三种柔鱼类SSR标记的通用性检测
2015-02-22隋宥珍刘连为陈新军
隋宥珍,刘连为,2,陈新军
(1.农业部重点渔场渔业资源科学观测实验站,浙江省海洋渔业资源可持续利用技术研究重点实验室,浙江省海洋水产研究所,浙江舟山 316021;2.浙江海洋学院,浙江舟山 316022;3.大洋渔业资源可持续开发省部共建教育部重点实验室,上海海洋大学海洋科学学院,上海 201306)
三种柔鱼类SSR标记的通用性检测
隋宥珍1,刘连为1,2,陈新军3
(1.农业部重点渔场渔业资源科学观测实验站,浙江省海洋渔业资源可持续利用技术研究重点实验室,浙江省海洋水产研究所,浙江舟山 316021;2.浙江海洋学院,浙江舟山 316022;3.大洋渔业资源可持续开发省部共建教育部重点实验室,上海海洋大学海洋科学学院,上海 201306)
分析了柔鱼、阿根廷滑柔鱼、茎柔鱼3种柔鱼类多态性微卫星标记在种间的通用性。结果表明,柔鱼与茎柔鱼多态性SSR标记在阿根廷滑柔鱼中不适用,同样,阿根廷滑柔鱼多态性SSR标记在二者中也不适用。柔鱼3个多态性SSR标记(位点Bo101、Bo106、Bo1018)在茎柔鱼DNA样本中扩增时显示出单态性,而茎柔鱼10个多态性SSR标记在柔鱼中具有较好的适用性。10个SSR位点(DG01、DG04、DG10、DG13、DG19、DG28、DG37、DG38、DG40、DGI8)等位基因数为4~44个,观测杂合度介于0.350~0.950之间,期望杂合度介于0.615~0.972之间。多态信息含量介于0.525~0.959之间,均为高度多态性位点(PIC>0.5)。经Bonferroni校正后,位点DG01、DG19极显著偏离Hardy-Weinberg平衡(P<0.01)。柔鱼群体的平均等位基因数、平均观测杂合度、平均期望杂合度、多态信息含量分别为13.40、0.631、0.814、0.723,显示出较高的遗传多样性水平。由此可见,茎柔鱼类多态性SSR标记在柔鱼中具有较高的通用性。
柔鱼;阿根廷滑柔鱼;茎柔鱼;微卫星标记;通用性
头足类被认为是世界海洋渔业资源中3种未充分开发利用的种类之一,其中规模性开发的种类中以柔鱼类所占比重最大,主要包括太平洋褶柔鱼Todarodes pacificus、柔鱼Ommastrephes batramii、双柔鱼Nototodarus sloani、阿根廷滑柔鱼 Illex argentinus、茎柔鱼 Dosidicus gigas、鸢乌贼 Symplectoteuthis oualaniensis等[1-2]。目前,柔鱼、阿根廷滑柔鱼、茎柔鱼已成为我国远洋鱿鱼钓的主要捕捞对象①上海海洋大学鱿钓技术组。2002-2013年我国鱿钓船柔鱼、阿根廷滑柔鱼、茎柔鱼生产总结。。柔鱼、阿根廷滑柔鱼、茎柔鱼分别隶属于柔鱼科中的柔鱼属Ommastrephes、滑柔鱼属Illex与茎柔鱼属Dosidicus,资源丰富且分布广泛,有关其种群遗传学研究已有大量报道[3-8]。