柑橘抗寒相关基因及抗寒基因工程研究进展
2015-03-07张真真邱立军刘博文徐超红徐建国
张真真,邱立军,刘博文,徐超红,徐建国
(1.舟山市定海区林业工作站,浙江舟山 316000;2.浙江省柑橘研究所,浙江台州 318020)
柑橘抗寒相关基因及抗寒基因工程研究进展
张真真1,邱立军1,刘博文1,徐超红1,徐建国2∗
(1.舟山市定海区林业工作站,浙江舟山 316000;2.浙江省柑橘研究所,浙江台州 318020)
柑橘是我国南方重要经济树种,而低温是影响柑橘产量和分布的重要制约因素。基因差异表达分析技术是寻找植物抗寒基因的重要技术手段,而基因工程是获得植物抗寒品种的重要方法。本文综述了利用基因差异表达分析技术获得柑橘抗寒基因的方法,重点介绍了近年来在柑橘中已被分离鉴定的抗寒相关基因,分析了这些基因的抗寒功能及抗寒调控分子机制,并探讨了通过基因工程转化寒敏感柑橘的研究进展。
柑橘;低温诱导基因;抗寒基因;基因表达调控;基因工程
柑橘属(Citrus)是全球产量最大的水果,同时我国柑橘年产量位居全球第一[1]。柑橘是亚热带植物,喜温暖,对低温的反应敏感,所以低温是影响柑橘品质和产量的重要制约因素。同时低温影响柑橘的区域分布,限制柑橘这一重要经济植物在我国广大北方地区的栽培种植。因此,研究柑橘寒敏感原因,培育柑橘耐寒品种,对于提高柑橘品质和产量,打破柑橘地域,促进柑橘产业快速发展具有重大意义。
基因差异表达分析技术是寻找植物抗性基因的重要技术手段,而基因工程是获得植物抗性品种的重要方法,因此利用基因差异表达和基因工程技术能够获得柑橘抗寒相关基因,揭示柑橘寒敏感本质,有助于今后通过人工方法,调控柑橘抗寒过程,提高柑橘抗寒性,培育柑橘抗寒品种。迄今为止,已有80多篇涉及柑橘抗寒基因研究的文章发表,这些文章从柑橘属植物温州蜜柑、葡萄柚、甜橙中,分离鉴定了一部分低温差异表达基因,分析了这些基因与植物抗寒性之间的关系,并通过基因工程转化了部分柑橘类寒敏感品种,从而增加柑橘抗寒性。
1 柑橘抗寒相关基因获得方法
高等生物的基因表达不仅受到发育和组织特异性的影响,而且还受到环境变化的影响。任何细胞的基因90%是基础性表达,只有10%是选择性表达。而生物表现出来的各种特性,正是由于这10%基因的差异表达导致的,这里面包含了新基因的表达与表达量差异基因的表达。所以,分析低温诱导下不同基因的表达差异,并克隆这些差异表达的基因与模式植物比对了解其抗寒功能,对于深入研究柑橘抗寒分子机理,获得柑橘抗寒基因,转化柑橘寒敏感品种,提高柑橘抗寒性具有重要作用。
