MiRNAs在苯丙胺类兴奋剂成瘾中作用的研究进展
2015-02-26江明金林莹波朱道琦莫志贤
江明金,李 婵,林莹波,朱道琦,莫志贤
(南方医科大学中医药学院中药药理教研室,广东广州 510515)
MiRNAs在苯丙胺类兴奋剂成瘾中作用的研究进展
江明金,李 婵,林莹波,朱道琦,莫志贤
(南方医科大学中医药学院中药药理教研室,广东广州 510515)
中国图书分类号:R-05;R342.2;R749.61;R971.7;R971.93;R977.9
摘要:苯丙胺类兴奋剂(amphetamine-type stimulants,ATS)是一组以精神依赖为主的新型合成毒品,近年来流行,滥用趋势愈发严峻。MicroRNAs(MiRNAs)作为一类非编码小分子RNAs,通过与靶基因mRNA的互补配对,在转录后水平上对基因的表达进行负调控,从而导致靶基因mRNA的降解或翻译抑制。ATS能诱导miRNAs表达水平的变化,而成瘾相关脑区miRNAs表达的改变直接参与了对ATS成瘾行为的调节。因此,研究miRNAs在ATS成瘾中的调控作用,对进一步揭示新型毒品的成瘾机制及发现新的药物作用靶点具有重要意义。
关键词:miRNAs;苯丙胺类兴奋剂;苯丙胺;甲基苯丙胺;成瘾;表达;调控;机制
网络出版时间:2015-9-14 14:53 网络出版地址:http://www.cnki.net/kcms/detail/34.1086.R.20150914.1453.012.html
药物成瘾又称药物依赖,是大脑在持续的成瘾物质作用下产生的慢性复发性脑病[1]。成瘾物质的长期反复使用会造成机体中枢神经系统的可塑性变化,表现为大脑相关核团(包括中脑腹侧被盖区、伏隔核、杏仁核、海马、前额叶皮质、下丘脑、尾状壳核等)神经元突触、神经环路的细胞形态及功能的可塑性变化,进而导致成瘾的一系列行为症状[1-2]。药物成瘾的本质是一种以药物引起的基因表达和神经突触可塑性改变为基础的病理性记忆[2]。苯丙胺类物质是一组具有类似化学结构的中枢神经系统兴奋剂,包括苯丙胺、甲基苯丙胺(俗称“冰毒”)、3,4-亚甲基二氧基甲基苯丙胺(俗称“摇头丸”)等其他苯丙胺类精神兴奋剂。苯丙胺类兴奋剂(amphetamine-type stimulants,ATS)是目前最为流行的一类新型合成毒品,滥用增长势头迅猛。ATS具有精神依赖性强、复吸率高和神经毒性大等特点,可造成滥用者身心严重损害,并且可导致由共用注射器和性行为造成的艾滋病病毒(human immunodeficiency virus,HIV)感染、传播,使滥用者形成暴力倾向,给社会治安带来严重的负面影响[3]。
MicroRNAs(MiRNAs)是一类长度约18-25个核苷酸的内源性非编码小分子RNAs。MiRNA基因首先转录合成初始miRNA(pri-miRNA),再经过剪切得到miRNA前体(pre-miRNA),在转运蛋白转运下从细胞核转运到细胞质,最后通过Dicer酶剪切形成成熟的miRNA。成熟的miRNA通过与AGO蛋白形成RNA诱导沉默复合物,经过碱基互补配对原则特异性地与靶基因mRNA结合。当miRNA和靶基因mR-NA碱基对不完全互补结合,则抑制翻译过程,进而调节蛋白表达水平;当miRNA和靶基因mRNA碱基对完全互补结合,则切割和降解靶基因mRNA,导致基因沉默。MiRNAs已被发现存在于哺乳动物神经系统中并且部分在脑部富集,对中枢神经系统发育、神经元分化和突触可塑性发挥重要的调控作用[4-5]。关于miRNAs参与药物成瘾调控的研究,自2008年,Pietrzykowski等[6]首次报道后迅速增多,成为研究的热点之一。目前,对于miRNAs与可卡因、酒精、尼古丁及阿片类成瘾性物质的研究报道较多,而对miRNAs在ATS成瘾中的调控作用所知甚少,处于起步阶段。本文主要针对miR-NAs与ATS成瘾的现阶段研究进展进行综述,旨在阐明miRNAs在ATS成瘾中的表达特征及调控功能,为进一步研究提供参考。
