紫茎泽兰入侵对林分土壤可萌发种子库特征及生物多样性的影响1)
2015-03-08王有兵严毅王亚婷袁思安李子光周庆宏何承忠
王有兵 严毅 王亚婷 袁思安 李子光 周庆宏 何承忠
(昆明市海口林场,昆明,650114) (西南林业大学)
责任编辑:潘 华。
土壤中具有活力的种子总和称为土壤种子库[1]。在一定区域内土壤种子库是植被更新的重要组成部分,其物种的组成及质量会对植被更新有重大影响[2]。土壤种子库的种子主要来源为地上植被的种子雨,同时一些漂移或其它因素带来的种子形成的种子库又会影响到地上群落的组成及物种多样性的维持[3-4]。很多植物具有长久性的土壤种子库,这些种子在土壤环境受到破坏后,在适宜的条件下仍然具有活力[5-6]。
紫茎泽兰(Eupatorium adenophorum)属菊科泽兰属多年生草本植物[7]。由于紫茎泽兰具有较强的入侵性和破坏性,其对环境的适应性极强,加之种子量大、细小易通过自然或人为生产活动传播。目前紫茎泽兰已经成功入侵了桉树(Eucalyptus)人工林、青冈(Cyclobanopsis glauca)林、柏树(Cupressus funebris Endl.)与油茶(Camellia oleifera Abel.)混交林、油茶与青冈混交林、落叶松(Larix gmelinii)林和柏树林[8-10]。由于紫茎泽兰的入侵会形成林—紫茎泽兰复合系统,随着时间的推移会改变林下植被的组成,从而影响到林木的健康生长,降低了森林生态系统的稳定性,一旦该系统受到破坏,将难以恢复。为了探索滇中地区不同程度紫茎泽兰入侵林分土壤可萌发种子库的特征及生物多样性,为将来该地区人工林采伐后自然更新或自然灾害破坏后的更新预测提供理论依据,特别为在植被更新过程中可能出现的紫茎泽兰爆发性扩张防控提供理论指导。
1 研究区概况
研究地点位于昆明海口林场宽地坝林区(24°44'~24°54'N,102°29'~102°41'E),海拔1 941~1 975 m,坡度10°~15°。年均气温14.6 ℃,极端低、高温为-7.8 ℃和31.5 ℃。无霜期138 d 左右,有少量降雪,年降水量909.7 mm,干湿季分明,5—10月雨季降雨量占全年雨量的80%;旱季为11月至翌年4月。西北坡向,土层厚1.1 m,石灰岩母质,山地红壤,pH 值6.6。采样点林分分别为:桉树林(为1986年中国林科院与“澳大利亚国际农业中心”合作的澳大利亚桉树引种试验项目(8457)试验区)、侧柏(Platycladus orientalis (L.)Franco)林(1984年造林)、云南松(Pinus yunnanensis faranch)林(1984年造林)。