获得柑橘低温差异表达基因的主要技术有mRNA差异显示(mRNA differential display)、差别杂交(differential hybridization)、交互扣除RNA差别显示技术(reciprocal subtraction differential RNA display)、扣除杂交(subtractive hybridization of cDNA)、抑制消减杂交法(suppression subtractive hybridization)、定量(quantitative interpretation)PCR、半定量(semi⁃quantitative interpretation)PCR、电子消减(electronic subtraction)、基因表达系列(serial analysis of gene expression)分析和DNA微列阵分析(DNA microarray)[2]。
2 柑橘抗寒相关基因及其作用
Weiser[3]在1970年首先提出,植物在抗寒性诱导过程中基因表达发生了改变。随后利用各种基因差异表达分析技术,已在苜蓿、菠菜、冬小麦、马铃薯和冬油菜以及其他植物中,分离鉴定了多个与植物抗寒性相关的低温诱导基因[4],而柑橘植物的抗寒相关基因也在研究中越来越多地被分离鉴定出来。
在柑橘的抗寒性基因研究中,柑橘被认为是对低温比较敏感的植物,而同为芸香科柑橘亚科的枳是与柑橘亲缘关系很近的柑橘类植物,常用做柑橘砧木,对低温抗性好,耐寒[5],是研究柑橘抗寒性很好的比较材料。在柑橘抗寒能力的研究过程中,科学家通过各种差异表达分析法在低温胁迫的柑橘和枳中分离鉴定了一些低温诱导基因,而这些基因的功能大多通过与模式植物中已知功能的抗寒基因进行同源序列比对得知,并比较其抗寒性的差异,分析柑橘寒敏感分子机理,旨在获得能够提高柑橘抗寒性的基因。陈文荣等[6]利用mRNA差异显示技术分析鉴定寒敏感佛手低温胁迫相关的基因,最终获得121个低温差异片段,经过半定量和生物学分析,最终确定29个上调基因和5个下调基因,这些基因主要涉及植物防御/应激反应,信号转导,细胞壁修饰,转录,蛋白质的合成和氨基酸转运。张真真[2]研究了低温胁迫前后这些基因在佛手和枳的表达量变化情况,其中14个基因在佛手中的表达发生变化,但在枳中表达量却不变,这些基因可能与佛手寒敏感相关,它们所介导的分子途径可能是揭示佛手寒敏感特性的有利证据。近15年来,柑橘类植物低温胁迫后分离鉴定的低温诱导基因列于表1。
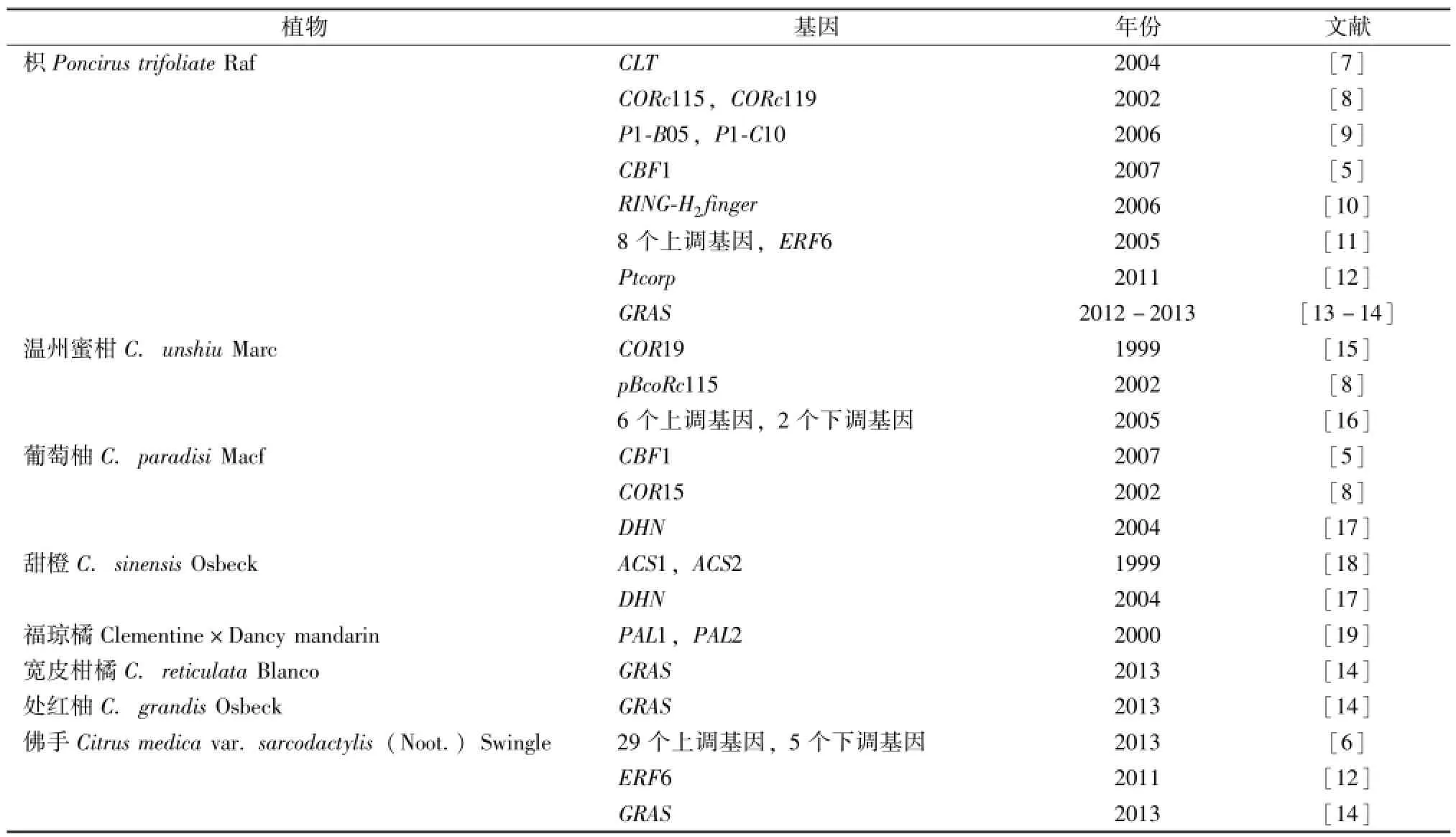
表1 近15年柑橘类植物中分离鉴定的低温诱导基因
在低温胁迫后,植物能够诱导体内表达某些基因,这些基因的表达蛋白直接参与低温胁迫应答或间接调控其他靶基因的表达[20]。陈得波等[21]归纳了自马铃薯、黑麦、小麦、菠菜、苜蓿和大麦以及其他植物里面分离鉴定的62个低温诱导基因,这些基因与植物抗寒能力有关系,而且这些基因的作用与膜脂相变、抗冻脱水、抗冻活性及低温下酶活性的变化有关。从表1中已经获得的柑橘类植物低温诱导基因,分析其合成的蛋白在抗寒中的作用可以分为3大类:1)编码具有催化作用的蛋白;2)促进新蛋白生成并附着细胞膜的表面或者镶嵌在膜脂之间,对于细胞膜的冷稳定性有促进作用;3)作为蛋白调节因子在信号转导和低温表达过程中起调节作用。下面对已经被分离鉴定的与柑橘低温诱导相关基因在抗寒方面的作用进行介绍。