1 miRNAs在ATS成瘾中的表达特征
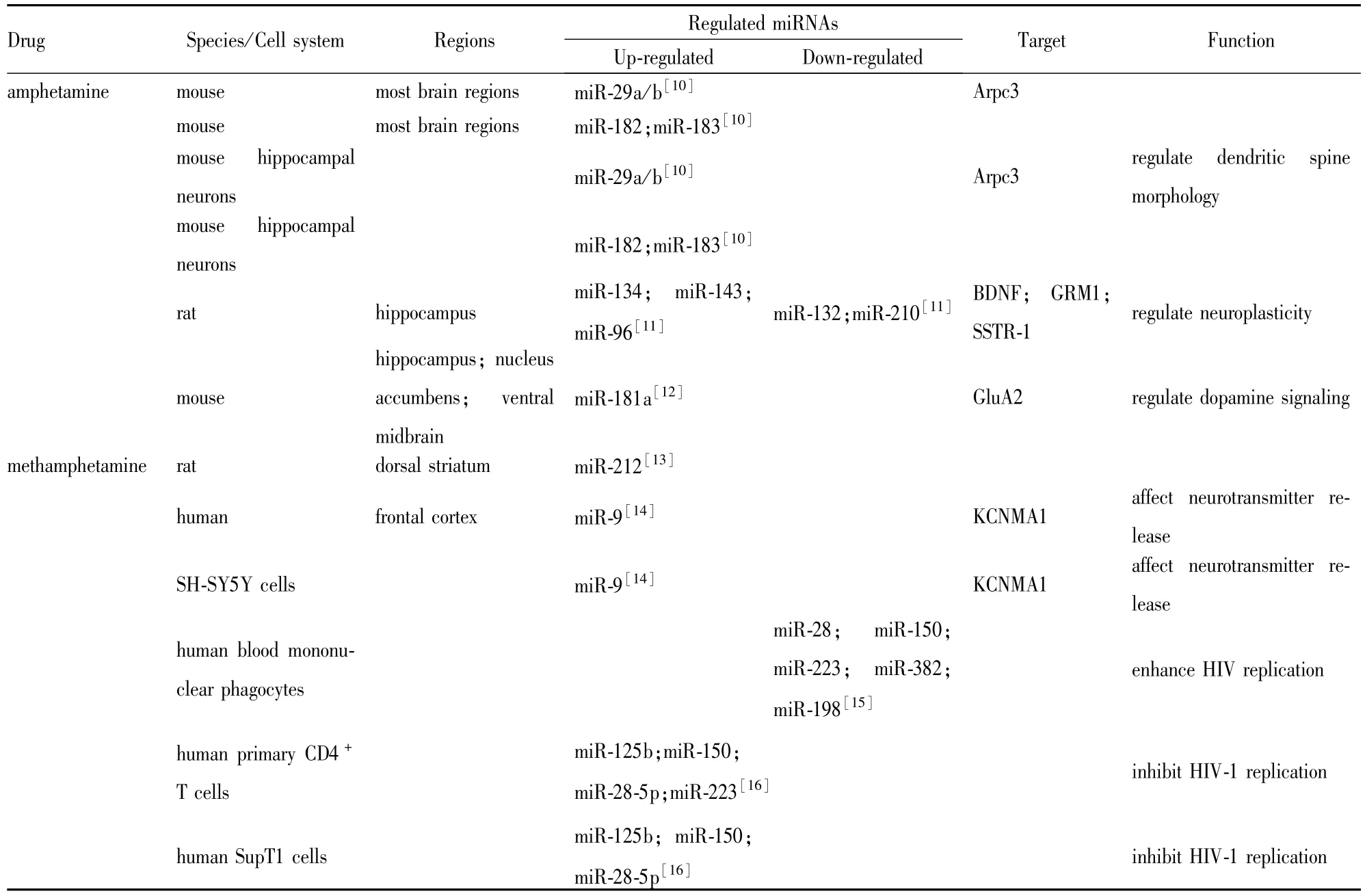
在不同成瘾性物质作用下,成瘾相关脑区中多种miR-NAs的表达水平发生特异性改变。2008年Pietrzykowski等[6]首次报道酒精导致大鼠视上核和纹状体神经元miR-9明显上调;2009年,Chandrasekar等[7]发现可卡因慢性处理能上调大鼠伏隔核和海马区miR-181a,而下调中脑腹侧被盖区和海马区miR-124和let-7d;Huang等[8]用尼古丁处理PC12细胞发现miR-140明显上调;Wu等[9]发现吗啡通过上调miR-23b导致μ-阿片受体蛋白水平降低;2011年,Lippi等[10]开始报道苯丙胺慢性处理可改变小鼠多个脑区的miR-NAs表达水平。目前发现的苯丙胺类兴奋剂成瘾中变化的miRNAs及其功能见Tab 1。
Lippi等[10]发现苯丙胺慢性处理可使C57BL6小鼠海马、前额叶皮质、皮质下前脑边缘和中脑腹侧区域的共32个miRNAs明显上调。苯丙胺能同时上调小鼠多个脑区的miR-29a/b、miR-182/183、miR-680和miR-190b表达水平,而针对性地上调中脑腹侧和皮质下前脑边缘区的miR-200/429家族水平。进一步用苯丙胺处理原代小鼠海马神经元
发现,体内实验中明显上调的32个miRNAs只有一小部分也出现上调,其中miR-29a/b上调3.5~4.2倍,miR-182/183家族上调13.6~17.6倍,而miR-680和miR-190b没有发生明显改变。荣晗等[11]通过大鼠腹腔注射苯丙胺2周建立大鼠狂躁模型,发现大鼠海马中miR-134、miR-143、miR-96出现明显上调,而miR-133、miR-210明显下调,表明苯丙胺能差异调控不同miRNAs的表达。Saba等[12]研究发现,苯丙胺和可卡因慢性处理能分别诱导小鼠特定脑区的miR-181a出现上调。苯丙胺能使小鼠海马、伏隔核和中脑腹侧区的miR-181a表达上调,其中以海马中miR-181a上调程度最多,而前额叶皮质中miR-181a未发生明显改变。相反,可卡因能使小鼠前额叶皮质中miR-181a出现明显上调,而不改变海马、伏隔核和中脑腹侧脑区中miR-181a的表达,表明不同成瘾药物诱导的miRNAs差异表达具有组织特异性。
Tab 1 miRNAs regulations in ATS addiction
金雪峰[13]在甲基苯丙胺大鼠自身给药模型中发现,miR-212在强迫性用药组动物背侧纹状体的表达上调,与可卡因自身给药模型的研究结果一致,提示miR-212在药物成瘾的强迫性用药行为中扮演重要的角色。同时,该研究发现miR-212仅在长时程给药动物中表达上调,在短时程给药动物中不发生表达变化,表明甲基苯丙胺诱导miRNAs的差异表达与给药时程密切相关。Tatro等[14]研究发现,HIV阳性甲基苯丙胺滥用患者的前额叶皮质中miR-9、miR-124a和let-7d明显增加。