基于突触连接视通路方位敏感的图像分级边缘检测方法
2015-02-20范影乐张梦楠
王 典 范影乐 张梦楠 武 薇
(杭州电子科技大学模式识别与图像处理实验室,杭州 310018)
基于突触连接视通路方位敏感的图像分级边缘检测方法
王 典 范影乐*张梦楠 武 薇
(杭州电子科技大学模式识别与图像处理实验室,杭州 310018)
视觉通路上的多级方位敏感特性对于视觉轮廓感知起着关键作用,将为更高层次的视皮层图像理解提供重要的特征信息。从视觉方位敏感机制出发,提出一种图像边缘检测的新方法。利用神经节细胞以及外膝体神经元感受野向心分布的生理结构特性,构建具有突触连接和多方向敏感特性的视皮层下功能层,融合多方向上的神经元脉冲发放信息,将视觉激励映射为边缘敏感图像;构建具有去最优方位感受野特性的初级视皮层的功能层,对前级结构生成的脉冲序列按时间信息进行神经编码,经过感受野内侧向抑制和阈值处理,获得边缘检测结果。对层次模糊而细节丰富的菌落图像进行处理,并以边缘置信度和重构相似度以及两者的加权和作为边缘检测评价指标。结果表明,该方法在完整检测图像边缘的同时,并不引入纹理噪声,有着明显的优势,其对12幅图像的加权和指标均值为0.746 8,显著高于其他对比方法。所提出的方法可以模拟视通路中初级视皮层及视皮层下的方向敏感特性,提供一种基于视觉机制的图像处理和理解新思路。
边缘检测;视通路;突触连接;方位敏感;侧向抑制
引言
轮廓特征提取将为图像理解或者运动目标行为分析提供重要的降维信息,其中在准确定位边缘的前提下不发生漏检以及避免出现虚假边缘是对轮廓特征描述最基本的两项要求。传统边缘检测方法,通常试图利用数学模型来刻画局部邻域像素之间在特定方向上的灰度变化[1]。例如Canny边缘检测方法,具有运算效率高和实时性较好的特点,但由于其从图像梯度出发设定阈值,面对具有多强度边缘或者丰富细节的图像时,很难同时满足上述两项要求。
考虑到人类视觉在图像信息提取以及运动目标行为理解等方面所表现出的近乎完美的能力,如初级视皮层对输入刺激的方向选择特性[2],使得视觉系统具有极好的轮廓感知能力,进而为后续高智能行为奠定了基础。因此,如何从人类视觉感知机制出发,构建具有神经电生理特性的神经元网络,进而对图像进行边缘检测,受到越来越多的研究人员关注。Kerr 等提出一种基于六边形感受野模型的脉冲神经网络(SNN)对图像进行边缘检测[3],与传统的矩形感受野进行比较,更符合生物特性,检测效果明显高于后者;Petkov以及Wei Hui等利用Gabor滤波器模拟神经节细胞方向选择,再结合非经典感受野抑制特性,在复杂背景图像上能有效抑制纹理噪声,提取有效边缘[4-5];Yang 等提出利用视锥细胞颜色信息处理的拮抗机制,对彩色图像进行边缘检测[6],后续进一步通过提取自然图像的方向、光强和局部光强对比等多种图像特征,对感受野输出图像施以大范围非经典感受野抑制来获取图像轮廓[7],检测效果相对单一特征的边缘检测方法优势较大;还有研究人员模拟视觉感光层功能特性,或者利用点阵神经元响应对信息进行输出编码,获取图像边缘[8-9],但弱化了视觉信号在视通路中传递时所依赖的生理结构及其功能流。因此,上述方法比传统边缘检测方法具有视觉机理上的优势,但忽略了视觉通路上不同结构对视觉信息的分工处理。所以,笔者提出一种基于突触连接视通路方位敏感机制的分级轮廓检测新方法。