塔河流域不同生境条件下柽柳种群结构异质性特征1)
2015-03-08刘华董玲李丕军王文月贾瑞琪
刘华 董玲 李丕军 王文月 贾瑞琪
(安徽农业大学,合肥,230036) (新疆林业科学院造林治沙研究所)
责任编辑:任 俐。
植物种群结构是种群内不同大小、年龄个体数量的分布状况[1],反映种群个体与环境间的关系及其群落的发展和演变趋势[2]。研究种群结构不仅可以了解种群的现状,也有助于分析种群未来的数量动态。柽柳(Tamarix chinensis)主要分布在干旱区、半干旱区沙荒地和盐碱化土地上,在新疆分布的面积最广,是南疆地区荒漠河岸植被的主要灌木植被类型之一,具有耐干旱、耐盐碱、防风固沙等功能[3-4]。对其的研究多以分析柽柳属的系统学[5]、生态适应性[6]、抗旱性[7]、生物多样性[8]、空间分布[9]、植被群落[10]等特征,对塔河流域柽柳的种群结构及其维持机理的研究相对较少。塔河流域地处新疆南部,具有干旱多风、水资源贫乏、土壤成土过程缓慢、植被稀疏、生物多样性相对较低、食物链结构简单的特征。为了遏制荒漠河岸植被退化进程,改善和提高其生态功能,国家和地方政府相继启动了多项生态建设工程[11],由于塔河流域不同地点的生境差异极大,水源供应难易不同,栽植树木虽多却难以成林。且在实施过程中缺乏对植物种群结构及其机理的充分认识和了解,生态治理效果不明显。柽柳灌丛在干旱区无需任何灌溉保障之下,可以直接利用干旱区水源而自然生存繁殖,自发萌蘖形成丛林。从生态保护、社会发展及经济价值的角度,增加柽柳种群分布面积,提高其生态服务功能是维护区域生态环境的关键。因此,本研究以塔河流域柽柳种群为研究对象,分析在沙地、盐生地、岩性土地等3 种典型分布区域的种群结构特征及其关联因子,明确不同立地条件对柽柳种群的影响。这对科学实施荒漠植被系统的生态修复和种群生态保护提供科学的指导意义。
1 研究区概况
研究区位于新疆塔河流域中游段的巴音郭楞蒙古自治州西部的轮台县境内(41°05'~42°32'N,83°38'~85°25'E)。该区属温带大陆性干旱气候,年平均气温为10.6 ℃,年日照时间2 787 h,积温为4 040~4 210 ℃,极端最低气温-25.5 ℃,年平均降水量65.6 mm,最大蒸发量为2 077 mm,土壤以盐土和沙土为主,通透性较好,pH 值为7.8~8.5。植被类型为荒漠植被,优势乔木树种以特有的第三纪残余的胡杨(Populus euphratica)为主;灌木主要为柽柳、铃铛刺(Halimodendron halodendron)、盐穗木(Halostachys caspica)、琵琶柴(Reaumuria soongonica)、白刺(Ni-traria tangutorum)、骆驼刺(Alhagi sparsifolia);草本植物有角果藜(Ceratocarpus arenarius)、花花柴(Karelinia caspica)、蓼子朴(Inula salsoloides)、胀果甘草(Glycyrrhiza inflate)等。