进一步利用多巴胺能神经元体外模型研究甲基苯丙胺和HIV对miRNAs表达的影响,发现用甲基苯丙胺和含感染HIV巨噬细胞的培养基对SH-SY5Y细胞进行处理亦使miR-9、miR-124a和let-7d明显上调。Wang等[15]分别用100和250 μmol·L-1甲基苯丙胺处理巨噬细胞,发现5个抗HIV miRNAs:miR-28、miR-150、miR-223、miR-382和miR-198均明显下调。另一项研究[16]观察甲基苯丙胺对原代CD4+T淋巴细胞和SupT1细胞株中抗HIV miRNAs水平的影响,发现当甲基苯丙胺浓度大于100 μmol·L-1能剂量依赖地上调两种细胞中miR-125b、miR-150和miR-28-5p表达。甲基苯丙胺还能上调原代CD4+T淋巴细胞中miR-223水平,而miR-223在SupT1细胞中却不表达。尽管两项研究得到相反的结果,但均表明甲基苯丙胺能诱导抗HIV miR-NAs的差异表达。
2 miRNAs在ATS成瘾中的调控机制
中枢神经系统中含有大量的miRNAs,它们对神经系统具有重要的调控作用。MiRNAs是突触可塑性、突触功能及形态修饰的调节因子,并且与高级认知功能,如学习和记忆有关[4-5]。据报道,在药物成瘾形成过程中发挥重要作用的多巴胺受体1表达与miR-504密切相关,miR-504参与多巴胺神经元回路的调控[17],而且miRNAs还直接调节谷氨酸受体的活性[18]。越来越多的证据显示,成瘾性物质能诱导miRNAs表达水平的变化,而成瘾相关脑区miRNAs表达的改变直接参与了对成瘾行为的调节[19-20]。
MiRNAs可通过调节突触可塑性和神经递质的释放参与苯丙胺的成瘾过程。Saba等[12]给小鼠腹腔连续注射苯丙胺5 d,发现其海马、伏隔核、中脑腹侧等多个脑区miR-181a表达明显上调,以海马区最为明显,并且在海马神经元中激活多巴胺信号可诱导miR-181a表达;过表达和抑制表达实验证实,miR-181a是通过负调控谷氨酸受体2亚单位(iono-tropic glutamate receptor AMPA alpha 2,GluA2)的表达进而调节突触功能。该研究认为miR-181a是哺乳动物α-氨基羟甲基异恶唑丙酸型谷氨酸受体(alpha-amino-3-hydroxy-5-methyl-4-isoxazole propionate,AMPA)的一个关键调节因子,对ATS引起的突触可塑性改变有潜在影响。苯丙胺慢性处理导致小鼠多个脑区miR-29a/b出现明显上调,进一步研究发现,miR-29a/b能导致肌动蛋白细胞骨架重塑,影响突触形成,是突触形态的一个调节因子[10]。MiR-29a/b通过靶向肌动蛋白相关蛋白2/3复合物亚单位3(actin related protein 2/3 complex subunit 3,Arpc3)调节树突棘形态的变化,可能增加奖赏通路的敏感性,引起神经元可塑性的改变,进而导致苯丙胺依赖的形成。研究发现[11],给予苯丙胺处理的大鼠海马中miR-134、miR-143、miR-96、miR-132和miR-210发生明显改变,而这5个miRNAs均与神经元可塑性有关。MiR-134可调节突触传递,降低人单丝氨酸蛋白激酶1基因的表达,减少树突棘的大小而不影响树突棘的数目[21]。MiR-143与神经干细胞的分化、发育和神经元可塑性密切有关[22]。