2.1 酶活性蛋白
柑橘在低温胁迫过程中诱导合成具有催化作用的蛋白,参与一些重要的生理活动,如苯丙氨酸解氨酶(PAL)[19]、ACC合成酶[18]、硫氰酸酶[6]、甘油⁃3⁃磷酸酰基转移酶[22]、核苷二磷酸激酶Ⅲ[16]、氨基酸透酶[16]、酮还原酶[11]等。
苯丙氨酸解氨酶(PAL)是催化苯丙烷类代谢第一步反应的关键酶和限速酶。Sanchez⁃Ballesta等[19]从经低温胁迫下寒敏感的福琼橘(Fortune)和耐寒的赫南蒂娜克力迈丁红橘(Hernandina)中分离获得2个全序列PAL1和PAL2。研究发现低温胁迫后寒敏感柑橘的PAL表达量升高而耐寒柑橘的PAL表达量和PAL活性均下降,表明低温胁迫下PAL的表达量积累与柑橘受到低温伤害有关,PAL表达量的变化是区分柑橘品种抗寒性强弱的一种指标。
氨基环丙烷羧酸(ACC)合成酶基因ACS,是编码乙烯合成的关键酶,乙烯主要调节植物的生长发育及多种生理反应,如植物逆境胁迫反应。Wong等[18]从低温胁迫的甜橙中分离鉴定了2个低温诱导表达的ACC合成酶基因ACS1和ACS2,这2个基因不仅被低温诱导表达,同时也被伤害诱导表达。
硫氰酸酶具有植物组织解毒作用,能使细胞中有毒的氰化物,转变为无毒的硫氰酸化合物。陈文荣等[6]在寒敏感的柑橘植物佛手中,获得硫氰酸酶基因,这说明即使是抗寒性差的柑橘植物种,也同样会产生抗寒物质,用于对抗低温下氰化物积累对细胞造成的伤害。
Lang[16]和Zhang等[11]利用mRNA差异显示和定量PCR技术,从低温驯化的温州蜜柑中获得6个正调控低温诱导基因,其中2个基因编码具有催化作用的氨基酸透酶6和核苷二磷酸激酶Ⅲ。从低温驯化的枳中获得8个正调控基因,其中1个编码具催化作用的酮还原酶。
2.2 冷保护蛋白
低温伤害的主要部位是生物膜,研究表明低温胁迫过程中柑橘诱导新的蛋白质合成,附着于膜表面或位于膜脂间,具有保护细胞生物膜,维持水相,及防止蛋白变性等功能。如防止蛋白质受冻而失活的COR(cold⁃regulated)蛋白[8,15],保护细胞机能及抗脱水的LEA(late⁃embryogenesis abundant)蛋白[8],及一些与LEA蛋白、脱水蛋白类似的蛋白[23-25]。
在柑橘植物中报道最多的COR基因有COR11,COR15,COR19,这3个基因受低温、高盐、干旱等逆境诱导,能形成一类独特的LEA蛋白基因家族,均与柑橘的抗寒性相关。Porat等[8]从葡萄柚中获得COR15基因,编码一个15.1 ku的蛋白质COR15。Hara等[15]从低温驯化的温州蜜柑叶片中获得COR19基因,从枳中获得CORc115基因和CORc119基因,分别编码蛋白质COR19和COR11。这2种基因和温州蜜柑低温诱导表达的COR19,氨基酸序列都富含赖氨酸丰富区[24],且高度同源。温州蜜柑作为柑橘中抗寒性较强的品种,在低温胁迫下与抗寒性极强的枳均有COR19类似蛋白的表达形成,表明该基因可增加温州蜜柑的抗寒性。