2 研究方法
柽柳种群调查:依据植被盖度的定义及研究区柽柳种群抽样结果,将柽柳种群盖度(G)划分为20%≤G<40%(G1)、40%≤G <60%(G2)、G >60%(G3)共3 个等级,并分别在沙地、盐生地、岩性土地等3 种立地上设置面积为100 m×100 m 的大样地各3 块,记录样地的具体位置(经纬度、海拔),在大样地内按照10 m×10 m 的小样方调查并记录样方中所有灌木的名称、株数、地径、高度、冠幅和草本植物的种类、频度及盖度。调查样地概况见表1。
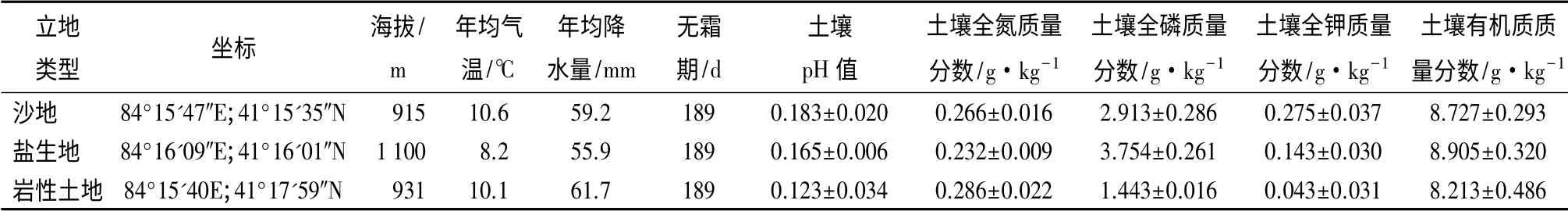
表1 不同立地调查概况
柽柳种群结构划分:本研究采用地径(D)和树高(H)2 个指标进行种群结构分析。按照上限排外法,以1 cm 为径阶,将径级划分为Ⅰ(D<1)、Ⅱ(1≤D<2)、III(2≤D<3)、IV(3≤D<4)、V(4≤D<5)、VI(5≤D<6)、VII(6≤D<7)、VIII(7≤D)共8 级。将树高划分为Ⅰ(H<0.5)、Ⅱ(0.5≤H<1)、III(1≤H<1.5)、IV(1.5≤H<2)、V(2≤H<2.5)、VI(2.5≤H<3)、VII(3≤H)共7 级。
数据处理:利用spss 软件进行统计分析和检验。
3 结果与分析
3.1 种群分布特征
研究区的柽柳主要分布在沙地、盐生地、岩性土地等3 种立地条件下,形成的以柽柳为优势种的群落,多以纯林形式出现,林下伴生着一些草本植物,共计8 科9 属9 种。主要有藜科盐穗木属的盐穗木(Halostachys caspica)、豆科骆驼刺属的骆驼刺(Alhagi sparsifolia)、铃铛刺属的铃铛刺(Halimodendron halodendron)、柽柳科琵琶柴属琵琶柴(Reaumuria soongonica)、蒺藜科白刺属的白刺(Nitraria tangutorum)、茄科枸杞属的苏枸杞(Lycium ruthenicum)、禾本科芦苇属的芦苇(Phragmites australis)、菊科旋覆花属的蓼子朴(Inula salsoloides)、蓼科角果藜属的角果藜(Ceratocarpus arenarius)。沙地条件下柽柳个体数量占总种群总数量的53.02%,盐生地的占29.46%,岩性土地的仅占17.52%。
3.2 不同立地条件种群径级和树高结构
