 从而使该海域浮游植物大量生长(刘莉等, 2013), 生物活动将无机氮转化为有机氮, 使DON浓度增加, DFAA的含量增加; 而进入秋季, 台湾暖流的影响减弱, 营养盐含量减少, 但浮游植物并未立刻减少, 从而出现此海域DFAA、叶绿素和DON浓度较大, 而DIN浓度为6.33μmol/L低于周围海域。在DH5-1至DH5-4海域叶绿素平均浓度为0.84μg/L,DIN的平均浓度为13.60μmol/L, DON的平均浓度为69.72μmol/L, 都远高于其整体平均浓度, 这是由于长江冲淡水中营养盐含量非常高, 其中硝酸盐的含量最高可能达到100μmol/L(张传松等, 2007), 强大的长江冲淡水将大量的营养盐带入该海域, 使浮游植物生长旺盛。除受长江冲淡水的影响外, 可能还受钱塘江等河流输入的影响(米铁柱等, 2012), 从而使DCAA出现高值; 而 DFAA在此海域的平均浓度为0.085μmol/L, 明显低于整体平均值, 这是因为DFAA容易受细菌和浮游植物等的影响, 如生物群的胞外释放、浮游动物的新陈代谢等, 是海水中最易受生物活动影响的组分。在DH4-5、DH4-6和DH4-7海域,chl a浓度(0.56μg/L)高于周边海域 chl a浓度(0.35μg/L), DCAA的平均浓度为1.08μmol/L, 这是由于受到高温高盐的黑潮水系的影响, 浮游植物在丰富的营养盐类物质和适宜的水温条件下生长旺盛,从而导致该海域 DCAA较周围海域有所增多。DH3-1、DH3-2和 DH3-3站位 DIN的平均值为13.43μmol/L, chl a和 DON 的浓度分别为 0.41μg/L、10.06μmol/L, 低于平均浓度, 由于长江水虽然带来了丰富的营养盐, 但此处海水的搅动和泥沙, 使该海域的透明度低, 浑浊度大, 浮游植物受光照的限制(林军等, 2011), 未能将无机氮转化为有机氮, 因而chl a的浓度并未增大, DON浓度较周围浓度低, DIN浓度远高于其平均浓度, 说明此海域主要受长江富营养化河水输入作用(张传松等, 2007), 因而THAA、DCAA浓度较周围海域有所增高。
2.2 PAA浓度的水平分布特征
东海表层海水中 PAA的浓度范围是 0.02—0.27μmol/L, 平均浓度为 0.11μmol/L, 这与刘宗广等(2013)对东海PN断面的调查结果相似。由图3可知,PAA的水平分布特点是由近岸海域向远海海域分布呈现逐渐减小的趋势, 在DH5-1站位, PAA出现最大值 0.27μmol/L, 同时该站位的 chl a也出现最大值1.69μg/L, DIN出现最大值34.57μmol/L, 这促使该海域浮游植物的大量繁殖并快速代谢, 浮游植物未被完全腐烂的尸体所组成的有机碎屑和颗粒(Lee et al,1982)。并且 DH5-1站位位于渔山渔场附近, 海洋生物本身及其自身的排泄物也是该海域 PAA浓度高于周围海域的重要原因。
2.3 THAA、DCAA、DFAA和PAA与chl a的相关性分析