按树引种试验林设3 个样地,分别为第五(E1)、六(E2)、七(E3)小区,样地E1 有紫茎泽兰(Eupatorium adenophorum (Spreng.))、白 酒 草(Conyza japonica (Thunb.)Less.)、三叶鬼针草(Bidens pilosa L.)、臭灵丹(Laggera pterodonta)、千里光(Senecio scandens)、艾蒿(Artemisia argyi)、黄泡(Rubus pectinellus Maxim.)、悬钩子(Rubus corchorifolius L.F.)、青刺尖(Prinsepia utilis Royle.)、马桑(Coriaria sinica)、小叶荚蒾(Viburnum parvifolium Hayata)、荩草(Arthraxon hispidus (Thunb.)Makino)、野古草(Arundinella hirta (Thunb.)C.Tanaka)、山葡萄(Vitis amurensis)、木通(Akebia quinata)、铁仔(Myrsine africana L.)、芒种花(Hypericum patulum Thunb.)、大花卫矛(Euonymus grandiflorus Wall.)、清香木(Pistacia weinmannifolia)、刺槐(Robinia pseudoacacia L.)、金 粉 蕨(Onychium japonicum(Thunb.)Kunze)、毛 蕨 菜(Pteridium aquilinum(Linn.)Kuhn var.)、泥炭藓(Herba sphagni)、香薷(Elsholtzia ciliata (Thunb.)Hyland.)、毛茛(Ranunculus japonicus Thunb.)、苔 草(Cyperaceae tristachya)、酢浆草(Oxalis corniculata L.)共有27 种;样地E2 除黄泡、青刺尖、马桑、小叶荚蒾、荩草、山葡萄、木通、大花卫矛、毛蕨菜以外共有18 种;样地E3 除艾蒿、黄泡、青刺尖、马桑、荩草、山葡萄、木通、大花卫矛、清香木、金粉蕨、毛蕨菜、毛茛以外共有15 种;侧柏林设2 个样地,样地PO1 有悬钩子、西南栒子(Cotoneaster franchetii Bois)、野古草、铁仔、芒种花、毛叶杜鹃(Rhododendron