首先考虑视皮层下视网膜神经节细胞和外膝体的方位敏感特性,探讨其感受野最优方位在视网膜上的分布规律,利用神经元突触间的动态连接特性,构建视皮层下多方位敏感层;再结合初级视皮层更强烈的特殊方位条形光刺激感知能力,对上一功能层输出的脉冲序列进行编码,并对首发脉冲时间信息加以侧向抑制和阈值处理,得到图像边缘;最后通过重构相似度和边缘置信度对检测结果进行有效性评价,评价结果显示本方法能在边缘漏检和噪声误检之间达到较好的平衡。
1 材料和方法
1.1 视通路模型
视信号自视网膜成像形成电脉冲信号开始,直至到达视皮层完成对信号的感知与识别,视觉通道上不同的组织结构对图像轮廓特征的形成均有着不同层次的贡献。视皮层下神经节细胞和外膝体具有方位敏感特性,能感知像素梯度变化,而初级视皮层的侧向抑制作用能对脉冲信息进行有效筛选。通常认为,视网膜神经节细胞和外膝体神经元的方向敏感性感受野有着相似的结构。这里为了简化模型,笔者将神经节细胞层和外膝体层的功能合并为一层(统称为视皮层下)来对视觉输入进行处理。于是可以将视通路分为视皮层下和初级视皮层两层功能作用区,构建具有边缘特征提取能力的分级模型,如图1所示。
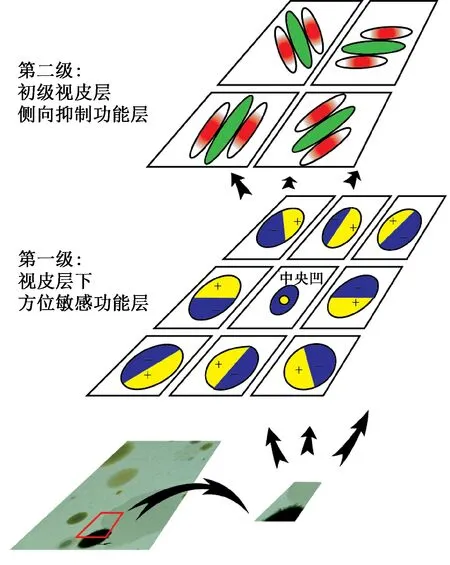
图1 视通路模型Fig.1 The model of visual pathway
1.1.1 视皮层下方位敏感
视网膜神经节细胞以及外膝体的方位敏感特性对视皮层精细的轮廓线段感知起着重要的前级安排作用[10-11]。利用神经元个体之间的动态突触连接特性,构建特定方位敏感的感受野模型,最终形成具有神经元群体边缘响应效应的网络,并对视觉输入进行差异性检测,将视觉激励映射为携带有边缘强度信息的脉冲序列。
1.1.1.1 动态突触连接
神经元是神经元网络的基本组成单元,而两个神经元发生相互作用直至形成神经元群体的互连结构称为突触。本研究选择既具备神经元电生理特性又计算简单、高效的基于漏积分放电(leaky integrate-and-fire, LIF)模型[12]来构建神经元网络。结合视觉系统感受野特性,将感受野内通过突触相连的外界激励综合作为模型输入;同时,由于后期对脉冲的编码只考虑了首次脉冲到达时间,所以去除绝对不应期。改进后的模型为
(1)
式中,v是神经元膜电压,cm是膜电容,gl是漏电导,RF是神经元感受野,wxy是突触连接权值,Ixy是激励电流,对应图像像素(x,y)处的归一化值,vth为脉冲发放阈值,vreset为静态电势。当v>vth时,神经元脉冲发放,同时v被瞬间重置为vreset。
突触在神经元与神经元之间起着信息传递的作用。神经生理学表明,这种突触传递的连接权值是动态变化的,这在一定程度上提高了神经系统对突变刺激的响应效率,而且还能充分体现空间差异信息[13]。笔者在神经元网络中引入动态突触作用时,综合考虑了神经元空间距离和输入激励差异两方面因素对突触连接强度的影响,具体表现为感受野内突触连接强度会随着距离的增大而减弱,且随着激励差异性的增大而增强,具体如下:
(2)
式中,|Pxy-Pxcyc|与|Ixy-Ixcyc|分别是周围神经元与中心神经元空间距离和激励差异,RFr是感受野半径,ΔI是感受野内最大刺激与最小刺激之差,wmax为权值调整常数。