MiR-96抑制5-羟色胺1B及5-羟色胺3E受体的转录,且在习得性无助大鼠模型中表达下调,提示其可能在调节神经元可塑性方面起到一定作用[23]。MiR-132可能通过调节环单磷酸腺苷反应元件结合蛋白(cAMP response element binding protein,CREB)和脑源性神经营养因子(brain-derived neuro-trophic factor,BDNF)介导的信号通路促进新生神经元树突成熟的过程[24]。MiR-210在神经细胞缺氧状态下表达增高,可能对神经细胞增殖、凋亡等起到重要作用[25]。进一步对这5 个miRNAs进行靶基因预测和验证发现,其可能分别通过靶向代谢型谷氨酸受体1(glutamate receptor metabotropic 1,GRM1)、BDNF以及生长抑素受体1(somatostatin receptor 1,SSTR-1)调节神经元可塑性和神经递质的释放。
MiRNA能影响甲基苯丙胺成瘾过程中神经递质的释放。一项对HIV阳性甲基苯丙胺滥用患者的研究表明,甲基苯丙胺和HIV均能诱导miR-9的表达上调[14]。MiR-9通过和大电导钙和电压激活的钾通道亚家族M成员α1(potassium large conductance calcium-activated channel subfamily M alpha member 1,KCNMA1)mRNA的3′非编码区结合,剪切KC-NMA1基因的转录产物,导致大电导钙和电压激活的钾通道剪接变异体的差异表达,进而影响多巴胺能神经元中神经递质的释放。给予甲基苯丙胺处理能使巨噬细胞中抗HIV miRNAs:miR-28、miR-150、miR-223、miR-382和miR-198明显下调[15]。进一步研究,发现甲基苯丙胺可能通过调控干扰素信号通路增加巨噬细胞感染HIV。然而,另一项研究表明,甲基苯丙胺能明显上调CD4+T淋巴细胞中抗HIV-1 miRNAs如miR-125b、miR-150和miR-28-5p的表达,转染miRNAs拮抗剂对miR-125b和miR-28-5p进行沉默发现抗HIV-1 miRNAs能负调控HIV-1的复制[16]。这两项研究中甲基苯丙胺对于抗HIV miRNAs的表达出现相反的结果,推测可能与作用在不同细胞密切有关,同时也表明抗HIV miR-NAs可能参与了甲基苯丙胺成瘾的调控机制,甲基苯丙胺和HIV之间可能存在更为复杂的相互作用,尚待进一步研究。
3 小结与展望
ATS的成瘾机制复杂,涉及脑内奖赏系统多个脑区的相互作用及互相调控。目前对其成瘾机制的研究主要集中在脑内的多巴胺、5-羟色胺、谷氨酸等中枢神经递质以及调控神经元基因表达的转录因子如CREB和ΔfosB蛋白等。随着对miRNA研究的不断深入,发现其在药物成瘾的形成过程中发挥着重要的调控作用。ATS使奖赏脑区相关miRNA的表达水平发生特异性改变,进而通过负调控一些关键基因的表达,调节神经元可塑性和神经递质的释放,导致成瘾记忆的形成,并使成瘾记忆与动机系统出现异常神经联系,形成异常的觅药动机和强迫性觅药行为,从而产生药物成瘾。阐明ATS成瘾中miRNAs的表达特征及调控功能,为研究新型毒品的成瘾机制提供新的可供参考的生物信息,同时也为发现新的药物作用靶点,建立新的防治策略提供科学依据。
(致谢:本文在南方医科大学中医药学院中药药理实验室完成,感谢李婵、林莹波和朱道琦对文章相关工作的支持,感谢香港Ka Lok Cheng先生对英文摘要的修改!)