溶解态氨基酸和颗粒态氨基酸都可来源于浮游植物, 而叶绿素可以一定程度地反映浮游植物的数量。受东海大陆沿岸水团(S<31)影响的海域, 含有丰富的营养盐, 浮游植物生长旺盛, chl a浓度高于周围海域。该海域THAA 、DCAA和PAA与chl a有很好的相关性, 说明东海大陆沿岸 THAA、DCAA和PAA的来源和去除与浮游植物活动密切相关,THAA、DCAA和PAA受陆源输入影响较大; 在台湾暖流(33<S<34)影响海域, THAA、PAA 和 chl a浓度较周围海域高。受暖流影响, 浮游植物增多, 使该海域有机质也随之增加; 在黑潮表层水团(34<S<35)影响的海域, chl a浓度随盐度的增加而降低, THAA和chl a没有相关性, 这主要是因为黑潮次表层水涌升,将营养盐等物质带入东海(Chen, 2011), 从而使溶解态氨基酸浓度升高。如图4所示, THAA、DCAA和PAA与chl a整体上有很好的相关性, 说明浮游植物在控制THAA、DCAA和PAA浓度方面发挥着重要的作用。将DFAA与chl a进行线性拟合, 并未发现相关性, 这是因为DFAA在海水中的含量很少, 约占THAA的1%—35%左右(Lee et al, 1982); DFAA的释放及摄食与细菌活动紧密相连(Behrends et al, 1999),浮游植物不但是海水中DFAA的生产者, 也是其消耗者(陆田生等, 1997); DFAA的变化是海水中浮游植物、浮游生物、细菌和河流输入等不同的生物化学过程共同作用的结果。
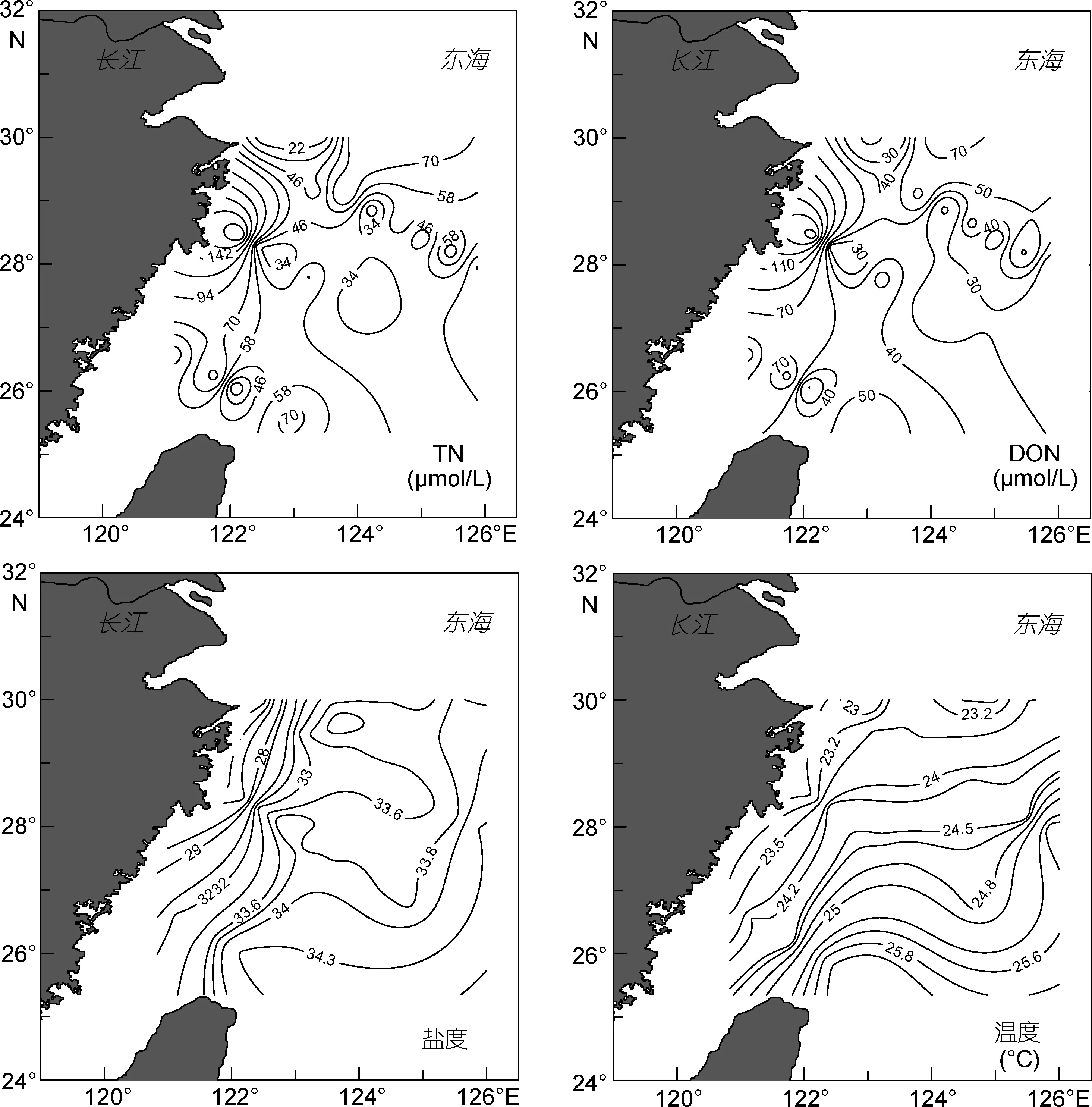
图3 东海表层海水中溶解态氨基酸、颗粒态氨基酸和环境因子的水平分布图Fig.3 Horizontal distributions of dissolved amino acids, particulate amino acids and environmental factors in the surface seawater of the East China Sea
2.4 东海海水中溶解氨基酸的垂直分布
选取第四断面进行溶解态氨基酸的垂直分析,由图 5可知, THAA和 DCAA的变化趋势相似。在DH4-1至 DH4-5海域 DCAA的垂直变化较复杂,DH4-5至DH4-8海域浓度变化较简单, 大体呈现由表层向底层浓度逐渐降低的趋势。由表1可知, DH4-2站位, 65m处THAA、DIN的浓度比30m处的浓度高,chl a的浓度比30m处的低, 出现这种现象的原因主要是由于海水底层沉积物中氨基酸的再悬浮、PAA的转化, 并且在深水处浮游植物减少, 对溶解态氨基酸的利用减少, 从而导致溶解态氨基酸在底层出现高值。在DH4-4站位, 35m处THAA、DCAA的浓度比表层和底层高, 同时该深度的DIN、chl a浓度也高于其它深度, 在秋季东海的盐跃层和温跃层向深层推移, 生物的生长速率和摄食行为增强(Liebezeit et al,1986)。
DFAA的垂直变化大体呈现表层高、底层低的趋势。由浅水层到深水层, 光照减少, 温度降低, 浮游植物生物量减少, 释放出的DFAA也减少, 在底层上升流引起间隙水中的氨基酸再次融入海水中(Henricha et al, 1979)。

图4 东海表层海水中THAA、DCAA、PAA、DCAA与chl a之间的相关性Fig.4 Relationships of THAA, DCAA, DFAA, PAA to chl a concentrations in the surface seawater of the East China Sea

表1 特殊站位溶解氨基酸及相关环境因子浓度的垂直分布Tab.1 Vertical distribution of the concentrations of THAA, DFAA, DCAA and environmental factors at some special stations