同时,根据突触连接功能特性,上述权值会形成兴奋与抑制两种状态。从而突触连接的动态性表现为根据感受野的方位不同而呈现出兴奋与抑制两种状态之间的切换,有
(3)
1.1.1.2 方位敏感性感受野
一直以来,人们都认为只有视皮层才具有方位以及方向选择特性[14]。近年来,神经生理学研究揭示,视皮层下视网膜神经节细胞以及外膝体同样具有此特性[15],而且大多数神经元的最优方位平行于其感受野中心和视网膜中央区的连线,即呈现向心分布规律[16],如图1的第一级所示。根据此分布规律,选择围绕视网膜中央凹8个方位(0°,45°,90°,135°,180°,225°,270°,315°)上的感受野,形成4类边缘(中心水平、中心垂直、正、负对角线),并以最优方位为界,将感受野分为两个分别具有兴奋和抑制突触连接作用的区域,以实现视皮层下细胞对特定方位亮度对比线段的选择作用。以中心水平边缘(即0°和180°)为例,它会产生响应方向为向上(下暗上亮)和向下(下亮上暗)两种类型的感受野,其数学模型如下:
(4)
(5)
式中,(xc,yc)是感受野RF中心神经元坐标,(x,y)∈RF。
依次类推,可得到其他类型边缘的感受野数学模型。
1.1.2 初级视皮层侧向抑制
视觉输入经由视皮层下结构的处理,将视觉激励转化为脉冲序列供后续视觉结构处理。初级视皮层细胞接受前级结构生成的脉冲序列,以一定的形式对其进行编码[17]。考虑到信息传递的有效性,笔者选择以首次脉冲发放的时间作为编码方式[18],结合侧向抑制机制达到边缘的精确提取。
前级结构以首次脉冲时间作为输出,并映射成灰度等级,得到一幅边缘敏感图像;对边缘敏感图像上每一点(x,y),选取8方位上最大响应所对应的边缘类别,角度记为β。例如,边缘方位为中心水平,则β=0°,β∈{0°,45°,90°,135°},作为该点的最优方位,即
(6)
从而得到与该敏感图像对应的最优方位矩阵。
依据最优方位矩阵,对敏感图像引入侧向抑制作用,即感受野内的神经元处于一种竞争状态,脉冲发放时间早的神经元会抑制发放晚的神经元[19],相当于会对其产生一种延迟脉冲发放效应。对边缘敏感图像施加侧向抑制作用,有利于准确定位真实边缘,增加边缘精细程度[20]。
本研究对传统侧向抑制的作用范围加以改进,采用如图2所示的去最优方位侧向抑制方式。以3×3大小神经元感受野RF为例,将首次脉冲时间比感受野中心早的神经元定义为先发状态,反之定义为后发状态。侧向抑制的方式为:最大响应方位(垂直于最优方位OF)上的神经元若均处于后发状态(如图2中右分支所示),那么该神经元正常输出;反之,最大响应方位上只要存在一个先发状态的神经元(如图2中左分支所示),则该神经元要受到去最优方位感受野内所有处于先发状态神经元的抑制作用。

图2 去最优方位侧向抑制方式Fig.2 Sketch map of removing optical direction lateral inhibition
一般认为,侧向抑制作用大小受神经元空间距离、神经元放电活动和最优方位3个要素的影响。神经元空间距离越大,突触连接强度越小,侧向抑制量也越小;而神经元放电时间差越小、最优方位越相似,表明该处存在冗余边缘的可能越大,故抑制量也越大。所以本研究将侧向抑制系数定义如下:
(7)
式中:Δd为两神经元间的空间距离;RFr是感受野半径;Txy和Txcyc分别是周围和中心神经元首次脉冲的到达时间;δ是最优方位最小差,等于45°;θ是两神经元最优边缘方位的角度差,定义为
