模拟长江滩地水淹胁迫对3种树种幼苗生理生态特征的影响1)
2015-03-06王晓荣胡兴宜唐万鹏庞宏东周少辉
王晓荣 胡兴宜 唐万鹏 庞宏东 周少辉
(湖北省林业科学研究院,武汉,430075) (衡阳市林业科学研究所)
责任编辑:任 俐。
水淹是植物主要非生物胁迫之一,它可以发生在江河洲滩、盐沼、沼泽等多个生态系统[1],每年在沿海、沿江地带都有可能发生的洪涝灾害,往往导致生长于该区域的植物形态结构发生变化,甚至死亡,对我国的农业生产和林业生态建设产生了巨大的影响[2-3]。因此,研究植物应对水淹胁迫下的生理生态响应过程以及机理,已经成为众多国内外学者关注的热点[4-6]。
长江中下游滩地是平原河床季节性淹水湿地,面积约为6.3×105hm2,每年汛期暴雨和洪水均对此区域造成洪涝灾害,形成冬陆夏水,类型复杂,具有独特的季节性淹水特征[7],植物为适应水淹环境会产生一系列复杂的生理代谢变化[8],这对植物能否在水淹胁迫下具有正常的生理功能和进行生长发育至关重要[2]。水淹会导致土壤中氧气和光照的不足,其中氧气的不足主要是因为在水分饱和状态下气体扩散放慢,使得植物必须要在低氧和低光照条件的生态环境中生长[9],从而影响其呼吸作用、光合作用、氧化还原稳态和胞质的pH值等生化活动[10]。已有研究表明,对于不耐淹的树种来说,无氧的环境抑制了根的有氧呼吸并且导致根活力降低,淹水导致植物体内的抗氧化酶活性、激素水平、可溶性物质质量分数的改变[11]、叶绿素降解、膜渗透性增加、气孔关闭,光合速率降低以及叶片的衰老凋落[5,9]。然而,耐淹植物通过形态结构变化,如形成肥大的皮孔、不定根的形成[1,12],以及相应的生理适应机制变化,如代谢途径转为无氧酵解途径、激素关系的改变等,进而适应缺氧的环境[13-14]。
长江中下游滩地是血吸虫病重疫区,枫杨(Pterocarya stenoptera)、夹竹桃(Nerium indicum)、乌桕(Sapium sebiferum)是营建抑螺防病林的重要树种,不仅具有良好的抑螺作用,也比较能适应水淹环境,在改善区域抑螺防病、水源涵养和美化景观过程中具有重要的作用。许多学者也针对其淹水胁迫下的生理生态影响进行模拟研究[1,15],但均以单一树种进行模拟研究,而且设计的淹水高度以及淹水时间与实际情况有所差异。以多种适宜树种并结合长江滩地季节性水淹特点的研究还未见报道。本研究以枫杨、乌桕和夹竹桃为研究对象,模拟了长江滩地70 d的间歇性水淹过程,研究水淹胁迫对此3种树种的生长、生物量分配、光合响应、以及叶绿素质量分数的影响,揭示不同树种对水淹胁迫的适应性和差异性,从而为长江滩地治理开发、抑螺防病林和防护林建设提供一定的科学依据。
1 材料与方法