另外Guy等[25]在低温驯化柑橘叶片里,观察到脱水蛋白CAP160和CAP85的累积,这体现出脱水蛋白参与了柑橘的抗寒过程。而且柑橘的CAP160与菠菜的CAP160无论分子量还是等电点都很相似,这体现了即使没有亲缘关系的植物同样也能生成一样的抗寒蛋白。2011年Porat等[17]在甜橙和葡萄柚热激后的低温诱导过程中获得脱水蛋白基因DHN,热激后冷藏时DHN的表达量明显增加。同时,发现葡萄柚在热激后放在常温下,被诱导增加表达的DHN是短暂的,而如果把热激后的葡萄柚进行冷藏,其诱导提高的DHN水平可以维持8周。Porat等[26]也曾有研究证实,柑橘冷藏前进行热处理可以提高柑橘储藏期间抗寒性。
2.3 植物的AFP抗冻蛋白
抗冻蛋白(antifreeze protein,AFP)首先发现于极地的鱼类中,它们与其他冷诱导蛋白的基本区别是:降低溶液冰点、抑制重结晶、修饰冰晶形态[27]。抗冻蛋白AFP已在冬黑麦、小麦、大麦、胡萝卜、燕麦、冬青、苔藓、裸子植物、细菌及真菌等生物中被发现[28-29],目前未见柑橘类植物获得AFP的报道。
2.4 转录因子
转录因子(transcription factor)指能结合基因上游特异核苷酸序列的蛋白质并起到调控基因转录的作用,转录因子与植物抗逆、发育及生命活动调控有重要联系。转录因子的调控作用研究一直是柑橘抗寒研究的热点,其中CBF(CRT/DRE binding factor)是目前柑橘中研究最深入的,参与低温逆境应答的转录因子。CBF是启动低温调节基因表达的重要转录因子,含有AP2结构,该结构域能与COR基因的CRT/DRE结合,启动COR蛋白,从而提高植物抗寒性[30]。2007年Champ等[5]从枳和葡萄柚中分离获得CBF,低温诱导下枳的CBF1比葡萄柚的CBF1积聚更多且达到更高的表达水平,而且枳的CBF1能够更迅速地识别并结合CRT/DRE片段,从而更早并更高水平地表达冷保护蛋白CORc115,从而增加枳的抗寒性。这项研究首次为2种亲缘关系相近但是抗寒性差距较大的柑橘植物,提供其抗寒差异与CBF表达(数量、时间)和低温胁迫程度相关性的证据。
乙烯应答因子(ethylene response factor,ERF)转录因子家族基因也受低温胁迫诱导表达,同样具有AP2结构,也在柑橘抗寒中起着重要作用。2011年曹诣斌等[12]通过RACE扩增从枳和佛手中获得ERF6,并使用定量PCR技术发现抗寒性不同的枳和佛手的ERF6低温胁迫下表达变化量完全不同,低温胁迫下佛手的ERF6表达量上升12倍,而枳的ERF6表达量上升4 000倍,说明ERF6表达量的变化与柑橘品种的抗寒性差异呈现正相关。2006年Sahin⁃Cevik等[9]通过扣除杂交法在枳的低温诱导cDNA文库中筛选获得Pl⁃B05和Pl⁃C10 2个基因,均含AP2/ERF结构域。在寒冷的不同时期对枳和葡萄柚的这2个基因表达量进行分析,发现这2个基因在枳中被诱导表达,葡萄柚中未被表达,说明ERF转录因子与柑橘抗寒性具有重要相关性。