其中,微卫星simple sequence repeat,SSR标记由于具有高度多态性、共显性、实验操作相对简单等优点而得以广泛应用[6-8]。目前,3种柔鱼类SSR标记均已开发出,但柔鱼与阿根廷滑柔鱼多态性SSR标记数目较少,仍需开发出更丰富的多态性SSR标记,以便更好地揭示其在全球分布范围内的遗传背景。由于SSR侧翼序列在近缘物种间较为保守,因此,采用近缘物种转移法是获得SSR标记的简便、快速的方法[9]。为此,本文分析了3种柔鱼类多态性SSR标记在种间的通用性。
1 材料与方法
1.1 实验材料与基因组DNA的提取
柔鱼、阿根廷滑柔鱼与茎柔鱼捕获后立刻剪取尾鳍肌肉组织保存于95%乙醇中,存放于船舱冷库中并运回至实验室。分别选取40个个体用于SSR标记的通用性检测,柔鱼样本采自西北太平洋海域(40°41′-42°34′N、149°29′-155°10′E),阿根廷滑柔鱼样本采自阿根廷专属经济区以外海域(45°17′-47°20′S、60° 16′-60°49′W),茎柔鱼样本采自赤道海域(3°N-5°S、114°-120°W)。采用Genomic DNA Isolation Kit(福际生物技术有限公司)提取基因组DNA,用1.2%琼脂糖凝胶电泳检测DNA质量,紫外分光光度计检测DNA浓度。
1.2 微卫星PCR扩增
柔鱼8对SSR引物引自刘连为等[6](位点:Bo101~Bo108),茎柔鱼28对SSR引物引自Liu等[10](位点:DG01、DG04~DG11、DG13~DG16、DG18~DG21、DG26、DG28~DG30、DG34、DG36~DG40、DGI8),阿根廷滑柔鱼7对SSR引物引自Adock等[11](位点:Ia112、Ia121a、Ia121b、Ia203、Ia207、Ia408、Ia422)。SSR引物及荧光标记SSR引物由上海杰李生物技术有限公司合成。首先利用普通SSR引物进行PCR扩增,PCR产物经过琼脂糖凝胶电泳初步筛选出能够扩增出清晰条带的SSR标记。然后用带有荧光标记的SSR引物进行PCR扩增,通过基因分型技术筛选多态性SSR标记。PCR反应总体积均为25 μL,其中10×PCR Buffer 2.5μL、Taq DNA polymerase(5 U/μL)0.2 μL、dNTP(各2.5 mmol/L)2 μL、上下游引物(10 μmol/L)各0.6 μL、DNA模板20 ng、ddH2O补足体积。反应程序为:94℃预变性2 min;94℃变性30 s,退火30 s,72℃延伸45 s,35个循环;72℃最后延伸2 min。
1.3 PCR产物的纯化及其分子量数据的读取
带有荧光标记的PCR产物经过1.2%琼脂糖凝胶电泳分离,用Biospin Gel Extraction Kit纯化后,送至上海迈浦生物科技有限公司。PCR产物稀释后与分子量内标(ROX-500)混合,通过ABI3730XL全自动DNA测序仪进行毛细血管电泳,利用Genemapper Version 3.5软件读取微卫星扩增产物的分子量数据。