3 种不同立地条件下柽柳种群的径级和树高结构见表2。不同立地条件下柽柳种群的个体数量随着径级的增大呈先增加后降低的单峰型变化趋势。在沙土立地条件下的柽柳种群径级结构相对完整,其中,III 径级的个体数量最高,占总株数的35.33%,明显高于其他径级的(p<0.05);Ⅱ径级的次之,其个体数量占到总株数的31.05%;Ⅱ、III 径级的个体占群体总量的66.38%;IV 径级的个体数量占种群数量的13.96%;Ⅰ、Ⅴ和Ⅷ径级的个体数量接近,Ⅵ、Ⅶ径级的个体数量较低。岩性土地柽柳种群中,径级大于IV 级的个体未出现。Ⅱ径级个体数量最多,III 径级的次之,Ⅰ径级个体数量占总体的15.52%;IV 径级最小。盐生地条件下柽柳种群中,Ⅰ径级和V 径级以上的个体未见分布,其中III 径级的个体数量最大,占总数量的46.67%;Ⅱ、IV 径级的个体数量接近,V 径级的最少。风沙地和盐生地柽柳种群的平均地径显著高于岩性土地的(p<0.05),但前二者之间差异不显著(p>0.05)。风沙地、盐生地和岩性土地平均地径分别为(2.73±0.11)cm、(2.68±0.06)cm 和(1.80±0.07)cm。
3 种不同立地条件下柽柳种群的树高结构的分布不同(表2)。沙地柽柳种群的树高结构较完整,与径级结构的变化规律一致,但Ⅱ级和III 级树高的个体数量分别占总体数量的53.43%和36.86%;Ⅰ级个体数量占总体的6%,IV~VII 级仅占总体的3.17%。岩性土地和盐生地条件下柽柳种群中IV级以上高度的个体数量没有出现。其中,在盐生地中随高度级的增加,相应的种群数量呈下降趋势,第Ⅰ高度级的个体数量占种群总量的46.15%,第Ⅱ、III 高度级的个体数量分别占总量的35.90%和5.13%,三者之间差异显著(p<0.05)。岩性土地中随着高度级的增加,柽柳种群数量呈上升趋势,第III 高度级的个体数量最多,占种群总量的61.54%,第Ⅱ、Ⅰ级分别占种群总量的36.92%和1.54%,三者之间差异显著(p<0.05)。在相同的径级下,不同立地条件柽柳种群的个体高度间差异极显著(p<0.01)。
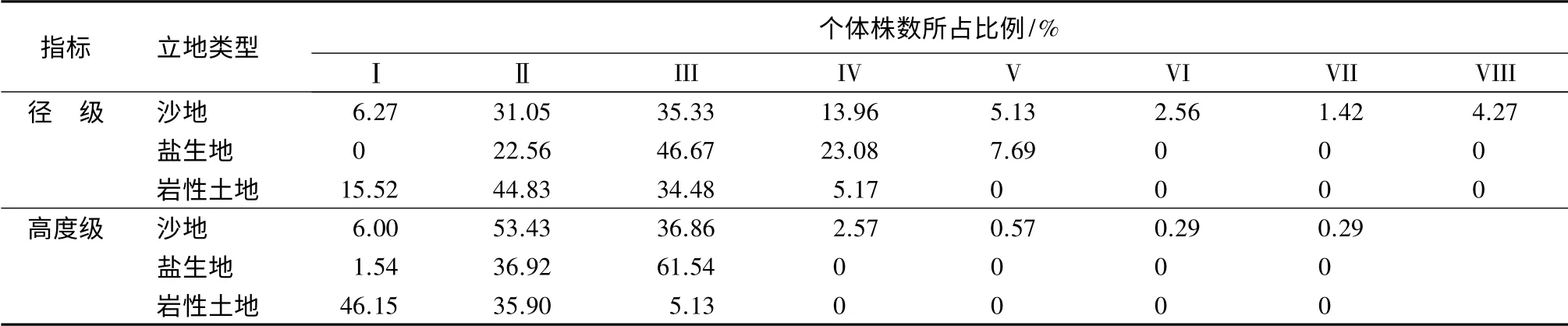
表2 3 种立地条件下不同径级和高度级柽柳个体株数所占比例
3.3 不同盖度沙地柽柳种群的径级和树高结构
表3为不同盖度下沙地柽柳种群的径级和高度级各个体数量分布。3 个盖度下的径级结构较为完整,但不同径级个体所占比例略有差异。G1和G2盖度的柽柳种群个体数量随径级的增大呈先增加后降低的趋势,且Ⅱ、III 径级的个体数量显著高于其他径级的(p<0.05)。其中,G1盖度中Ⅱ、III 径级的株数百分比差异不显著(p>0.05),而G2盖度中Ⅱ、III 径级的株数百分比差异极显著(p<0.01),前者是后者的2.1 倍。