GRAS是高等植物特有的转录因子家族,参与植物的生长发育、解毒作用、信号转导、生物和非生物胁迫的应答过程[31]。叶杰君等[14]2013年采用定量PCR,检测GRAS基因在低温胁迫的佛手、瓯柑、本地早、处红柚和枳,这5种抗寒性不同的柑橘中的表达量差异。这5种柑橘类植物半致死温度测定确定抗寒性强弱顺序是枳>本地早>瓯柑>处红柚>佛手,最终发现低温胁迫下GRAS基因在这5种柑橘中表达量均上调,并且表达量上调的幅度大小与植株的抗寒能力的强弱呈正相关。
3 柑橘抗寒基因工程
柑橘抗寒基因工程是指将外源抗寒基因导入到柑橘细胞,并培养成整个植株的过程。近15年来,国内外学者发现了大量植物低温诱导基因,并逐步探明低温胁迫的分子机理,柑橘抗寒基因工程取得了巨大的进步,通过导入植物抗寒基因进而提高柑橘抗寒性的方法已成为现代柑橘育种的重要手段。植物抗寒基因主要包括,与膜脂不饱和相关的酶基因,防止低温对细胞伤害的酶(SOD、脂脱氢酶)基因,抗冻及防止蛋白受冻失活的COR基因,抗脱水及保护胞体机能的LEA蛋白基因等。以下对柑橘中已有报道的抗寒基因工程途径做具体的介绍。
3.1 鱼类抗冻基因途径
抗冻蛋白(AFP)首先在鱼类中被发现,人们通过分离纯化获得高活性AFP,并筛选、克隆抗冻基因,以期能改良其他生物的抗冻性能,已有在昆虫和鱼类中获得AFP基因成功转化烟草和胡萝卜的报道[32-33]。万巧兰[34]利用含AFP基因的根癌农杆菌菌株感染锦橙、椪柑和枳的实生苗上胚轴部分,经卡那霉素筛选出抗性芽,并微嫁接成株,最后获得3株转基因柑橘(1株锦橙、2株枳)。通过Southern杂交和斑点杂交检测证实AFP基因已整合到柑橘基因组中。该研究结果初步表明,这种动物AFP能够在柑橘细胞中得到表达,但对柑橘抗寒性的影响尚待进一步研究。
3.2 脂肪酸去饱和代谢关键酶基因途径
植物的细胞膜是整个细胞中对外界胁迫反应最大的部位,所以细胞膜的组成成分与植物的抗寒性有着密切的关系。植物受到低温胁迫时不饱和脂肪酸含量会增加,同时质膜中磷脂含量及其与蛋白质的比值也会发生改变。有研究发现74种植物的抗寒性与反式单不饱和磷脂酸的含量密切相关[35],其不饱和度越高,抗寒性也就越强,因此在柑橘中导入脂肪酸去饱和酶基因,减少脂肪酸的饱和程度,能够大大提高柑橘的抗旱能力。甘霖等[36]研究证明抗寒性强的金柑和宜昌橙叶片的亚油酸、棕榈酸含量低,亚麻酸含量高,相对的脂肪酸不饱和度程度也高;然后抗寒能力比较差的哈姆林甜橙、伏令夏橙和粗柠檬却与此相反,由此可见脂肪酸不饱和程度与质膜的相变及流动性有着密切关系。孙中海等[37]同样认为柑橘质膜的不饱和程度与抗寒性呈正相关。