参考文献:
[1] 杨黎华,白 洁.单胺类神经递质在药物成瘾中的作用机制[J].中国药理学通报,2015,31(2):149-52.
[1] Yang L H,Bai J.Roles of monoamine neurotransmitters in the mechanism of drug addiction[J].Chin Pharmacol Bull,2015,31 (2):149-52.
[2] Robison A J,Nestler E J.Transcriptional and epigenetic mecha-nisms of addiction[J].Nat Rev Neurosci,2011,12(11):623-37.
[3] Brensilver M,Heinzerling K G,Shoptaw S.Pharmacotherapy of amphetamine-type stimulant dependence:An update[J].Drug Al-cohol Rev,2013,32(5):449-60.
[4] Fiore R,Schratt G.MicroRNAs in synapse development:tiny mol-ecules to remember[J].Expert Opin Biol Ther,2007,7(12):1823-31.
[5] Im H I,Kenny P J.MicroRNAs in neuronal function and dysfunc-tion[J].Trends Neurosci,2012,35(5):325-34.
[6] Pietrzykowski A Z,Friesen R M,Martin G E,et al.Posttranscrip-tional regulation of BK channel splice variant stability by miR-9 un-derlies neuroadaptation to alcohol[J].Neuron,2008,59(2):274 -87.
[7] Chandrasekar V,Dreyer J L.MicroRNAs miR-124,let-7d and miR-181a regulate cocaine-induced plasticity[J].Mol Cell Neuros-ci,2009,42(4):350-62.
[8] Huang W,Li M D.Nicotine modulates expression of miR-140,
which targets the 3′-untranslated region of dynamin 1 gene (Dnm1)[J].Int J Neuropsychoph,2009,12(4):537-46.
[9] Wu Q,Zhang L,Law P Y,et al.Long-term morphine treatment decreases the association of μ-opioid receptor(MOR1)mRNA with polysomes through miRNA-23b[J].Mol Pharmacol,2009,75(4):744-50.
[10]Lippi G,Steinert J R,Marczylo E L,et al.Targeting of the Arpc3 actin nucleation factor by miR-29a/b regulates dendritic spine mor-phology[J].J Cell Biol,2011,194(6):889-904.
[11]荣 晗,刘铁榜,杨海晨,等.苯丙胺模型大鼠海马差异表达微小核糖核酸的研究[J].中华精神科杂志,2013,46(5):301 -5.
[11]Rong H,Liu T B,Yang H C,et al.Microaaray-based analysis of micro-ribonucleic acid expression in an animal model of ampheta-mine[J].Chin J Psychiatry,2013,46(5):301-5.
[12]Saba R,Strchel P H,Aksoy-Aksel A,et al.Dopamine-regulated microRNA miR-181a controls GluA2 surface expression in hipp-ocampal neurons[J].Mol Cell Biol,2012,32(3):619-32.
[13]金雪峰.甲基苯丙胺成瘾大鼠中枢及血浆中差异表达microR-NA的发现[D].长沙:中南大学,2013.
[13]Jin X F.Discovery of microRNA with differential expressions in central nervous system and plasma of methamphetamine-addicted rats[D].Changsha:Central South University,2013.