G3盖度的柽柳种群个体数量除Ⅱ级略低于III 级外,总体上呈随径级的增加而下降的趋势,Ⅰ径级个体数量显著高于Ⅱ、III 和IV 径级的(p<0.05),极显著高于Ⅴ~Ⅷ径级的(p<0.01),但Ⅱ、III和IV 径级之间差异不显著(p>0.05)。G1和G2盖度柽柳种群的平均地径显著低于G3盖度的(p<0.05),但前二者之间的差异不显著(p>0.05)。随着盖度的增加,柽柳种群中小径级个体数量明显增加,说明林地郁闭度的增加,林地植被固沙能力增加,为柽柳的天然更新创造了有力的条件,保证了种群结构的完整。G3盖度的柽柳大径级的个体所占比例明显高于G2和G1盖度的,说明林地郁闭度的增加为柽柳个体的存活及生长提供了良好的生境条件。
不同盖度下柽柳种群各树高级个体数量随高度级的增加呈先增加后降低的趋势(表3)。G1和G2盖度柽柳种群中V 级以上高度的个体数量很少,其种群主要分布在Ⅱ、III 高度级;G3盖度的柽柳种群树高结构为典型的正态分布,III 高度级个体数量所占比例达到35.22%,Ⅱ和IV 高度级、Ⅰ和Ⅴ高度级以及Ⅵ和Ⅶ高度级的个体数量比例接近。
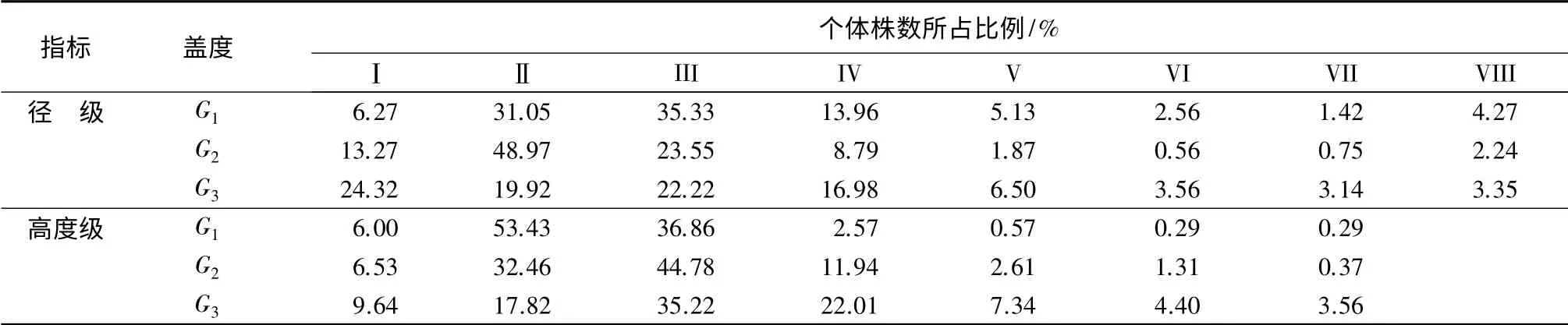
表3 沙地条件下不同盖度的柽柳种群各径级和高度级个体株数所占比例
3.4 种群生长与土壤特性的相关性
根据沙地、岩性土地和盐生地3 种立地条件下柽柳种群的分布特征可以看出,土壤理化性质与种群结构之间存在一定的关系,测定沙地、盐生地和岩性土地土壤的氮、磷、钾、钙、镁、有机质质量分数,土壤含水量及pH 值,分析土壤特性与该柽柳的株高和地径之间的相关性(表4)。沙地和盐生地柽柳的高径生长有显著的相关性(p<0.05),沙地柽柳高径生长的相关系数高于盐生地的,说明这两种立地条件下的株高生长直接受土壤理化性质的影响,高生长受粗生长的影响明显。而岩性土地的显著性不明显(p>0.05)。
沙地柽柳灌丛的地径生长与土壤全氮质量分数有着极显著的正相关关系(p <0.01),与土壤含水量、镁和有机质质量分数均呈显著正相关性(p<0.05);树高生长极显著受土壤含水量的影响(p<0.01),同时与土壤氮和有机质质量分数之间也呈显著正相关性(p<0.05)。盐生地柽柳灌丛地径生长仅与有机质质量分数呈极显著正相关性关系(p<0.05),与土壤含水量呈显著负相关关系(p<0.05);树高生长也与土壤含水量呈显著负相关关系(p<0.05),与土壤钾和有机质质量分数之间的相关系数分别为0.715 和0.710,但其相关性不显著(p>0.05)。岩性土地柽柳灌丛的高径生长均与土壤氮、有机质质量分数和土壤含水量之间呈显著的正相关关系(p<0.05)。
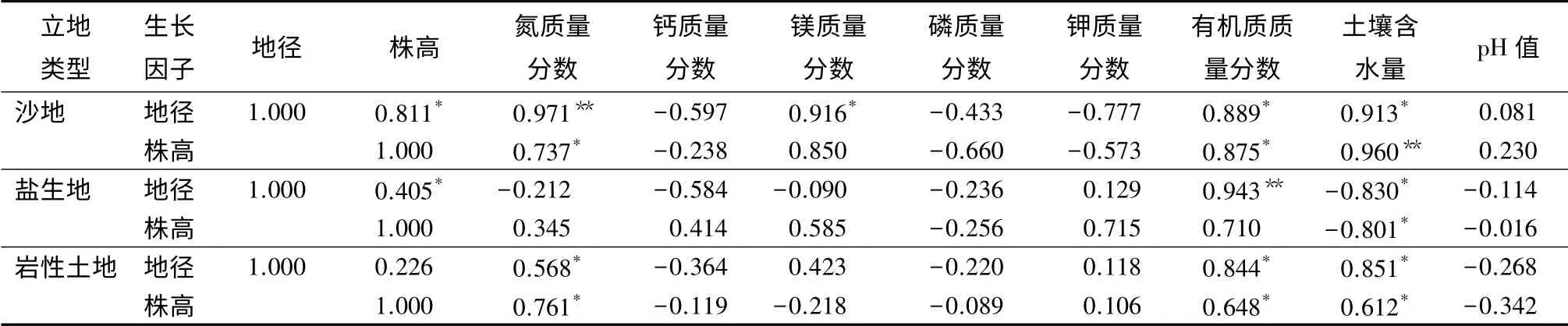
表4 沙地和盐生地柽柳灌丛生长与土壤特性的相关系数
4 结论与讨论