championiae)、泥炭藓、香薷、苔草、酢浆草共10 种;样地PO2 特有沙针(Osyris wightiana Wall.Ex Wight),除此之外和其它样地相比少了三叶鬼针草、黄泡、青刺尖、荩草、山葡萄、木通、刺槐、金粉蕨、毛蕨菜,共有21 种;云南松林设1 个样地,样地PY 有臭灵丹、千里光、艾蒿、马桑、小叶荚蒾、野古草、铁仔、大花卫矛、泥炭藓、香薷、苔草、麦冬(Ophiopogon japonicus)、酢浆草共13种。
2 材料与方法
2.1 样地调查及土壤采集
每个样地设3 个20 m×20 m 的大样方,统计各林分主要乔木的密度,大样方四角及中心各设1 个2 m×2 m 的小样方,采用统计法[11]测量紫茎泽兰的盖度,统计小样方内的物种丰富度。采用直径5.1 cm 的环刀在每个小样方4 个角及中心分别采土壤,分0~5 cm 和>5~10 cm 2 层,每层采1 环刀土壤,每个大样方内同一层面的土壤进行混合备用。
2.2 土壤种子萌发及统计调查
试验于2014年1月5日采用萌发法进行。将采集的土壤放置于室内无风处自然风干,手工碾碎较大的土块,直到不能在碾碎为止,搓揉石块、根、枝、叶等杂物,轻轻敲打这些杂物后剔除。将初步处理好的土壤置于铺有一层塑料膜的育苗盘(40 cm×40 cm×5 cm)中,摊开土壤,厚度大约3~5 mm。采用质量浓度为10 mg/L 的GA3溶液将土壤喷洒透后置于温室大棚内自然光照下培养,后期采用自来水喷洒保湿,每间隔20 d 翻动土壤后喷洒GA3溶液一次进行催芽。每间隔20 d 分别统计鉴别小苗的类别和数量,计数单子叶植物和双子叶植物的数量,无法鉴别的采用育苗杯将其移植培养大后进行鉴别。种子萌发试验持续到无种子萌发为止。
2.3 数据统计处理
土壤中通过萌发检测到的可萌发种子总数除以土壤样本面积得到该土壤的可萌发种子库数量。计算以下指标[12-13]:土壤种子库的物种丰富度(S);物种多样性Shannon -Winener 指数H' = -∑(Ni/N)ln(Ni/N);群落优势度Simpson 指数D =1 -∑(Ni/N)2;群落均匀度Pielou 指数E=H'/lnS;群落结构组成差异Whittaker 指数β=S/ma-1;群落相似性Sorenson 指数Cs=2c/(a+b)。各式中:N 为群落中所有种的个体总和,Ni为群落中第i 种的个体数目,ma为群落的平均物种数量,c 为两个群落中共有物种数,a+b 为地表及土壤中所有物种数之和。
3 结果与分析