[14]Tatro E T,Hefler S,Shumaker-Armstrong S,et al.Modulation of BK channel by microRNA-9 in neurons after exposure to HIV and methamphetamine[J].J Neuroimmune Pharmacol,2013,8(5):1210-23.
[15]Wang X,Wang Y Z,Ye L,et al.Modulation of intracellular re-striction factors contributes to methamphetamine-mediated enhance-ment of acquired immune deficiency syndrome virus infection of macrophages[J].Curr HIV Res,2012,10(5):407-14.
[16]Mantri C K,Mantri J V,Pandhare J,Dash C.Methamphetamine inhibits HIV-1 replication in CD4+T cells by modulating anti-HIV-1 miRNA expression[J].Am J Pathol,2014,184(1):92-100.
[17]Huang W,Li M D.Differential allelic expression of dopamine D1 receptor gene(DRD1)is modulated by microRNA miR-504[J].Biol Psychiat,2009,65(8):702-5.
[18]Karr J,Vagin V,Chen K,et al.Regulation of glutamate receptor subunit availability by microRNAs[J].J Cell Biol,2009,185 (4):685-97.
[19]Chandrasekar V,Dreyer J L.Regulation of miR-124,let-7d,and miR-181a in the accumbens affects the expression,extinction,and reinstatement of cocaine-induced conditioned place preference[J].Neuropsychopharmacology,2011,36(6):1149-64.
[20]Hollander J A,Im H I,Amelio A L,et al.Striatal microRNA con-trols cocaine intake through CREB signalling[J].Nature,2010,466(7303):197-202.
[21]Gao J,Wang W Y,Mao Y W,et al.A novel pathway regulates memory and plasticity via SIRT1 and miR-134[J].Nature,2010,466(7310):1105-9.
[22]Tam S T,Bastian I,Zhou X F,et al.MicroRNA-143 expression in dorsal root ganglion neurons[J].Cell Tissue Res,2011,346 (2):163-73.
[23]Millan M J.MicroRNA in the regulation and expression of seroton-ergic transmission in the brain and other tissues[J].Curr Opin Pharmacol,2011,11(1):11-22.
[24]Magill S T,Cambronne X A,Luikart B W,et al.MicroRNA-132 regulates dendritic growth and arborization of newborn neurons in the adult hippocampus[J].Proc Natl Acad Sci USA,2010,107 (47):20382-7.
[25]Xiong L,Wang F,Huang X,et al.DNA demethylation regulates the expression of miR-210 in neural progenitor cells subjected to hypoxia[J].FEBS J,2012,279(23):4318-26.
Research progress on the role of miRNAs in amphetamine-type stimulants addiction
JIANG Ming-jin,LI Chan,LIN Ying-bo,ZHU Dao-qi,MO Zhi-xian
(Dept of Pharmacology,School of Traditional Chinese Medicine,Southern Medical University,Guangzhou 510515,China)
Abstract:Amphetamine-type stimulants(ATS),a group of new-type synthetic drugs mainly in psychological dependence,are abused more and more severely in recent years.MicroRNAs (MiRNAs)are an important class of endogenous non-coding small RNAs that mediate posttranscriptional negatively regulation of gene expression by targeting specific mRNA sequences to in-hibit the translation of mRNAs or degrade the expression of mR-NAs.ATS can induce the changes in the expression of miRNAs in addiction-related brain regions which directly involve in the regulation of ATS-induced addictive behaviors.Therefore,to study the regulatory role of miRNAs in ATS-induced addiction has important implications for dependent mechanisms of new-type drugs and the discovery of the new targets of drug actions.
Key words:miRNAs;amphetamine-type stimulants;ampheta-mine;methamphetamine;addiction;expression;regulation;mechanism
作者简介:江明金(1988-),男,博士生,研究方向:中药有效成分抗药物成瘾的临床和实验研究,Tel:020-61648540,E-mail:comity2006@126.com;莫志贤(1958-),女,硕士,教授,博士生导师,研究方向:中药有效成分抗药物成瘾的临床和实验研究,通讯作者,Tel:020-61648261,E-mail:cherrymo@fimmu.com
基金项目:国家自然科学基金资助项目(No 81229003,81173581)
收稿日期:2015-05-27,修回日期:2015-08-15
文献标志码:A
文章编号:1001-1978(2015)10-1352-04
doi:10.3969/j.issn.1001-1978.2015.10.006