通过基因工程手段改变植物膜脂脂肪酸的组成,从而提高植物抗寒性的研究已有报道[23]。郭惠红等[22]将从枳中获得甘油⁃3⁃磷酸酰基转移酶(glycerol⁃3⁃phosphat⁃e⁃acyl⁃transferase,GPAT)基因导入温州蜜柑和甜橙中,以提高这2个柑橘品种的抗寒性,GPAT是磷酰甘油PG生成的关键酶,很大程度上决定了细胞膜PG的不饱和度。
3.3 外源抗氧化酶基因途径
低温胁迫下寒敏感植物细胞膜的伤害,可能与活性氧的升高有关。细胞原本就具有抗氧化能力,能够清除活性氧和自由基,其中包括超氧化物歧化酶(SOD)、过氧化物酶(POD)、谷胱甘肽还原酶(GR)、过氧化氢酶(CAT)等。细胞的抗氧化能力与柑橘的抗寒性呈正相关,因此细胞的氧化应激机制可能是调节柑橘抗寒性的另一重要机制。Sala等[38]发现柑橘品种间的抗旱性能的不同主要在于CAT,GR对过氧化氢的清除能力不同,经低温驯化后的柑橘,体内POD和SOD活性显著增强,叶片抗寒能力也显著提高。另一方面,有研究表明低温驯化提升了柑橘细胞体内抗氧化剂的含量和抗氧化酶活性,从而提高了柑橘的抗寒性。瞿金旺[39]利用根癌农杆菌介导法,将苹果中获得的亚精胺合成酶基因MSPDS1转入金钱橘,设想通过基因工程技术手段来提高柑橘内源多胺的含量,从而增强金钱橘抗寒性。研究结果表明低温胁迫下转入亚精胺合成酶基因MSPDS1的金钱橘叶片比对照植株叶片的相对电导率低,表现出更强的抗寒性。
4 前景展望
对柑橘抗寒基因的研究其最大意义在于通过基因工程方法提高一些寒敏感柑橘的抗寒能力。目前,已从鱼类抗冻基因途径、脂肪酸去饱和代谢关键酶基因途径、外源抗氧化酶基因途径这3个方面取得了一定的进展,其应用前景非常广阔[40]。柑橘作为重要的经济物种,在进行抗寒育种的同时应将基因工程技术手段与遗传育种、化学调控及科学栽培方法相结合,以最终提高柑橘抗寒性,防止冷害,打破柑橘种植地域限制,提高柑橘品质和产量。
[1] 郭卫东,张真真,蒋小韦,等.低温胁迫下佛手半致死温度测定和抗寒性分析[J].园艺学报,2009,36(1):81-86.
[2] 张真真.佛手低温胁迫相关基因的差异表达研究[D].金华:浙江师范大学,2009.
[3] Weiser C J.Cold resistance and injury in woody plants[J]. Science,1970,169:1269-1273.
[4] 沈漫,王明麻,黄敏仁.植物抗寒机理研究进展[J].植物学通报,1997,14(2):1-8.
[5] Champ K I,Febres V J,Moore G A.The role of CBF transcriptional activators in two citrus species(Poncirus and Citrus)with contrasting levels of freezing tolerance[J]. Physiologia Plantarum,2007,129(3):529-541.