3.1 各样地地表植被调查
通过对各样地地表植被物种类型及紫茎泽兰盖度进行了调查(表1),所有样地中共含有31 种植物,隶属21 个科。其中E1 含18 科、E2 含13 科、E3 含11 科、PO1 含9 科、PO2 含15 科、PY 含10 科,菊科和蔷薇科植物较多。6 个样地中,物种丰富度最高的为E1,达27 种,但每个物种的个体数量极少;最低的为PO1,仅有10 种,地表几乎被泥炭鲜所覆盖。紫茎泽兰的盖度最高的为E1,达97.0%,由于桉树大面积死亡,为紫茎泽兰的入侵及扩张提供了条件,紫茎泽兰在该区域已经成为了林下单优势种群。各样地的物种丰富度与紫茎泽兰的盖度变化趋势基本一致,物种类型主要是一些多年生藤本植物、灌木及乔木。
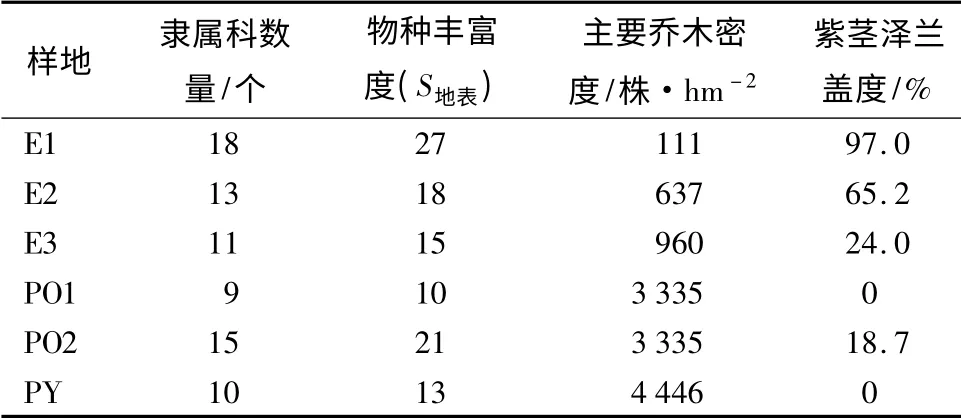
表1 样地信息
3.2 土壤可萌发种子库物种组成及密度
对不同林分土壤进行萌发检测了种子含量(表2),由表2可见,6 个样地土壤中检测到4~8 种植物,物种数最多的为桉树林(E2、E3),最少的为侧柏林(PO1、PO2)。除了样地E2 检测到相应林分乔木种子外,其余均未检测到。各样地土壤中以紫茎泽兰、香薷及野古草种子居多。土壤种子库种子主要分布在0~5 cm 的土层中,6 个样地的该层可萌发种子量占0~10 cm 土层种子量的比例为70.0%~91.2%,平均为83.4%,紫茎泽兰种子量为326~19 833 粒·m-2;>5~10 cm 土层可萌发种子量仅占0~10 cm 土层种子量的比例为8.8%~30.0%,平均为16.6%,紫茎泽兰种子量为0~6 448 粒·m-2。紫茎泽兰种子量在各样地中占比为16.7%~91.2%,平均为65.1%,即使在地表无紫茎泽兰生长的样地PO1 及PY 的土壤中,紫茎泽兰的种子量占比分别达76.7%和16.7%。PO1 样地紫茎泽兰种子占比较高,主要是因为该样地土壤种子库种子含量本身较低而导致的。种子密度最高的为样地E1,可萌发种子总数高达37 218 粒·m-2,最低的为样地PY,可萌发种子总数为1 959 粒·m-2。
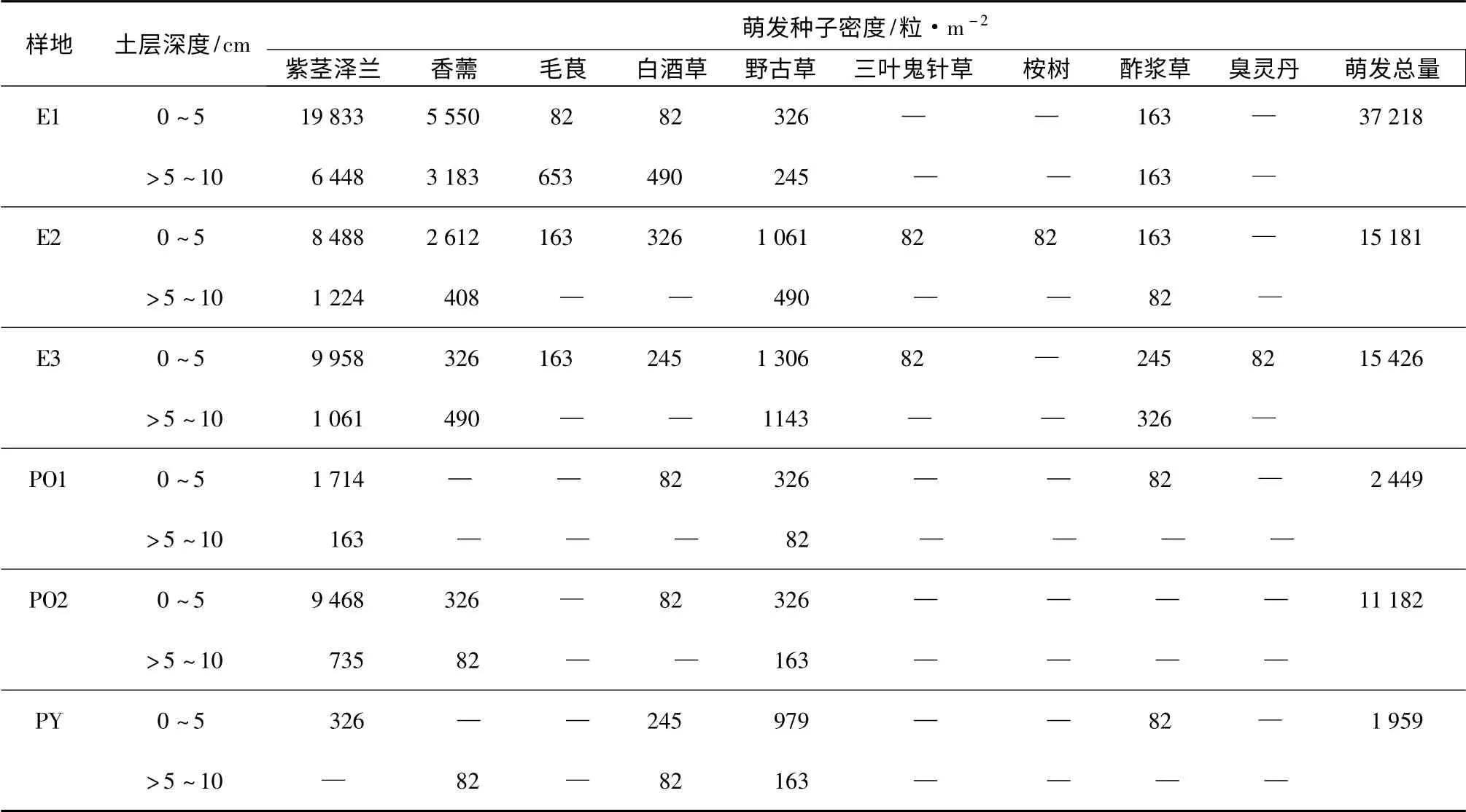
表2 各样地土壤可萌发种子库组成及密度
3.3 各样地土壤种子库种子的萌发特征
由表3可见,样地E1、E2、E3、PO1、PO2 土壤中种子的萌发时间主要集中前20~40 d,该时间段是种子的萌发高峰期,80 d 以后很少有种子萌发,样地PY 由于土壤中种子数量较少,种子的萌发高峰期也较其它样地靠后。该批检测到的土壤种子主要为双子叶植物,除样地PY 以外,其余样地土壤中双子叶植物占比达83.3%以上。