挖取湖北省黄冈长江滩地土壤,运至湖北省九峰森林国家公园试验基地淹水胁迫试验池,作为模拟试验的土壤。2012年3月份,将枫杨、夹竹桃、乌桕树苗按照40 cm×60 m的株行距种植在2 m×3 m的试验池中,每个试验池每个树种各5株,4个水池共计20株。为保证苗木的相对一致性,于2013年7月份淹水试验前对其进行修枝处理,使树高基本一致。由于滩地多属阶段性积水,水流、水位变化大,水淹造成不同高程滩地植被影响存在差异。因此,依据长江滩地淹水时间和水位变化情况,试验周期为70 d,从7月10日始至9月20日止。本试验4个处理水平分别为:CK组为对照组(日常浇水管理);T1组为长期浅深度水淹(水面在距土壤表面10 cm),模拟轻度水淹胁迫;T2组为短期浅深度水淹(水面在距土壤表面10 cm,水淹1个月后退水),模拟轻度水淹胁迫的干湿交替;T3组为长期深度水淹(水面在距土壤表面1 m),模拟重度水淹胁迫。定期更换水池中的水,每日观察水位情况,以保证水位高度正常。
株高、地径的测定:定期观察试验苗木生长形态变化特征,如叶片发黄脱落、不定根形成以及皮孔出现等现象,进行详细记录。株高用卷尺测量(精确到0.1 cm),地径用游标卡尺交叉测定2次(精确到0.01 mm),取平均值,于淹水前和试验结束各测量1次。生长期不同处理植株的株高、地径相对生长率按以下公式计算,R=((H2-H1)/H1)×100%,其中H1和H2分别表示水淹前和水淹结束两次测量时的株高和地径。
生物量的测定:于2013年9月份试验末期,应用全收割法,将幼苗整棵挖出清洗干净后,按叶、枝、茎、根进行鲜质量称量,之后放置在烘箱中80℃烘干至恒质量,分别称其干质量,计算叶、枝、茎、根生物量以及根冠比(根干质量/地上部分干质量)。其中枫杨在重度水淹处理50 d时已全部死亡,叶子全部脱落,导致叶生物量数据未采集。
叶绿素质量分数测定:于2013年8月底,采用乙醇浸泡法进行叶绿素测定。取0.2 g新鲜叶片剪碎,长度2 mm左右,然后放入50 mL试管容量瓶中,用95%乙醇定容至刻度,避光浸提24 h,且每隔2 h震荡1次直至叶片完全发白为止,然后利用分光光度计在波长665和649测定吸光度,重复3次,计算出相应叶绿素质量分数[16]。
光合作用的测定:于2013年9月初,利用LI-6400XT便携式光合测定系统,选择晴天09:00—12:00进行植株的光响应测定。每个树种随机选3株幼苗,选择中上部第6~10片叶片,利用CO2注入系统控制CO2质量分数为400μg·g-1,温度控制在25~30℃,相对湿度55%左右,采用Li-6400红蓝光源做内置光源,设定光合有效辐射梯度为(PA,R):2 000、1 500、1 200、1 000、800、600、400、200、150、100、75、50、25、10和0μmol·m-2·s-1,测定其净光合速率。
光响应曲线的相关参数利用叶子飘等[17-18]研究的直角双曲线修正模型求得,模型函数表达式为:Pn=α(1-βI/1+γ)I-Rd,式中,Pn为净光合速率;α是光响应曲线的初始斜率,即表观光合量子效率;β和γ为系数;I为光合有效辐射;Rd为暗呼吸速率。植物的光饱和点(LS,P)和最大净光合速率(Pmax)分别由和求得。