[6] 陈文荣,叶杰君,李永强,等.佛手低温胁迫相关基因的差异表达[J].生态学报,2013,33(5):1594-1606.
[7] Jia Y,Rio H S,Robbins A L,et al.Cloning and sequence analysis of a low temperature⁃induced gene from trifoliate orange with unusual pre⁃mRNA processing[J].Plant Cell Reports,2004,23:159-166.
[8] Porat R,Pavoncello D,Lurie S,et al.Identification of a grapefruit cDNA belonging to a unique class of citrus dehydrins and characterization of its expression patterns under temperature stress conditions[J].Physiologia Plantarum,2002,115:598-603.
[9] Sahin⁃cevik M,Moore G A.Two AP2domain containing genes isolated from the cold⁃hardy citrus relative Poncirus trifoliate are induced in response to cold[J].Functional Plant Biology,2006,33(9):863-875.
[10] Sahin⁃cevik M,Moore G A.Isolation and characterization of a novel RING·H2 finger gene induced in response to cold and drought in the interfertile Citrus relative Poncirus trifoliata[J]. Physiologia Plantarum,2006,126(1):153-161.
[11] Zhang C K,Lang P,Dane F,et al.Cold acclimation induced genes of trifoliate orange(Poncirus trifoliata)[J].Plant Cell Reports,2005,23(10/11):764-769.
[12] 曹诣斌,石瑞,陈文荣,等.低温胁迫下佛手和枳乙烯应答因子6(ERF6)表达变化的比较分析[J].园艺学报,2011,38(10):1873-1882.
[13] Long G Y,Song JY,Deng Z N,et al.Ptcorp gene induced by cold stress was identified by proteomic analysis in leaves of Poncirus trifoliat a(L.)Raf[J].Molecular Biology Reports,2012,39(5):5859-5866.
[14] 叶杰君.柑橘类植物EF6和GRAS基因的低温胁迫应答及启动子序列分析[D].金华:浙江师范大学,2013.
[15] Hara M,Wakasugi Y,Ikoma Y.cDNA sequence and expression of a cold⁃responsive gene in Citrus unshiu[J]. Bioscience,Biotechnology,and Biochemistry,1999,63(2):433-437.
[16] Lang P,Zhang C K,Ebel R C,et al.Identification of cold acclimated genes in leavesof Citrusunshiu bymRNA differential display[J].Gene,2005,359:111-118.
[17] Porat R,Pasentsis K,Rozentzvieg D,et al.Isolation of a dehydrin cDNA from orange and grapefruit citrus fruit that is specifically induced by the combination of heat followed by chilling temperatures[J].Physiologia Plantarum,2004,120:256-264.
[18] Wong W S,Ning W,Xu P L,et al.Identification of two chilling⁃regulated 1⁃aminocyclopropane⁃1⁃carboxylate syntheses genes from citrus(Citrus sinensis Osbeck)fruit[J].Plant Molecular Biology,1999,41(5):587-600.
[19] Sanchez⁃ballesta M T,Lafuente M T,Zacarlas L,et al. Involvement of phenylalanine ammonialyase in the response of Fortunemandarin fruits to cold temperature[J].Physiologia Plantarum,2000,108:382-389.
[20] Sahin⁃cevik M.Identification and expression analysis of early cold⁃induced genes from cold⁃hardy Citrus relative Poncirus trifoliata(L.)Raf[J].Gene,2013,512(2):536-545.
[21] 陈得波,张爱平,姚泉洪.植物抗寒基因工程研究进展[J].生物技术通报,2001(4):14-20.
[22] 郭惠红,高述民,李凤兰,等.植物抗冻蛋白和抗寒基因表达的调控[J].植物生理学通讯,2003,39(6):555-561.
[23] Zhu L,Cai Z,Zhang Y,etal.Engineering stress tolerance ofEscherichia coli by stress⁃induced mutagenesis(SIM)⁃based adaptive evolution[J].Biotechnology Journal,2014,9(1):120-127.
[24] Sahin⁃cevik M,Moore G A.Cold⁃induced dehydrins from Poncirus trifoliata localized in the nucleus[J].Journal of Plant Biochemistry and Biotechnology,2012,21(1):134-139.