1.4 数据统计与分析
根据分子量数据确定个体各位点基因型,利用Popgen 3.2[12]进行群体遗传学分析,计算等位基因数(Na),有效等位基因数(Ne),观测杂合度(Ho),期望杂合度(He)与Shannon多样性指数(Shannon's information index,I)。多态信息含量(PIC)由Cervus 3.0[13]软件计算,并采用马尔科夫链(Markov Chain)方法进行Hardy-Weinberg平衡检验。
2 结果
琼脂糖凝胶电泳检测结果显示,阿根廷滑柔鱼7对SSR引物在柔鱼、茎柔鱼DNA样本中均未扩增出任何条带;柔鱼8对SSR引物在阿根廷滑柔鱼DNA样本中未扩增出任何条带,而在茎柔鱼DNA样本中有3对引物扩增出清晰条带;茎柔鱼28对SSR引物在阿根廷滑柔鱼DNA样本中未扩增出任何条带,而在柔鱼DNA样本中有15对引物扩增出清晰条带。用带有荧光标记的SSR引物进行PCR扩增,通过等位基因分型技术进一步筛选多态性SSR标记。其中,在茎柔鱼DNA样本中扩增出清晰条带的柔鱼 3个位点(Bo101、Bo106、Bo108)为单态性位点;在柔鱼DNA样本中扩增出清晰条带的茎柔鱼 5个位点(DG08、DG11、DG18、DG29、 DG36)为单态性位点,10个位点为多态性位点(表1)。图1为位点DG40 PCR扩增的琼脂糖电泳检测结果。
图1 茎柔鱼位点DG40对柔鱼PCR扩增结果Fig.1 Results of PCR amplification in Ommastrephes bartramii by locus DG40 from Dosidicus gigas
茎柔鱼10对SSR引物的核心序列、扩增产物大小、退火温度以及SSR位点在柔鱼群体中的扩增结果见表1。各位点等位基因数为4~44个,观测杂合度介于0.350~0.950之间,期望杂合度介于0.615~0.972之间。多态信息含量介于0.525~0.959之间,均为高度多态性位点(PIC>0.5)。经Bonferroni校正后,位点DG01、DG19极显著偏离Hardy-Weinberg平衡(P<0.01)。柔鱼群体的平均等位基因数、平均观测杂合度、平均期望杂合度、多态信息含量分别为13.40、0.631、0.814、0.723,显示出较高的遗传多样性水平。
3 讨论