数据处理:数据统计均采用SPSS 16.0和Excel 2007软件进行处理和分析,根据各项测定指标,将水淹处理水平作为独立因素,以采用单因素方差分析(one-way ANOVA),且进行多重LSD比较,以检验数据之间的差异显著性(P=0.05);采用Sigma-Plot 10.0作图。
2 结果与分析
2.1 水淹胁迫对3种树种幼苗生长特征的影响
试验期间,水淹除了导致枫杨在长期重度水淹50 d死亡外,其他处理的树种存活率达到100%。由表1可知,水淹胁迫对3种树种幼苗地径和株高均产生了明显的影响。枫杨和夹竹桃地径相对生长由大到小的顺序均表现为T2、CK、T1、T3,乌桕由大到小的顺序为T2、CK、T3、T1,说明短期轻度水淹在一定程度上促进了植物地径的生长,而长期轻度水淹和重度水淹均对地径生长具有显著的抑制作用。株高生长变化趋势各物种有所差异,分别为枫杨由大到小的顺序是CK、T2、T1、T3,夹竹桃为CK、T1、T2、T3,说明水淹均在一定程度上抑制了此两种树种株高的生长(P<0.05),而乌桕则为T2、CK、T1、T3,退水后甚至超过了对照组。

表1 水淹胁迫对3种树种幼苗生长的影响
2.2 水淹胁迫对3种幼苗生物量分配的影响
如表2所示,不同水淹胁迫处理对不同树种生物量累积影响变化各异,水淹明显抑制了3种树种生物量的累积。其中,夹竹桃、乌桕均表现为重度水淹胁迫显著降低了总生物量的积累(P<0.05),其T3组分别较对照组降低42.18%和33.90%,T2组与对照组未表现出显著差异(P>0.05),但仍可发现轻度水淹对植株正常生理生物量积累产生了一定的抑制作用,而退水后植株又能够通过快速生长逐渐得到恢复,从而提升了一定的生物量。从生物量的分配格局来看,3种树种的茎生物量均未受到明显影响,而叶、枝和根生物量都受到不同程度的影响,说明植物的叶、枝和根器官受到胁迫后感应最为敏感。对比植物根冠比发现,重度水淹导致夹竹桃和乌桕树种根冠比较对照具有显著的降低(P<0.05),分别降低了41.66%和37.82%,而轻度水淹对3种幼苗根冠比均未造成显著影响(P>0.05),但从中也可看出短期轻度水淹致使植物根冠比具有增加的趋势,这可能与退水导致根部生长活性不断恢复有关,特别是夹竹桃根部生物量累积超过了对照组,说明其对季节性水淹适应的能力更强。
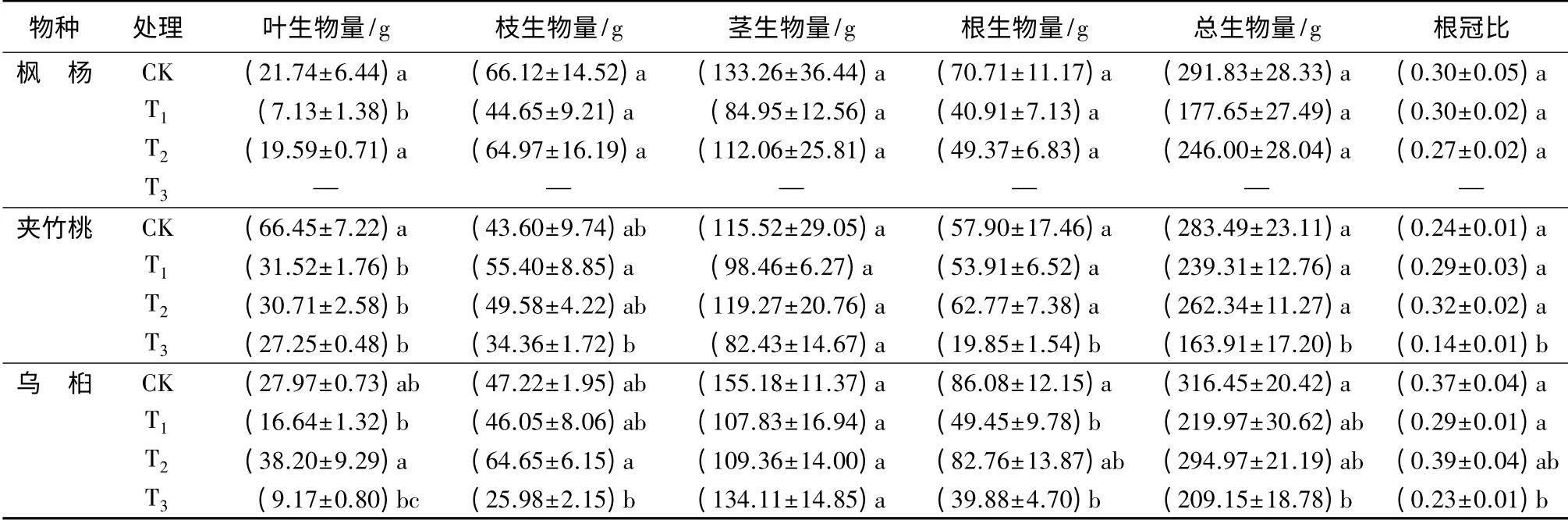
表2 水淹胁迫对3种树种幼苗生物量分配的影响
2.3 水淹胁迫对3种树种幼苗光合作用的影响