(8)
式中,σxy和σxcyc分别是周围和中心神经元最优边缘方位所对应的角度。
1.2 检测过程
利用动态突触构建的多方位敏感型感受野对输入图像进行边缘强度检测,以首发脉冲时间进行编码,映射形成边缘敏感图像;然后引入改进的侧向抑制方法,对敏感图像进行有选择性的抑制处理,再经过阈值处理形成最终边缘。程序执行流程如图3所示,具体检测过程如下所述。
步骤1:根据原始图像Io(x,y)(x=1,2,…,M;y=1,2,…,N)大小构建相同尺寸的神经元网络GC(x,y),各个神经细胞在其感受野RFxy,gc(5×5)内以突触的形式与周围神经元互连,依式(2)计算突触连接权值wxy的大小。
步骤2:根据式(4)、(5)以及其他方位边缘的感受野数学模型,将步骤1中得到的权值wxy赋予兴奋和抑制功能,形成8个对不同方位敏感的感受野内突触连接权值,记为wxy,α,α=0,45,90,135,180,225,270,315。
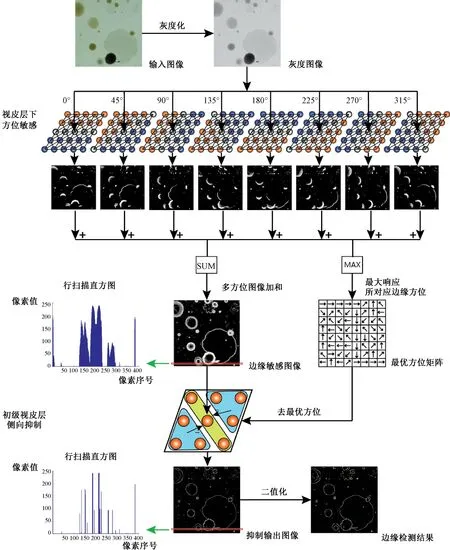
图3 边缘检测算法流程Fig.3 The flow diagram of image edge detection
步骤3:对每个神经细胞(x,y)采用式(1)所示的LIF神经元模型,并将原始图像像素值归一化,以激励电流的形式输入神经元,分别将步骤2获得的8个突触连接权值代入该模型。
步骤4:记录神经元网络GC(x,y)在不同类型的方位敏感性感受野作用下,各个神经元首次脉冲的达到时间,从而得到8个时间矩阵T1st,α(x,y)。
步骤5:利用式(9)融合多方向上的时间矩阵,并映射成灰度等级,得到视网膜前级安排作用下的边缘敏感图像,有
(9)
式中,Tmax是融合后的时间矩阵∑αT1st,α(x,y)的最大值。
另外,根据式(6)计算得到与原始图像尺寸相同的最优方位矩阵D(x,y)。
步骤6:再次构造初级视皮层细胞感受野RFxy,vc(3×3),由图2所示规则确定神经元是否受侧抑制作用及其在受到侧向抑制时的作用范围,再根据式(7)、(8)计算侧抑制系数wxy,vc,然后计算得到视皮层精细边缘提取结果IT(x,y),有
(10)
式中,ITxy和ISxy分别是IT(x,y)和IS(x,y)在(x,y)处的值。
步骤7:对步骤6的结果进行阈值处理,得到最终边缘图IE(x,y),有
(11)
式中,IExy是IE(x,y)在(x,y)处的值,Ith是设定阈值。
1.3 算法验证

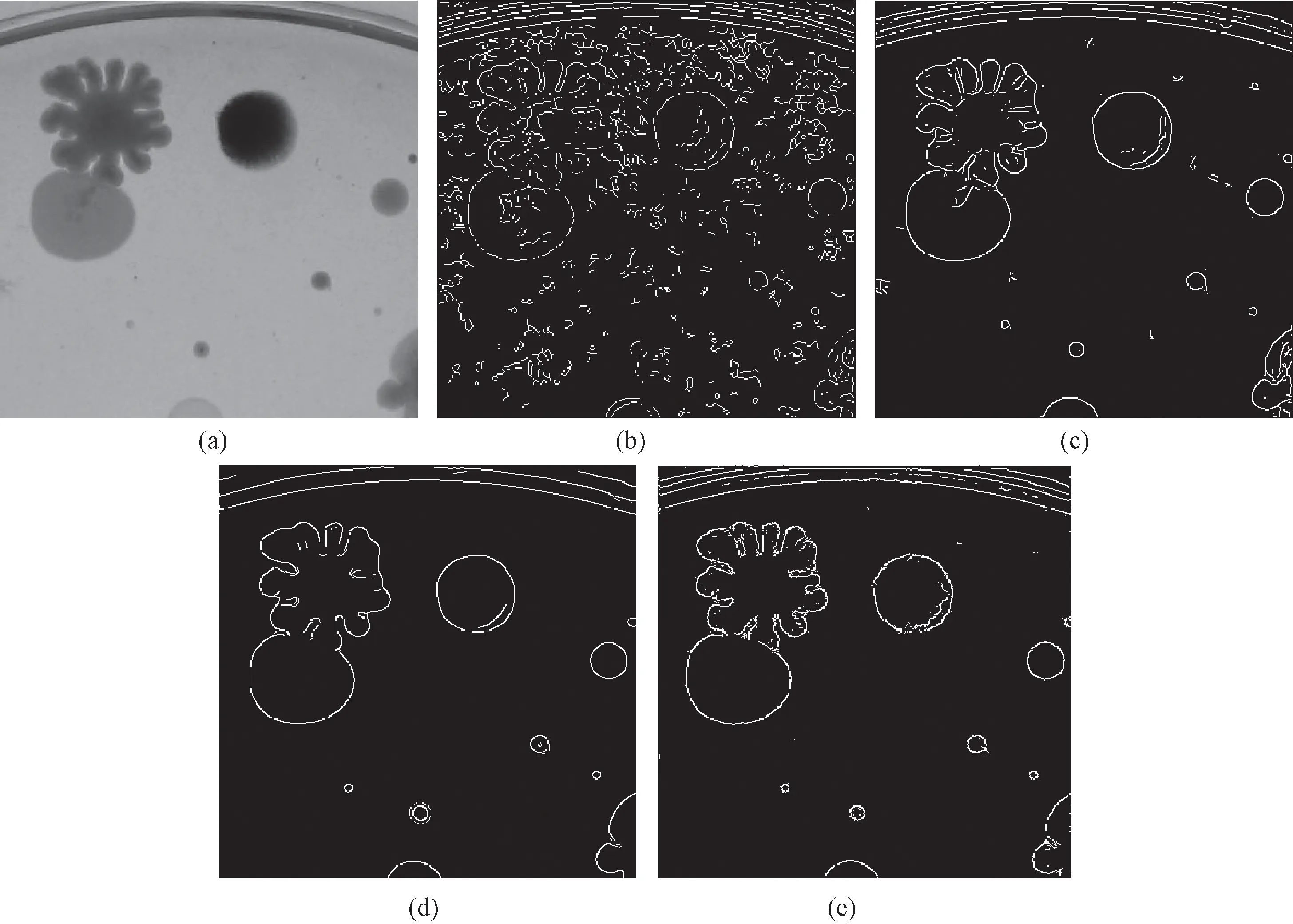
图4 菌落图像。(a)多强度边缘菌落;(b)外光环深层菌落;(c)形态丰富菌落;(d)粘连菌落Fig.4 Bacterial colony image for later processed. (a) Colony image with multi-intensity edge; (b) Deep colony image with outer ring; (c) Colony image with rich form; (d) Adhesive colony image