研究区内以柽柳为优势种的群落中伴生有盐穗木、骆驼刺、白刺、苏枸杞等旱生灌木,共计8 科9 属9 种。沙地、盐生地和岩性土地的柽柳种群个体的径级均以Ⅱ、III 级为主,高生长和粗生长关系显著(p<0.05),但沙地环境条件下柽柳种群结构较盐生地和岩性土地的相对完整,其生长主要受土壤氮质量分数的影响,其次为镁质量分数、土壤含水量及其有机质质量分数;盐生地柽柳种群主要受土壤有机质质量分数和土壤含水量的影响;岩性土地主要受土壤氮、有机质质量分数和含水量的影响。
柽柳属植物新疆的分布范围较广,在盐质量分数较高(30~50 g·kg-1)的土壤中均可以正常生长[12],这与柽柳植物的泌盐特征相适应[13]。但在沙地、盐生地和岩性土地等立地条件下,柽柳种群数量的差异明显,这是由于柽柳种子微小,吐絮后种子会随风飞扬,沙地和盐生地的植被盖度、水热条件及地表粗糙度等较岩性土地的优越,为柽柳种子的定居和萌发提供了较为适宜的场所。管海英等[14]研究表明,荒漠区植被盖度与土壤环境因子关系密切,地表植被的覆盖有效降低了土壤盐质量分数,利于土壤微生物的积累,增加了土壤肥力,促进植物的生长和碳水化合物的合成。文中的试验结果进一步显示在沙地和盐生地条件下柽柳种群的生长均受土壤有机质质量分数的影响。除种子更新之外,根蘖繁殖是研究区内柽柳种群天然更新的另一种主要方式,种群个体聚集性较强。随着龄级的增长,在有限的资源条件下,种内竞争加剧,优胜劣汰,种群数量逐渐减少,结合本研究,枯死木在柽柳种群中呈现出聚集分布。因此,在沙地、盐生地和岩性土地的柽柳种群个体的径级均以Ⅱ、III 级为主,种群的配置不均匀,成熟、过熟林木数量较少,种群处于生长状态;但沙地环境条件下柽柳种群结构较盐生地和岩性土地的相对完整。且随着盖度的增加,沙地的柽柳不同径级和高度级个体数量也呈增长的趋势,说明林地郁闭度的增加有效地阻止了沙丘的流动和土壤的侵蚀[15],为柽柳个体的存活及生长提供了良好的生境条件,特别是幼龄(Ⅰ级)个体数量及比例的增加,保证了种群的持续发展[16-17]。从土壤理化性质来看,土壤含水量是影响3 种立地环境中柽柳种群的主要因子,而沙地和岩性土地土壤含水量大小与柽柳高径生长之间呈显著正相关关系,盐生地的却相反,这是由于荒漠地区土壤蒸散强烈,随着地表蒸散作用的进行,土壤水分向地表运移的过程导致土壤表面返盐碱现象明显,有大量的盐分滞留在表层土壤中,随着降水的发生,又使土壤盐质量分数增加,也导致其肥力的降低,故对柽柳的生长是起到了一定的抑制作用[18]。
[1] Arista M.The structure and dynamics of an Abies pinsapo forest in southern Spain[J].Forest Ecology and Management,1995,74(1/3):81-89.