从表3可知,不同水淹胁迫处理对3种树种最大净光合速率Pmax均具有显著影响(P<0.05),夹竹桃由大到小的顺序表现为T2、CK、T1、T3,枫杨由大到小的顺序表现为T2、CK、T1,乌桕由大到小的顺序表现为CK、T2、T1、T3,表明长期水淹胁迫不利于光合干物质的积累,而短期轻度胁迫则在一定程度上促使光合能力不断恢复。本研究中3种树种在不同水淹处理下,LC,P、α、Rd与对照均无显著性差异(P>0.05),表明三者在水淹后光能利用效率及利用弱光进行物质生产的能力并没有受到水淹的明显影响,但仍可看出暗呼吸速率呈现明显增加的趋势。水淹胁迫导致枫杨和乌桕的LS,P与对照存在显著性差异(P<0.05),均表现为显著性降低,可见两者对强光的耐受性发生明显降低。同时,由于水淹胁迫对夹竹桃的α、LC,P、LS,P、Rd均未因水淹处理的不同而与对照存在显著差异,可见该物种并未因水淹胁迫而光合生理受到太大变化,属于耐水淹较强的树种。

表3 水淹胁迫下3种树种幼苗光响应变化
2.4 水淹胁迫对3种幼苗叶绿素质量分数的影响
如表4所示,长期轻度水淹和重度水淹导致3种树种幼苗叶绿素a和叶绿素(a+b)质量分数显著降低,而轻度水淹退水后又明显上升。叶绿素b则除乌桕外,均表现出相对稳定的态势,水淹胁迫导致乌桕叶绿素b分别降低44.28%、32.47%和43.14%,但短期轻度水淹后植物叶绿素a和叶绿素b均恢复正常水平。对比不同物种在不同水淹处理下的叶绿素a/叶绿素b发现,长期水淹胁迫会导致枫杨和夹竹桃叶绿素a/叶绿素b显著降低(P<0.05),而乌桕则无显著变化(P>0.05)。

表4 水淹胁迫对3种幼苗叶绿素质量分数的影响
3 结论与讨论
植物的生物量、株高和基径与其生长发育、营养物质的形成密切相关,对其所处的生长环境的综合表征作用明显[19]。大量研究表明,长期淹水后植株虽然会在形态结构上产生一定的适应性,但会显著减少植物的生长,甚至导致植株死亡[4]。本研究中,除长期重度水淹导致枫杨发生死亡外,其他物种均存活良好,表明了3个物种具有很好的耐水淹特性。曹福亮等[20]研究认为,淹水造成幼苗高生长降低的直接影响是阻碍节间的生长,间接作用是通过阻止叶原基分化、叶片生长以及诱导叶衰老、脱落等造成的,但退水胁迫的解除,反而刺激了其后期的株高生长。然而,长期水淹胁迫对植物地径和株高具有显著的抑制作用,而短期轻度水淹在一定程度上促进了3种植物地径的生长,这可能与通气组织的形成有关,从而促进氧气从植株地上部分向地下部分扩散以保证根系正常的呼吸代谢[15],本试验在乌桕和夹竹桃淹水过程中,发现其长出大量的不定根。同时,水淹明显抑制了3种树种生物量的累积,但退水后植株均能够通过快速生长逐渐得到恢复,从而提升了一定的生物量。长期重度水淹导致夹竹桃和乌桕树种根冠比较对照具有显著的降低,分别降低了41.66%和37.82%,这与曹福亮等[1]等研究结果一致,而长期轻度水淹对3种幼苗根冠比均未发生显著影响,短期轻度水淹致使植物根冠比具有增加的趋势,表明3种树种对季节性水淹的适应性。