2.5 秋季东海海水中溶解态和颗粒态氨基酸组成
图 6所示为东海表层海水中 14种氨基酸在DFAA、THAA、DCAA和PAA中个体氨基酸所占的平均摩尔百分比。DFAA中的优势氨基酸是天门冬氨酸、谷氨酸、丝氨酸、甘氨酸和丙氨酸, 占DFAA的66.96%, 其中丝氨酸占的比例最大, 这与 Pettin等(1999)对意大利北部亚得里压海(Adriatic Sea)的调查结果相似。DCAA和THAA的优势氨基酸相似, 天门冬氨酸、谷氨酸、丝氨酸、甘氨酸和苏氨酸占总DCAA的71.02%。PAA中天门冬氨酸、谷氨酸、丝氨酸、甘氨酸、丙氨酸和亮氨酸占较大比例, 它们的总和占PAA的69.37%, 这与Duan等(2007)对美国密西西比河和我国珠江的调查结果类似。丙氨酸和甘氨酸占PAA的 31.60%, 这与 Liebezeit等(1986)对南极布兰斯菲尔德海峡(Bransfield Strait)调查的结果有差异。这种差异主要是因为东海海水中硅藻是优势藻种,而硅藻细胞的主要构成成分是天门冬氨酸、丝氨酸和谷氨酸等(Kirchman et al, 1984)。硅藻死亡后所形成的有机碎屑, 使海水中颗粒态氨基酸的含量增加。综合看来, 东海表层水中氨基酸以天门冬氨酸、谷氨酸、丝氨酸、甘氨酸为主, 它们也是构成藻类蛋白质的主要氨基酸, 秋季东海海域藻种为聚生角毛藻(罗民波等, 2009), 藻类会通过光合作用, 细胞破裂, 被浮游生物摄食等方式, 将细胞内的氨基酸释放到海水中称为溶解态氨基酸和颗粒态氨基酸的重要来源。

图5 秋季东海海水中溶解态氨基酸及环境因子的垂直分布Fig.5 Vertical profiles of dissolved amino acids and environmental factors at transect 4 in the East China Sea in autumn

图6 东海表层海水中DFAA、THAA、DCAA和PAA中个体氨基酸的平均摩尔百分比Fig.6 The mean molar percentage of individual amino acid in THAA, DFAA, DCAA and PAA in the surface seawater of the East China Sea
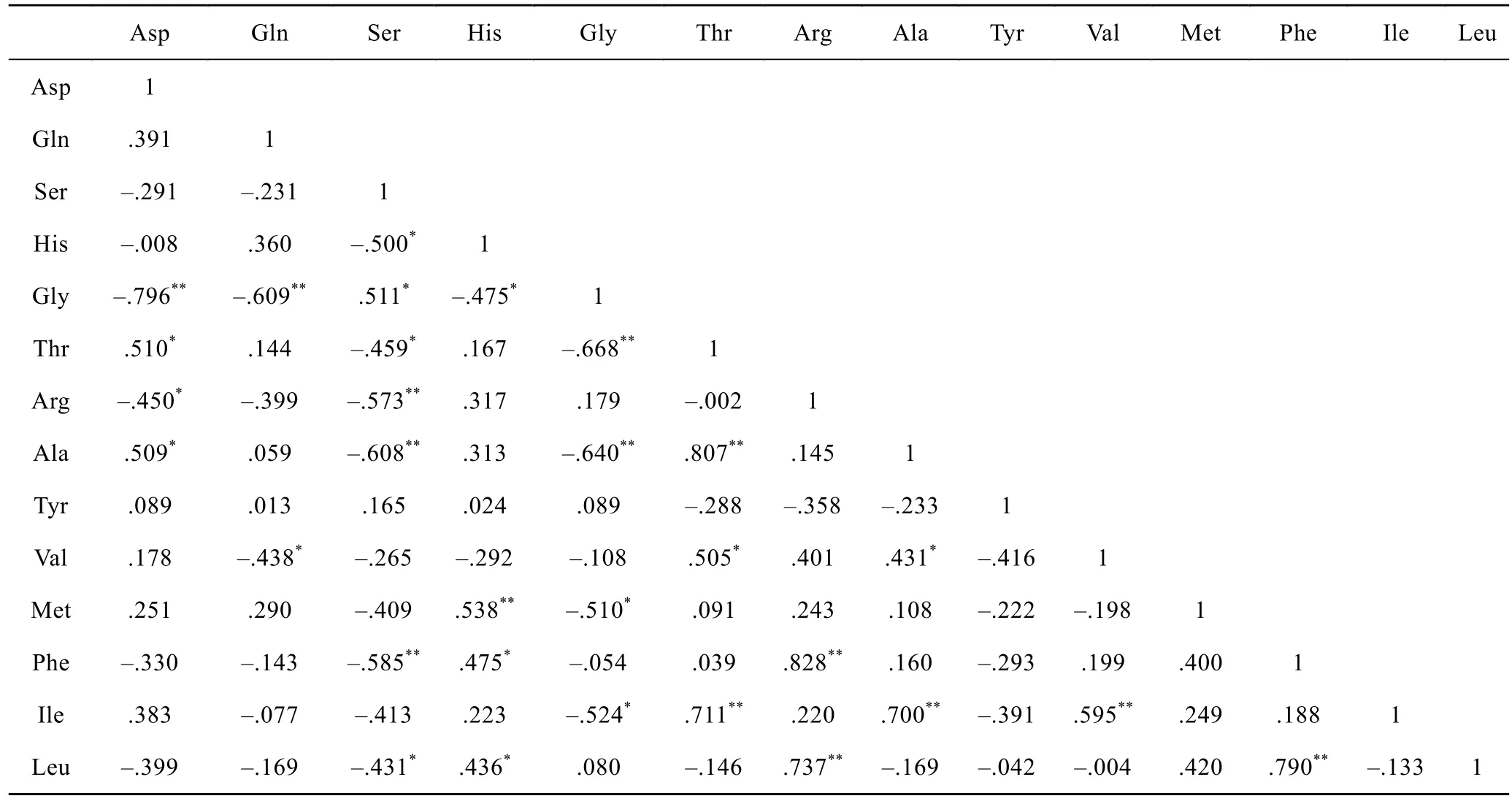
表2 秋季东海表层海水PAA中个体氨基酸相对含量的相关性矩阵Tab.2 Matrix of correlation of individual amino acid in relative abundance (mol%) of PA
以PAA为例, 基于氨基酸的标准化数据(表层海水中每种氨基酸占PAA的摩尔百分比, 如表2所示)做相关性矩阵, 来评估量化个体氨基酸在表层海水中动态行为的异同。来自 21个站位的数据显示有 7对氨基酸(甘氨酸-天门冬氨酸、甘氨酸-谷氨酸、苏氨酸-甘氨酸、精氨酸-丝氨酸、丙氨酸-丝氨酸、丙氨酸-甘氨酸、苯丙氨酸-丝氨酸)之间存在负相关性, 表明它们在生物地球化学过程中表现出不同的动态行为。同时也有 8对氨基酸(丙氨酸-苏氨酸、蛋氨酸-组氨酸、苯丙氨酸-精氨酸、异亮氨酸-苏氨酸、异亮氨酸-丙氨酸、异亮氨酸缬氨酸、亮氨酸-精氨酸、亮氨酸-苯丙氨酸)之间存在正相关性, 这些关系表明它们是作为一个整体在生物地球化学变化机制中产生相似的作用。