3.4 土壤种子库的群落特征
表4显示6 种林分土壤种子库的群落结构指标。从表4可知,紫茎泽兰入侵较重的桉树林地土壤种子库物种丰富度相对其它林分较高,平均在5.0~6.5 种之间;无紫茎泽兰入侵的侧柏林(PO1)土壤种子库物种丰富度最低,平均仅有3 种。从Shanon-Wiener 指数(H')看,物种多样性指数最高的为样地(PY),其次为(E2),再次为(E3),最小的为(PO2)。从Simpson 指数(D)看,样地(PY)的群落优势度最高,最低的为样地(PO2),这主要是与有无有害植物的入侵及林分密度有关。从Pielou 指数(E)看,指数最高的为样地(PY),其次是(PO1)。从群落结构组成差异Whittaker 指数(β)看,柏树林地(PO1、PO2)的群落结构组成差异指数较高,其余林分的指数较为接近,这和林分类型及地表植物有一定关系。从相似性指数(Cs)看,桉树林(E2、E3)的土壤种子库物种数与地被植物物种数的相似性较高,分别为0.62、0.70,相似性最低的为样地E1 和PO2,相似性指数分别为0.36、0.32。
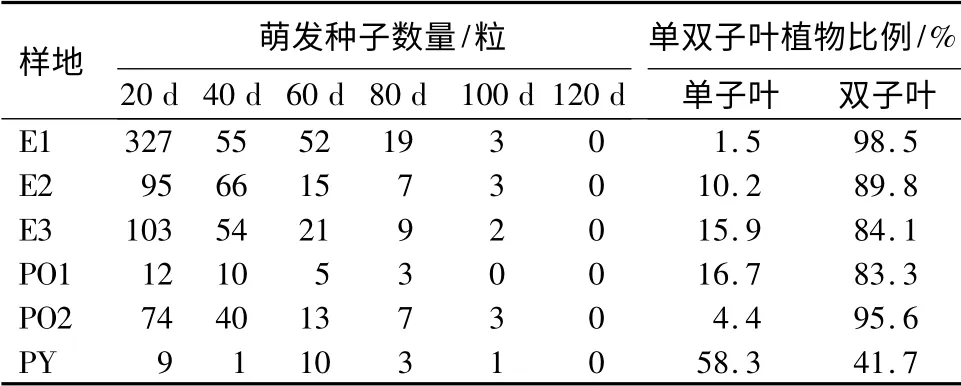
表3 不同样地土壤种子库种子萌发特征及单双子叶植物比例
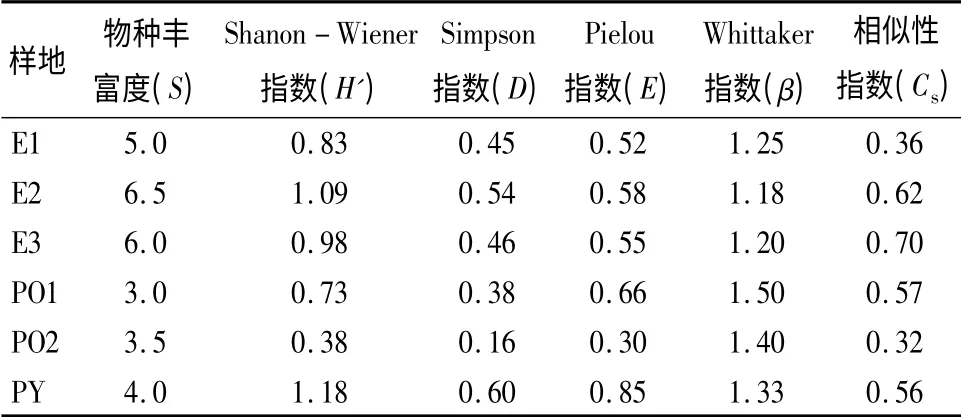
表4 各样地土壤中可萌发种子库的群落特征
4 结论与讨论
4.1 紫茎泽兰入侵对林下植被物种多样性的影响
入侵植物对自然生境土壤养分的正反馈作用促进了入侵,主要是因为入侵植物对入侵生境肥力平衡的打破而增加了其对本地植物的竞争能力[14]。土壤养分的提高通常有利于植物的生长竞争[15]。有研究表明,紫茎泽兰的入侵显著提高了根际土壤有机质、NH+4—N、NO-3—N、有效钾、土壤脲酶、磷酸酶和蔗糖酶的含量,而对土壤全氮含量没有明显影响[16-17]。王有兵等[10]对紫茎泽兰入侵桉树林地土壤含水率进行了研究,表明紫茎泽兰入侵后会提高土壤含水率,重度入侵区土壤含水率高达37.1%,几乎为轻度入侵区的2 倍。在6 个调查样地中,紫茎泽兰入侵过的林分,林下植被物种丰富度相对于未入侵林分较高,这可能与紫茎泽兰入侵后提高了土壤肥力及含水率有关,主要是一些较常见的草类、灌木及多年生藤本植物,这些生长在紫茎泽兰入侵地的草本植物耐阴性好,灌木高能够超过紫茎泽兰的高度,藤本植物可攀爬到紫茎泽兰或其它植物上而获得营养空间,但这些植物在样方中出现的频率并不高。