已有研究表明,最大净光合速率、表观量子效率、光饱和点、光补偿点等光响应参数是研究植物光合生理特性以及适应环境变化能力的重要指标[21]。在水淹条件下,植物保持较高的光合速率和正常的光合特性是植物能耐受水淹的重要原因之一[5],如果某植物个体具有较高的最大净光合速率、光饱和点、表观量子产额和较低的光补偿点,说明该个体具有较高的光能利用效率,喜光但也适应在低光照条件下生长,具有更好的光环境适应性与生物生产潜力[22-23]。本研究中,长期水淹均导致最大净光合速率Pmax降低,而短期轻度胁迫又光合能力不断恢复,这与大部分的研究结论相一致[14,24]。LC,P、α以及Rd与对照均无显著性差异,说明3种树种对弱光的利用能力维持较高水平,从而尽可能的维持光合干物质的生产。枫杨和乌桕的LS,P对水淹胁迫均表现为显著降低,可见两者对强光的耐受性明显降低,而夹竹桃没有显著变化,进一步反映出夹竹桃适应深水淹环境生长的优良特性。
光合色素是植物叶片光合作用的物质基础,其质量分数的高低能反映植物的生长状况和叶片光合能力。水淹胁迫下,大部分植物叶绿素质量分数下降,光合作用减弱[25]。本研究发现,长期轻度水淹和重度水淹导致3种树种幼苗叶绿素a、叶绿素(a+b)质量分数显著降低,而轻度水淹退水后又明显上升,这与耐淹水植物的光合动态特征一致[15]。另外,长期水淹胁迫会导致枫杨和夹竹桃叶绿素a/叶绿素b显著降低,这暗示淹水胁迫导致枫杨和夹竹桃叶片光合反应中心的降解程度要高于捕光蛋白色素复合物的降解[11]。但乌桕则无显著变化,且呈现上升的趋势,表明受到水淹影响后,乌桕通过调节叶片叶绿素a/叶绿素b来维持其较高的光合能力。
根据本水淹试验的观察发现,夹竹桃和乌桕均在与水接触的茎部产生大量的不定根来适应淹水环境,特别是在较长时间的淹水环境中,不定根的产生有利于植物的水分、养分及激素的供应和调节[3],呈现出较强的耐受能力,存活率达到100%,而枫杨由于未见外部通气组织的发生,不适应长期重度水淹胁迫环境,全部发生死亡现象。另外,短期轻度水淹胁迫退水后,3种植物快速恢复其生理活动,生长和光合生理特征逐渐恢复正常水平或维持在较为稳定的状态。陈芳清等[26]研究认为,退水后迅速恢复生长活力的植物,一方面表明了该物种对水淹具有较强的适应能力,另一方面反映了植物的“饥饿”效应。植物受水淹胁迫过程中,体内的可溶性糖质量分数急剧上升,因维持其生命力而消耗了大量的营养物质,植株处于“饥饿”状态,只有通过快速地生长来弥补营养不足。从模拟长江不同水淹程度来看,夹竹桃、乌桕和枫杨均具有较强的耐水淹特性,但淹水时间不宜过长,且枫杨的淹水深度不宜过深,均更适宜在季节性浅度水淹的长江滩地生长。
[1]曹福亮,蔡金峰,汪贵斌,等.淹水胁迫对乌桕生长及光合作用的影响[J].林业科学,2010,46(10):57-61.
[2]刘泽斌,程瑞梅,肖文发,等.水淹胁迫对植物光合生理生态的影响[J].世界林业研究,2013,26(3):33-38.
[3]赵竑绯,赵阳,张驰,等.模拟淹水对杞柳生长和光合特性的影响[J].生态学报,2013,33(3):898-906.