2.6 降解指数评价颗粒态氨基酸来源
PAA经过主成分分析(PCA)降维变换后得到几个新的主成分。用降解指数DI来评估氨基酸的来源和降解情况(Dauwe et al, 1998), 在数据分析中, 只采用第一主成分进行降解指数的计算, 其计算结果如表 3和图7所示。

表3 秋季东海表层海水中PAA%的PAC参数Tab.3 Parameters of PAC in relative abundance (mol%) of PAA in the surface seawater of East China Sea in autumn
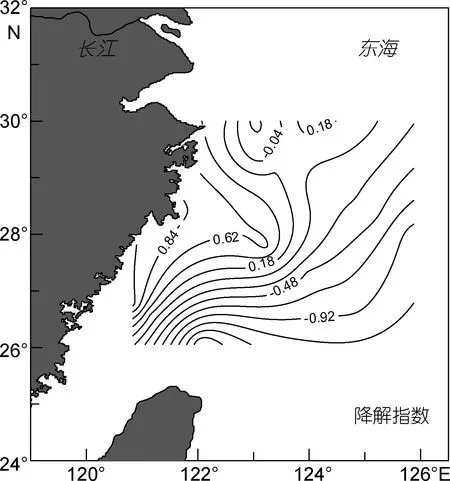
图7 秋季东海表层海水中降解指数的水平分布Fig.7 Horizontal distribution of degradation index in the surface seawater of the East China Sea during autumn
由主成分分析得知, 第一主成分特征值占总方差的 34%, 并且与丝氨酸、甘氨酸和缬氨酸成负相关。由图7可知, DI的范围是-1.8—1.2, DI的数值越接近1.2, 表明PAA的降解程度越低, 氨基酸主要是由浮游植物现场产生的; DI的数值越接近-1.8, 表明PAA的降解程度越高, 可以推断其来源主要是降解程度高的沉积物(Dauwe et al, 1998)。PAA的DI大体呈现近岸降解程度比远岸降解程度低, 这是因为由近岸到远岸, 陆源输入的贡献降低, 浮游植物生长所需的条件减弱, 所以远岸海域的 PAA主要是由高度降解的悬浮颗粒物产生的。DI的高值出现在 DH8-1和DH5-1站位, chl a浓度也较高, 表明该海域浮游植物的现场生产对PAA的贡献较大; DI的低值出现在DH4-8和DH8-5海域, 该海域远离近岸, 浮游植物对PAA的贡献较小, 在该海域悬浮颗粒物对PAA的贡献增大。
3 结论
(1)秋季东海表层海水 THAA的平均浓度为(0.72±0.51)μmol/L (0.21—1.98μmol/L), DFAA、DCAA和PAA的平均浓度分别为(0.12±0.04)μmol/L (0.06—0.19μmol/L)、(0.61±0.51)μmol/L (0.15—1.79μmol/L)、(0.11±0.06)μmol/L (0.02—0.27μmol/L)。 DCAA 是THAA中的主要组成部分。THAA和PAA的分布大致呈现出近岸高于远岸的趋势, 在闽浙沿岸、台湾海峡北部海域 THAA浓度高于周围海域, 可能与台湾暖流、上升流和沿岸河流输入有关。PAA的浓度分布与陆源输入、浮游植物的现场生产和周围海域渔场分布有关。
(2)THAA、DCAA和PAA与chl a有很好的相关性, 说明浮游植物生物量在控制 THAA、DCAA和PAA浓度方面发挥着重要作用。而DFAA与chl a并无明显相关性, 这主要是因为DFAA受多种环境因素的共同影响。
(3)研究表明: THAA中的优势氨基酸是天门冬氨酸、谷氨酸、丝氨酸、甘氨酸; PAA中天门冬氨酸、谷氨酸、丝氨酸、甘氨酸、丙氨酸和亮氨酸占较大比例, 它们的总和占PAA的69.37%。氨基酸个体在表层海水中是作为一个整体而对生物地球化学过程产生影响的。
王甜甜, 康建成, 李卫江等, 2006. 东海上升流研究进展. 见:中国地理学会 2006年学术年会论文摘要集. 北京: 中国地理学会, 7