获得SSR标记有多种方法,常见方法为磁珠富集法和基于第二代测序技术的基因组测序或转录组测序,如上文提到的柔鱼与茎柔鱼SSR标记的开发[6,10]。而获得SSR标记的简便、快速的方法为检索Gen-Bank、EMBL、DDBJ等DNA序列数据库,或者利用亲缘性较近的物种的SSR标记。作者利用基因组测序方法获得茎柔鱼39个多态性SSR标记,并应用于东南太平洋茎柔鱼群体遗传学研究,而通过磁珠富集法从柔鱼部分微卫星富集文库中筛选出8个多态性SSR标记以及已报道的阿根廷滑柔鱼多态性SSR标记仅8个,均不能满足其群体遗传学研究[6,10,11]。本文采用近缘物种转移法分析了3种柔鱼类多态性SSR标记在种间的通用性。结果表明,柔鱼与茎柔鱼SSR标记在阿根廷滑柔鱼中不适用,同样,阿根廷滑柔鱼SSR标记在二者中也不适用。茎柔鱼SSR标记在柔鱼中具有较好的适用性,而柔鱼3个SSR标记在茎柔鱼DNA样本中扩增时显示出单态性,这可能与选择的柔鱼SSR标记数量太少有关。柔鱼与茎柔鱼隶属于柔
鱼亚科(Ommastrephinae),而阿根廷滑柔鱼隶属于滑柔鱼亚科(Illicinae),它们在分类地位上的差异可能造成种间SSR标记适用性的差异。在今后的研究中,可将茎柔鱼多态性SSR标记用于柔鱼的群体遗传学研究。
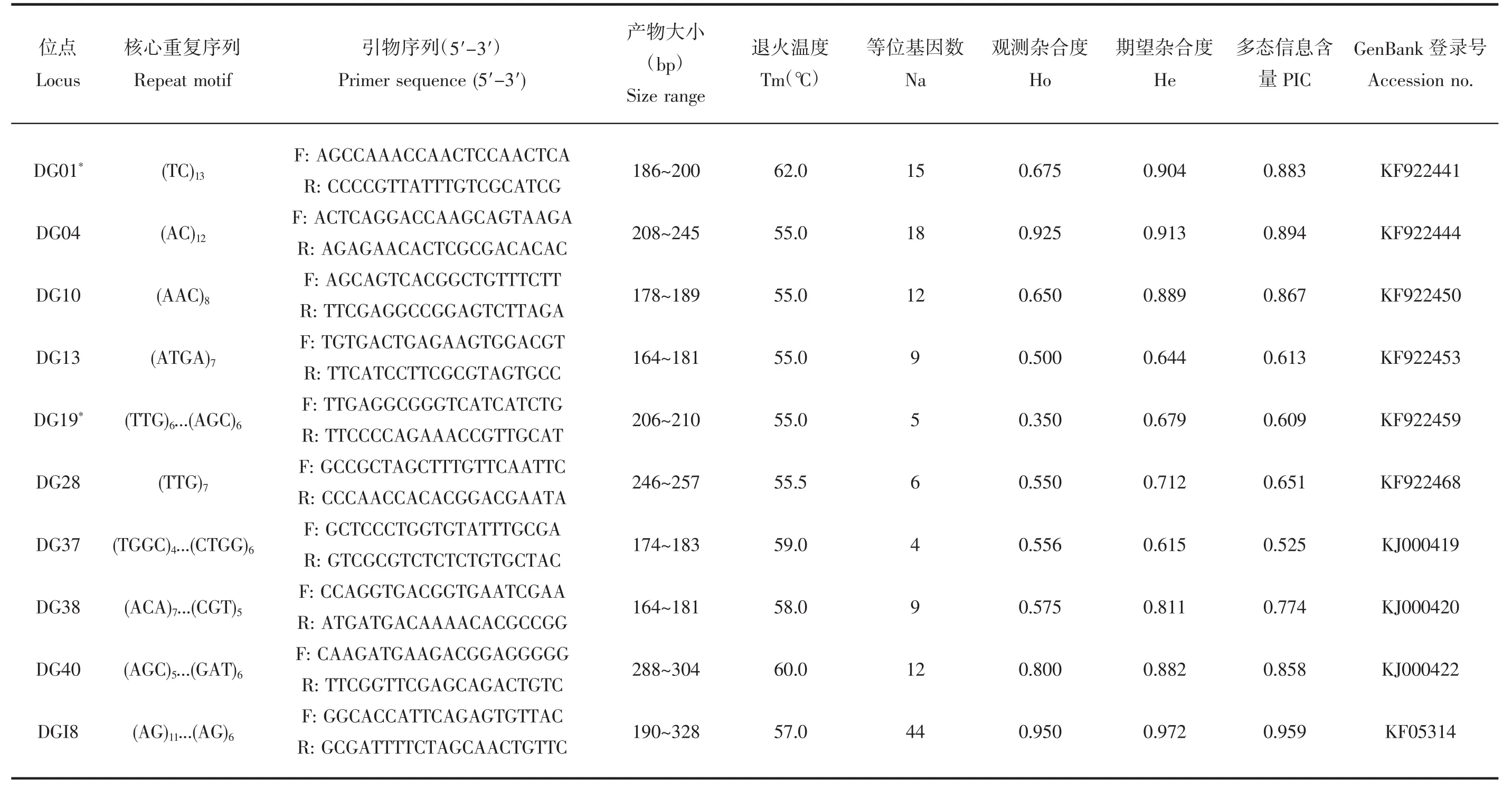
表1 茎柔鱼SSR 标记特征及其对柔鱼中的通用性Tab.1 Characterization of microsatellite markers derived from D. gigas and their transferability in O. bartramii
位点DG01、DG19极显著偏离Hardy-Weinberg平衡(P<0.01),且它们均存在纯合体个体过剩现象。因此,这2个位点很可能存在无效等位基因(null allele)。无效等位基因产生的原因主要为SSR侧翼序列变异,从而导致该位点无法正常扩增,电泳检测时无可见的扩增条带或者杂合子只表现出一条带[14]。建议在以后的实验中可通过重新设计特异性SSR引物从根本上消除无效等位基因的影响或者估算无效等位基因频率进行群体遗传学研究。
[1]陈新军,陆化杰,刘必林,等.大洋性柔鱼类资源开发现状及可持续利用的科学问题 [J].上海海洋大学学报,2012,21(5): 831-840.
[2]陈新军,刘必林,王尧耕.世界头足类[M].北京:海洋出版社,2009.
[3]刘连为,许强华,陈新军.基于线粒体COⅠ和Cytb基因序列的北太平洋柔鱼种群遗传结构研究[J].水产学报,2012,36 (11):1 675-1 684.
[4]刘连为,陈新军,许强华,等.基于线粒体DNA标记的阿根廷滑柔鱼2个产卵群体遗传变异分析[J].海洋渔业,2014,36 (1):16-23.
[5]闫 杰,许强华,陈新军,等.东太平洋公海茎柔鱼种群遗传结构初步研究[J].水产学报,2011,35(11):1 617-1 623.
[6]刘连为,陈新军,许强华,等.北太平洋柔鱼微卫星引物的筛选及遗传多样性分析[J].生态学报,2014,34(23):6 847-6 854.
[7]刘连为,陈新军,许强华,等.阿根廷滑柔鱼2个产卵群体遗传变异的微卫星分析[J].海洋渔业,2013,35(2):125-130.