[4]Mielke M S,Almeida A F,Gomes F P,et al.Leaf gas exchange,chlorophyll fluorescence and growth responses of Genipa amsricana seedlings to soil flooding[J].Environment and Experimental Botany,2003,50(3):221-231.
[5]Chen Hongjun,Quails R G,Blank R R.Effect of soil flooding on photosynthesis,carbohydrate partitioning and nutrient uptake in the invasive exotic Lepidium latifolium[J].Aquatic Botany,2005,82(4):250-268.
[6]Mommer L,Lenssen JP M,Huber H,et al.Ecophysiological determinants of plant performance under flooding:a comparative study of seven plant families[J].Jourhal of Ecology,2006,94(6):1117-1129.
[7]彭镇华,江泽慧.中国新林种抑螺防病林研究[M].北京:中国林业出版社,1995:66-73.
[8]Parolin P.Morphological and physiological adjustments to waterlogging and drought in seedlings of Amazonian floodplain trees[J].Oecologia,2001,128(3):326-335.
[9]陈芳清,李永,郄光武.水蓼对模拟水淹的生理生态学响应[J].生态环境,2008,17(3):1096-1099.
[10]谭淑端,朱明勇,张克荣,等.植物对水淹胁迫的影响与适应[J].生态学杂志,2009,28(9):1871-1877.
[11]薛艳红,陈芳清,樊大勇,等.宜昌黄杨对夏季淹水的生理生态学响应[J].生物多样性,2007,15(5):542-547.
[12]罗芳丽,曾波,陈婷,等.三峡库区岸生植物秋华柳对水淹的光合和生长响应[J].植物生态学报,2007,31(5):910-918.
[13]McManmon M,Crawford R M M.A metabolic theory flooding tolerance:the significance of enzyme of distribution and behavior[J].New Phytologist,1971,70(2):299-307.
[14]衣冠英,樊大勇,谢宗强,等.模拟水淹对池杉和栓皮栎光合生理生态过程的影响[J].生态学报,2008,28(12):6025-6033.
[15]衣冠英,樊大勇,谢宗强,等.模拟水淹对枫杨和栓皮栎气体交换、叶绿素荧光和水势的影响[J].植物生态学报,2006,30(6):960-968.
[16]李志丹,韩瑞宏,廖桂兰,等.植物叶片中叶绿素提取方法的比较研究[J].广东第二师范学院学报,2011,31(3):80-83.
[17]Ye Zipiao.A new model for relationship between irradiance and the rate of photosynthesis in Oryza sativa[J].Photosynthetica,2007,45(4):637-640.
[18]叶子飘,赵则海.遮光对三叶鬼针草光合作用和叶绿素含量的影响[J].生态学杂志,2009,28(1):19-22.
[19]张晔,李昌晓.水淹与干旱交替胁迫对湿地松幼苗光合与生长的影响[J].林业科学,2011,47(12):158-164.
[20]曹福亮,罗伯特·法门.人工淹水胁迫处理对美洲黑杨无性系苗形态和生长的影响[J].南京林业大学学报,1993,2(17):39-44.
[21]王明怀,许秀玉,魏龙,等.木麻黄无性系光响应特性研究[J].华南农业大学学报,2011,32(2):76-79.
[22]钟全林,胡松竹,贺利中,等.刨花楠不同种源主要光响应指标分析[J].林业科学,2008,44(7):118-123.
[23]伍维模,李志军,罗青红,等.土壤水分胁迫对胡杨、灰叶胡杨光合作用-光响应特性的影响[J].林业科学,2007,43(5):30-35.[24]吴统贵,袁涛,王宗星,等.两种消浪树种幼苗光合特性对模拟潮汐水淹的响应[J].植物研究,2012,32(5):615-620.
[25]王宗星,吴统贵,虞木奎.植物光合特性对水淹胁迫的响应机制[J].世界林业研究,2010,23(5):20-24.
[26]陈芳清,黄友珍,樊大勇,等.水淹对狗牙根营养繁殖植株的生理生态学效应[J].广西植物,2010,30(4):488-492.