4.2 土壤可萌发种子库的萌发特征
种子库的密度是土壤种子库的重要特征之一[18]。根据土壤中种子活力的长久,可将土壤中的种子库分为长久性种子库和短暂性种子库两种[5]。试验中6 个样地共检测到9 种植物的种子,可萌发种子量处于1 959~37 218 粒·m-2,其中数量最多的为紫茎泽兰的种子,且主要集中在0~5 cm 的土层中,其数量可高达19 833 粒·m-2。所有样地的土壤种子量平均有83.4%分布在0~5 cm 的土层中,紫茎泽兰的种子量占总量的65.1%,在无紫茎泽兰入侵的林地土壤中也可检测到较高比例的紫茎泽兰种子。通过对土壤种子库种子的萌发,发现大部分种子的萌发高峰期主要在前20~40 d,并且双子叶植物居多,占萌发数量的83.3%以上,该研究结果与郭曼等[19]研究结果基本一致。沈有信等[6]对紫茎泽兰种子库进行了研究,表明紫茎泽兰具有长久性的种子库,它的种子至少能存活到新的萌发季节到来前。昆明地区自2012年冬季至今已经历过2 次冻害,2013年冬季甚为严重(-3.6 ℃),除房檐下及墙脚还有少量紫茎泽兰未受冻外,林下紫茎泽兰几乎全军覆没,至今已近2年未开花结实。本研究结果进一步说明紫茎泽兰存在长久性土壤种子库,在今后的林地更新过程中,需要对紫茎泽兰潜在的长久性土壤种子库可能出现的爆发性扩张采取一定的防控措施。
4.3 不同林分土壤种子库的物种多样性
土壤种子库是植物潜在的种群,其时空分布格局对植物群落的动态与稳定性有着举足轻重的影响[20]。有关杂草种子库的研究结果表明,杂草种子库潜群落优势种与地上部分杂草显群落优势种具有一致性[12],从试验样地土壤中紫茎泽兰的种子数量优势来看,其与地表紫茎泽兰的生长优势是一致的,并且紫茎泽兰种子的数量在总量中处于绝对优势。从土壤种子库可萌发种子种类来看,各样地的物种丰富度均不高,数量与地表物种种类相比少很多,这主要和各种植物的生殖特性及所处环境有关。有相关研究表明,植物群落与其土壤种子库的发育具有明显的不同步性[21],加之各样地的异质性导致了土壤种子库植物物种多样性、群落优势度、均匀度、群落结构组成差异性及与地表植物种类的相似性有较大的差别。如今在我国南方地区已形成了大面积的林—紫茎泽兰复合系统,该系统是否有利于当地生物多样性的稳定与发展,是否有利于人工林的健康生长及如何进一步防控紫茎泽兰值得深入研究。
[1] 张志权.土壤种子库[J].生态学杂志,1996,15(6):36 -42.