[2] Wu Xiaopu,Zheng Yu,Ma Keping.Population distribution and dynamics of Quercus liaotungensis,Fraxinus rhynchophlla and Acer mono in Dongling Mountain,Beijing[J].Acta Botanica Sinica,2002,44(2):212-223.
[3] 公维昌,庄丽,赵文勤,等.多枝柽柳与梭梭光合器官形态解剖结构的生态适应性[J].中国沙漠,2011,31(1):129-136.
[4] 曾凡江,张希明,李小明.柽柳的水分生理特性研究进展[J].应用生态学报,2002,13(5):611-614.
[5] 张道远,尹林克,潘伯荣.柽柳属(Tamarix L.)系统学研究历史及现状[J].干旱区研究,2002,19(2):41-46.
[6] 尹林克.柽柳属植物的生态适应性与引种[J].干旱区研究,2002,19(3):12-16.
[7] 张道远,尹林克,潘伯荣.柽柳属植物抗旱性能研究及其应用潜力评价[J].中国沙漠,2003,23(3):252-256.
[8] 王雷涛,尹林克.准格尔盆地柽柳科植物生物多样性及保护对策[J].干旱区资源与环境,2004,18(6):139-145.
[9] 叶尔江·拜克吐尔汗,李海燕,朱小虎,等.北疆荒漠优势灌木种群空间异质性分析[J].干旱区资源与环境,2014,28(8):71-76.
[10] 丁新华,李铭,姜继元,等.塔里木河上游典型湿地植被群落特征[J].东北林业大学学报,2014,42(6):47-52.
[11] 俞言琳.新疆将启动林业十大重点工程[J].中国林业,2009,2B(4):26.
[12] 刘铭庭.柽柳属植物综合研究及大面积推广应用[M].兰州:兰州大学出版社,1995.
[13] 张道远,杨维康,潘泊荣,等.刚毛柽柳群落特征及其生态、生理适应性[J].中国沙漠,2003,23(4):446-450.
[14] 管海英,赵鑫,靳佳,等.荒漠生态系统土壤表层微生物量碳空间分布及其影响因子[J].干旱区研究,2014,31(6):1125-1131.
[15] Hultine K R,Bean D W,Dudley T L,et al.Species introductions and their cascading impacts on biotic interactions in desert riparian ecosystems[J].Integrative and Comparative Biology,2015,55(1):1-15.
[16] 解婷婷,苏培玺,周紫鹃,等.荒漠绿洲过渡带沙拐枣种群结构及动态特征[J].生态学报,2014,34(15):4272-4279.
[17] Liu Rentao,Zhu Fan,Steinberger Y.Effectiveness of afforested shrub plantation on ground-active arthropod communities and trophic structure in desertified regions[J].Catena,2015,125:1-9.
[18] 韩路.塔里木河上游荒漠河岸林植物群落动态与优势种群生态特征研究[D].北京:北京林业大学,2014.