[25] Guy C L,Haskell P,Yelenosk Y G.Changes in freezing tolerance and polypeptide content of spinach and citrus at 5℃[J].Cryobiology,1988,25:264-271.
[26] Porat R,Pavoncello D,Peretz J,etal.Effectsof variousheat treatments on the induction of cold tolerance and on the postharvest qualities of‘Star Ruby’grapefruit[J].Postharvest Biology and Technology,2000,18(2000):159-165.
[27] 江勇,贾士荣,费云标.抗冻蛋白及其在植物抗冻生理中的作用[J].植物学报,1999,7(41):687-692.
[28] Griffith M,Ala P,Yang D S,et al.Antifreeze protein produced endogenously in winter rye leaves[J].Plant Physiology,1992,100:593-596.
[29] Dwnan JG,Olsen T M.Thermal hysteresis protein activity in bacteria,fungi,and phylogenetically diverse plants[J]. Cryobiology,1993,30:322-328.
[30] Hsieh T H,Lee JT,Yang P T,et al.Heterology expression of the Arabidopsis C⁃repeat/dehydration response element binding factor I gene confers elevated tolerance to chilling and oxidative stresses in transgenic tomato[J].Plant Physiology,2002,129(3):1086-1094.
[31] Li J,Yu C,Wu H,et al.Knockdown of a JmjC domain⁃containing gene JMJ524 confers altered gibberellin responses by transcriptional regulation of GRAS protein lacking the DELLA domain genes in tomato[J].Journal of Experimental Botany, 2015,66(5):1413-1426.
[32] Worrall D,Ellas L,Ashford D,et al.A carrot leucine rich repeat protein that inhibits ice recrystallization[J].Science,1998,282(5386):115-117.
[33] Kenward K D,Brandle J,Mcpherson J,et al.Type II fish antifreeze protein accumulation in transgenic tobacco does not confer frost resistance[J].Transgenic Research,1999,8(2):105-117.
[34] 万巧兰.柑橘离体形态建成及转鱼类抗冻蛋白基因研究[M].北京:中国农业科学院,1996:25-38.
[35] Roughan P G.Phosphatidylglycerol and chilling sensitivity in plant[J].Plant Physiology,1985,77:740-746.
[36] 甘霖,孙中海,邓秀新,等.柑橘体细胞杂种的抗性研究[J].园艺学报,1995,22(3):209-214.
[37] 孙中海,章文才,区胜祥,等.柑橘抗寒性与其膜脂脂肪酸组分的关系研究[J].武汉植物学研究,1990,8(1):79-86.
[38] Sala JM,Lafuente M T.Catalase in the heat⁃induced chilling tolerance of cold⁃stored hybrid Fortune Mandarin fruits[J]. Journal of Agricultural and Food Chemistry,1999,47:2410-2414.
[39] 瞿金旺.根癌农杆菌介导亚精胺合成酶基因MdSPDSl的柑橘遗传转化及离体再生[D].武汉:华中农业大学,2006:12-16.
[40] Zhang Z,Honda C,Kita M,et al.Structure and expression of spermidine synthase genes in apple:two cDNAsare spatially and developmentally regulated through alternative splicing[J]. Molecular Genetics and Genomics,2003,268(6):799-807.
(责任编辑:张 韵)
S 666
A
0528⁃9017(2015)11⁃1845⁃06
文献著录格式:张真真,邱立军,刘博文,等.柑橘抗寒相关基因及抗寒基因工程研究进展[J].浙江农业科学,2015,56(11):1845-1850.
DOI 10.16178/j.issn.0528⁃9017.20151146
2015⁃07⁃02
浙江省果品农业新品种选育(2012C12904);国家现代农业产业技术体系建设专项(CARS⁃27)
张真真(1983-),浙江舟山人,农艺师,硕士研究生,主要从事浙江省特色经济植物生物技术研究工作。E⁃mail:zhangzhenzhen1983@sina.com。
徐建国。E⁃mail:xujg@mail.zaas.ac.cn。