刘 莉, 杨桂朋, 陈 岩, 2013. 秋季中国东海海水中溶解氨基酸的分布与组成. 海洋环境科学, 32(4): 510—517
刘宗广, 吴 莹, 胡 俊等, 2013. 东海陆架典型断面颗粒态氨基酸的分布及控制因素分析. 海洋与湖沼, 44(3):563—569
米铁柱, 姚庆祯, 孟 佳等, 2012. 2011年春、夏季黄海、东海营养盐分布特征研究. 海洋与湖沼, 43(3): 678—688
张传松, 王修林, 石晓勇等, 2007. 东海赤潮高发区营养盐时空分布特征及其与赤潮的关系. 环境科学, 28(11):2416—2424
陆田生, 纪明候, 1997. 小角刺藻生长过程中溶解游离氨基酸含量在海水中的变化. 海洋与湖沼, 28(3): 256—261
陈 岩, 杨桂朋, 2010. 春季北黄海表层海水中溶解游离氨基酸的分布与组成研究. 中国海洋大学学报, 40(7): 93—98
林 军, 朱建荣, 张 经等, 2011. 长江口外海区浮游植物生物量分布及其与环境因子的关系. 水产学报, 35(1):74—87
罗民波, 陆健健, 王云龙, 2007. 东海浮游植物数量分布与优势种. 生态学报, 27(12): 5076—5085
Aufdenkampe A K, Hedges J I, Richey J E et al, 2001. Sorptive fractionation of dissolved organic nitrogen and amino acids onto fine sediments within the Amazon Basin. Limnology and Oceanography, 46(8): 1921—1935
Behrends B, Liebezeit G, 1999. Particulate amino acids in Wadden Sea waters—seasonal and tidal variations. Journal of Sea Research, 41(1—2): 141—148
Burdige D J, 1991. Microbial processes affecting alanine and glutamic acid in anoxic marine sediments. FEMS Microbiology Letters, 85(3): 211—231
Chen C T A, 2011. Downwelling then upwelling again of the upwelled Kuroshio water in the southern East China Sea.Journal of Geophysical Research, 116(C7): C07003,http://dx.doi.org./10.1029/2011JC007030
Dauwe B, Middelburg J J, 1998. Amino acids and hexosamines as indicators of organic matter degradation state in North Sea sediments. Limnology and Oceanography, 43(5):782—798