菌落图像的边缘检测是用图像方法分析菌落的基本步骤之一,菌落图像具有细节丰富、层次模糊、形态各异等特征,对其进行边缘检测存在一定难度。本算法有着准确定位强弱边缘、抑制纹理噪声以及保护弱小细节等优点,从原理上能较好地克服上述困难。本实验室常规微生物实验需要选取采集的12幅典型菌落图像作为实验用图,部分图像如图4所示。另外,选取Canny、具有方向选择特性的Gabor滤波器方法以及文献[7]的方法(MCI),与所提出的算法进行效果比较,验证本算法的有效性。其中,Gabor滤波器选取了16方向,MCI方法中Gaussian函数设置为8方向。
为客观地对检测结果进行评价,利用边缘检测结果对原图进行重构[21],再计算重构图像相似度指标MSSIM[22]和边缘置信度指标BIdx[23],从而定量验证本算法的有效性。同时,考虑到MSSIM和BIdx在一定程度上可以形成互补关系,且均依赖于边缘像素点数量EdgeNum,因此以两者的加权和作为综合指标EIdx[24],评价不同算法的优劣程度,即

图5 图4(a)边缘检测结果。(a)图4(a)的灰度图;(b) Canny方法;(c) Gabor滤波器方法;(d) MCI方法;(e)本方法Fig.5 Edge maps of Fig.4(a). (a) Gray image of Fig.4(a); (b) Canny method; (c) Gabor method; (d) MCI method; (e) Proposed method
(12)
式中,M×N是图像大小,由于本研究的菌落图像中弱边缘以及细节信息较多,所以0.5 最后,对所选取的12幅菌落图像的EIdx指标作t检验,将上述3种对比方法分别与本方法进行比较,P<0.05时被认为具有显著性差异。 图6 图4(b)边缘检测结果。(a)图4(b)的灰度图;(b) Canny方法;(c) Gabor滤波器方法;(d) MCI方法;(e)本方法Fig.6 Edge maps of Fig.4(b). (a) Gray image of Fig.4(b); (b) Canny method; (c) Gabor method; (d) MCI method; (e) Proposed method 图7 图4(c)边缘检测结果。(a) 图4(c)的灰度图;(b) Canny方法;(c) Gabor滤波器方法;(d) MCI方法;(e)本方法Fig.7 Edge maps of Fig.4(c). (a) Gray image of Fig.4(c); (b) Canny method; (c) Gabor method; (d) MCI method; (e) Proposed method 图8 图4(d)边缘检测结果。(a) 图4(d)灰度图;(b) Canny方法;(c) Gabor滤波器方法;(d) MCI方法;(e)本方法Fig.8 Edge maps of Fig.4(d). (a) Gray image of Fig.4(d); (b) Canny method; (c) Gabor method; (d) MCI method; (e) Proposed method 图5~8分别是对图4所示4幅菌落图像进行边缘检测后的结果。观察这些结果,可以发现本方法对这些菌落图像具有更好的边缘信息提取能力。