[8]刘连为,陈新军,许强华,等.茎柔鱼赤道海域群体与秘鲁外海群体遗传变异的微卫星分析[J].中国海洋大学学报:自然科学版,2014,45(7):53-57.
[9]SUN Dian-qiao,LI Hai-yan,XU Tian-jun,et al.Development of microsatellite markers for the small yellow croaker Larimichthys polyactis(Sciaenidae)by cross-species amplification[J].Genetics and Molecular Research,2012,11:1 469-1 474.
[10]LIU Lian-wei,CHEN Xin-jun,XU Qiang-hua,et al.Development of 39 polymorphic microsatellite markers for the jumbo squid Dosidicus gigas using Illumina paired-end sequencing[J].Conservation Genetics Resources,2014,6:673-676.
[11]ADOCK G J,CARVALHO G R,RODHOUSE P G,et al.Highly polymorphic microsatellite loci of the heavily fished squid genus Illex(Ommastrephidea)[J].Molecular Ecology,1999,8(1):157-168.
[12]NEI M.Estimation of average heterozygosity and genetic distance from a small number of individuals[J].Genetics,1978,89(3): 583-590.
[13]KALINOWSKI S T,TAPER M L,MARSHALL T C.Revising how the computer program cervus accommodates genotyping error increases success in paternity assignment[J].Molecular Ecology,2007,16(5):1 099-1 106.
[14]文亚峰,UCHIYAMA K,韩文军,等.微卫星标记中的无效等位基因[J].生物多样性,2013,21(1):117-126.
Cross-species Amplifiation of Microsatellite Markers in Three Ommastrephidae Species
SUI You-zhen1,LIU Lian-wei1,2,CHEN Xin-jun3
(1.Scientific Observing and Experimental Station of Fishery Resources for Key Fishing Grounds,MOA,Key Laboratory of Sustainable Utilization of Technology Research for Fisheries Resources of Zhejiang Province, Marine Fisheries Research Institute of Zhejiang Province,Zhoushan 316021;2.Zhejiang Ocean University, Zhoushan 316022;3.The Key Laboratory of Sustainable Exploitation of Oceanic Fisheries Resources, Ministry of Education,College of Marine Sciences,Shanghai Ocean University,Shanghai 201306,China)
The transferability of polymorphic short sequence repeats(SSRs)from Ommastrephes bartramii, Illex argentinus and Dosidicus gigas was assessed in the three closely-related species.A population of 40 individuals of each Ommastrephidae species were tested with 7 loci from I.argentinus,8 loci from O.bartramiiand 28 loci from D.gigas.The results showed that all the loci from I.argentinus failed to be amplified in O. bartramii and D.gigas,while all the loci from O.bartramii and D.gigas failed to be amplified in I.argentinus. Three loci(Bo101,Bo106 and Bo1018)from O.bartramii were monomorphic in D.gigas,and 10 loci(DG01, DG04,DG10,DG13,DG19,DG28,DG37,DG38,DG40 and DGI8)from D.gigas were polymorphic in O.bartramii.The number of alleles per polymorphic locus ranged from 4 to 44(mean=13.40)in these 10 loci,and the observed and expected heterozygosity ranged from 0.350 to 0.950(mean=0.631)and from 0.615 to 0.972 (mean=0.814),respectively.Two loci(DG01,DG19)deviated significantly from the HWE test after Bonferroni correction(P<0.05),and the polymorphism information content(PIC)of 10 loci ranged from 0.525 to 0.959 (mean=0.723),which indicated all the loci were highly polymorphic.From this we can see that these 10 loci from D.gigas are expected to be useful for further studies of genetic diversity of O.bartramii.
Ommastrephes bartramii;Illex argentinus;Dosidicus gigas;microsatellite markers;crossspecies amplifiation
S917.4
A
1008-830X(2015)06-0532-05
2015-09-10
国家自然科学基金(41276156);国家高技术研究发展计划(863计划)(2012AA092303);上海市科技创新行动计划(12231203900)
隋宥珍(1983-),女,辽宁大连人,研究方向:分子生态学.E-mail:shuichansui@126.com
陈新军,男,教授.E-mail:xjchen@shou.edu.cn