Degens E T, Mopper K, 1976. Factors controlling the distribution and early diagenesis of organic material in marine sediments.In: Riley J P, Chester R eds. Chemical Oceanography. 2nd ed.New York: Academic Press, Inc, 59—113
Ditmar T, Fitznar H P, Kattner G, 2001. Origin and biogeochemical cycling of organic nitrogen in the eastern Arctic Ocean as evident from D- and L-amino acids.Geochimica et Cosmochimica Acta, 65(22): 4103—4114
Duan S, Bianchi T S, 2007. Particulate and dissolved amino acids in the lower Mississippi and Pearl Rivers (USA). Marine Chemistry, 107(2): 214—229
Garrasi C, Degens E T, Mopper K, 1979. The free amino acid composition of seawater obtained without desalting and preconcentration. Marine Chemistry, 8(1): 71—85
Henricha S M, Farrington J W, 1979. Amino acids in interstitial waters of marine sediments. Nature, 279(5711): 319—322
Kirchman D, Hodson R, 1984. Inhibition by peptides of amino acid uptake by bacterial populations in natural waters:Implications for the regulation of amino acid transport and incorporation. Applied and Environmental Microbiology,47(4): 624—631
Lee C, Cronin C, 1982. The vertical flux of particulate organic nitrogen in the sea: decomposition of amino-acids in the Peru upwelling area and the equatorial Atlantic. Journal of Marine Research, 40(1): 227—251
Liebezeit G, Bölter M, 1986. Distribution of particulate amino acids in the Bransfield Strait. Polar Biology, 5(4): 199—206