分析各子图(b)所示的Canny方法检测结果:当菌落图像的丰富细节信息得到很好的检测时,会出现大量由纹理噪声形成的伪边缘,如图5(b)、图6(b)和图7(b)均出现这样的现象,说明Canny方法很难在弱边缘和纹理噪声之间达到平衡。分析各子图(c)所示的Gabor滤波器方法检测结果:对小的菌落会存在边缘断裂、不完整现象,这在一定程度上又增加了虚假边缘,如在图6(c)和图8(c)中均有所体现。分析各子图(d)所示的MCI方法检测结果:虽然纹理噪声得到了很好的抑制,但基本上只检测出菌落的大致轮廓,细节丢失较为严重,如图7(d)触角形态的边缘很多都连在一起。相比之下,各子图(e)所示的本方法对各类菌落图像都具有相对完整的检测效果,不仅能在不造成虚假边缘的条件下对多强度边缘图像(见图5(e))提取有效的边缘轮廓,而且包含大量细节信息图像(见图6(e)、图7(e))的边缘提取效果也优于其他方法。 为了对检测结果进行定量评价,利用边缘检测结果对原图进行重构,重构结果如图9所示。计算该图与原图的MSSIM指标,计算结果见表1左侧;再计算BIdx指标,计算结果见表1右侧。可以看 出,本方法的重构相似度均显著大于其他方法的重构相似度,这说明其对于原图边缘的完整性有很好的保留,边缘信息丰富;本方法的边缘置信度也基本比前3种方法好,表明其检测出来的边缘质量好,真实度高。另外,Canny方法的重构指标明显要优于Gabor滤波器方法和MCI方法的重构指标,而其边缘置信度却不如后两种方法的边缘置信度,所以有必要使用二者的综合指标EIdx对结果进行评价。 表1 4种边缘检测方法的重构图像相似度和边缘置信度 图9 基于菌落边缘轮廓重构图像(每行从左至右分别对应基于Canny、Gabor、MCI方法以及本方法所获得的边缘重构得到的图像)。(a)~(d)分别对应于图4中的(a)~(d)Fig.9 Reconstructed images based on the edge of colony using different method(the maps from left to right in every row correspond to Canny method, Gabor method, MCI method and proposed method).(a)~(d) correspond to Fig.4(a)~Fig.4(d) 对12幅菌落图像(其中前4幅如图4所示)计算其在不同方法下边缘检测的EIdx指标,并统计该指标均值及标准差,统计结果见表2。可以看出,Canny、Gabor以及MCI方法的综合指标均值分别为0.520 8、0.522 5和0.524 4,而本方法所对应的数据为0.746 8。这表明,本方法从整体上来看也是具有较好的边缘检测效果,不仅完整检测出边缘细节信息,而且真实度高。另外,对这12幅菌落图像的综合指标做t检验,结果显示3种对比方法与本方法数据的P值均小于0.01,表明本方法与对比方法的综合指标具有显著性差异。 表2 4种边缘检测方法综合指标的均值及标准差(均值±标准差) 通过以上实验结果不难看出,本方法具有较好的边缘提取能力,特别是针对具有弱对比度边缘且细节丰富的图像效果更好。从表1所展示的重构图像相似度和边缘置信度两项指标,能反推本算法的优势所在。 首先,分析表1左侧重构图像相似度评价指标。重构相似度是基于亮度、对比度和结构三者综合,满足人类视觉感官的图像相似度指标。