[2] Henderson C B,Petersen K E,Redak R A.Spatial and temporal patterens in the seed bank and vegetation of a desert grassland community[J].Journal of Ecology,1988,76(3):717 -728.
[3] 杨跃军,孙向阳,王保平.森林土壤种子库与天然更新[J].应用生态学报,2001,12(2):304 -308.
[4] 赵平,彭少麟.种、种的多样性及退化生态系统功能的恢复和维持研究[J].应用生态学报,2001,12(1):132 -136.
[5] Thompson K,Grime J P.Seasonal variation in the seed banks of herbaceous species in ten contrasting habitats[J].Journal of Ecology,1979,67(3):893 -921.
[6] 沈有信,刘文耀.长久性紫茎泽兰土壤种子库[J].植物生态学报,2004,28(6):768 -772.
[7] 吕霞,张汉波,张婷,等.紫茎泽兰根分泌物的化感潜力[J].云南大学学报:自然科学版,2008,30(3):314 -317.
[8] 黄娟,杨蕊.紫茎泽兰入侵对桉树人工林土壤的影响分析[J].绿色科技,2011(12):92 -95.
[9] 朱文达,曹坳程,颜冬冬,等.不同林木种群对紫茎泽兰营养生长和生殖生长的影响[J].生态环境学报,2013,22(11):1790 -1794.
[10] 王有兵,李子光,罗建中,等.紫茎泽兰对桉树苗木的化感作用[J].桉树科技,2014,31(1):27 -31.
[11] 何银忠,王有兵,严毅,等.滇中地区火烧迹地植被恢复研究[J].林业调查规划,2014,39(1):60 -64.
[12] 章超斌,马波,强胜.江苏省主要农田杂草种子库物种组成和多样性及其与环境因子的相关性分析[J].植物资源与环境学报,2012,21(1):1 -13.
[13] 万开元,潘俊峰,李儒海,等.长期施肥对旱地土壤杂草种子库生物多样性影响的研究[J].生态环境学报,2010,19(4):836-842.
[14] 蒋智林,刘万学,万方浩,等.紫茎泽兰与本地植物群落根际土壤酶活性和土壤肥力的差异[J].农业环境科学学报,2008,27(2):660 -664.
[15] Katharine N S,Julia R I,Eileen T,et al.Species effects on resource supply rates:do they influence competitive interactions[J].Plant Ecology,2004,175(1):47 -58.
[16] 蒋智林,刘万学,万方浩,等.紫茎泽兰入侵对土壤肥力特征的影响及其动态研究[J].农业环境科学学报,2008,27(1):267 -272.
[17] 李会娜,刘万学,戴莲,等.紫茎泽兰入侵对土壤微生物、酶活性及肥力的影响[J].中国农业科学,2009,42 (11):3964 -3971.
[18] 冯伟,潘根兴,强胜,等.长期不同施肥方式对稻油轮作田土壤杂草种子库多样性的影响[J].生物多样性,2006,14(6):461-469.
[19] 郭曼,郑粉莉,安韶山,等.黄土丘陵植被恢复过程中土壤种子库萌发数量特征及动态变化[J].干旱地区农业研究,2009,27(2):233 -238.
[20] 党伟光,高贤明,王瑾芳,等.紫茎泽兰入侵地区土壤种子库特征[J].生物多样性,2008,16(2):126 -132.
[21] 史鸿飞,张劲峰,李勇鹏,等.滇西北亚高山退化林地的土壤种子库[J].东北林业大学学报,2011,39(4):67 -70.