Linares F, 2006. Effect of dissolved free amino acids (DFAA) on the biomass and production of microphytobenthic communities. Journal of Experimental Marine Biology and Ecology, 330(2): 469—481
Mayer L M, Keil R G, Macko S A et al, 1998. Importance of suspended participates in riverine delivery of bioavailable nitrogen to coastal zones. Global Biogeochemical Cycles,12(4): 573—579
Pettine M, Patrolecco L, Manganelli S et al, 1999. Seasonal variations of dissolved organic matter in the northern Adriatic Sea. Marine Chemistry, 64(3): 153—169
Rosenstock B, Simon M, 1993. Use of dissolved combined and free amino acids by planktonic bacteria in Lake Constance.Limnology and Oceanography, 38(7): 1521—1531
Spitzy A, Ittekkot V, 1991. Dissolved and particulate organic matter in rivers. In: Mantoura R F C, Martin J M, Wollast R eds. Ocean Margin Processes in Global Change. New York:Wiley, 5—17
Svensson E, Skoog A, Amend J P, 2004. Concentration and distribution of dissolved amino acids in a shallow hydrothermal system, Vulcano Island (Italy). Organic geochemistry, 35(9): 1001—1014
Yang G P, Chen Y, Gao X C, 2009. Distribution of dissolved free amino acids, dissolved inorganic nitrogen and chlorophyll a in the surface microlayer and subsurface water of the Yellow Sea, China. Continental Shelf Research, 29(14): 1737—1747
Zhu Z Y, Wu Y, Zhang J et al, 2014. Can primary production contribute non-labile organic matter in the sea: Amino acid enantiomers along the coast south of the Changjiang Estuary in May. Journal of Marine Systems, 129: 343—349