笔者在构建视皮层下方位敏感功能层时,考虑了视皮层下视网膜神经节细胞以及外膝体神经元最优方位的向心分布规律,利用突触动态连接特性,形成多方位敏感的感受野模型,能更大程度地保留图像边缘细节信息。另外,对相同类型的边缘,如水平边缘,笔者计算响应方向为向上和向下时的边缘响应,分别对应下暗上亮和下亮上暗两种情形的图像边缘,更进一步地保证图像轮廓的完整性。从相似度评价指标的结果来看,本算法数据均在0.8以上,优于其他对比方法,同时,通过图9所示图像重构结果,也不难看出该评价结果与定性的观测结果是吻合的。 其次,分析表1右侧边缘置信度评价指标。边缘置信度是利用邻域相对标准差来说明所检测边缘的真实可靠性。本研究在构建初级视皮层侧向抑制功能层时,利用了前层神经元脉冲输出的先后次序设定抑制规则。图像边缘通常会形成像素梯度变化的区域,而该变化中总会有一个最大的变化即为真实边缘所在。在最大响应方位上有选择地进行侧向抑制,不仅有利于去除伪边缘和纹理噪声,而且能保护真实边缘不受破坏,进而准确定位;对抑制区域去除最优方位,能够避免边缘之间自抑制,保护弱小细节,还原更真实的图像轮廓。故本方法能在拥有高相似度指标的情况下,还能保证边缘置信度指标也优于对比方法。与之相反的是,Canny算法虽然也拥有较好的相似度指标,但边缘可靠性不高,引入大量纹理噪声,故置信度指标明显低于其他方法。分析其原因,这是由于重构图像的质量会随算法检测出的边缘数量增加而提高,Canny方法检测出来的边缘数量较多,其中不仅包含真实边缘,也包含大量虚假边缘,从检测结果图5~8的子图(b)中也很容易看出其误检较多。 综合来看,Canny算法通过高低阈值来选取确定最终的边缘,不能有效区分细节信息和纹理噪声,稳定性较差;MCI方法虽然考虑了非经典感受野抑制作用,使重构相似度和边缘置信度有很好的平衡,表现出较好的稳定性,但大范围的非经典感受野抑制使主要菌落边缘更清晰的同时丢失了大量细节;Gabor滤波器方法虽然也考虑了方向敏感特性,但没有神经元之间的侧向抑制作用,导致大量伪边缘,而且边缘连续性上表现较差。综上所述,这三种方法都不适合于检测层次模糊而细节丰富的图像边缘。此外,文献[8]的方法检测结果边缘较粗,准确性不高,重构相似度为0.8左右,也与本方法的结果存在差距。相反,本方法不仅能准确在纹理噪声中区分出弱边缘和细节信息,保证边缘完整性,而且还不失真实性,优势较大。 笔者通过讨论视觉通路上神经节细胞、外膝体以及初级视皮层在轮廓特征提取过程中的具体分工,探讨视皮层下细胞感受野分布与最优敏感方位之间的关系,同时引入初级视皮层在信息筛选过程中的脉冲编码和侧向抑制机制,摒弃采用黑箱形式的数学模型对视觉机制进行模拟的思路,提出一种基于突触连接皮层下和皮层上分级处理的图像边缘检测方法,并与Canny、Gabor滤波器以及MCI等方法进行分析比较,表明所提出的方法检测出的图像边缘细节更加丰富,且不引入纹理噪声。菌落图像的边缘检测实验结果表明,通过视皮层下功能层凸显边缘强度信息,再通过初级视皮层功能层进行有效信息筛选,并对微弱信息进行加强,不但能降低纹理和噪声对真实轮廓的干扰,还能加强细节信息,从而获得更好的整体效果。本研究提出的基于突触连接视通路方位敏感的图像分级边缘检测方法,将为图像轮廓检测技术提供一个新的思路,后续可研究用该方法提取的图像轮廓特征在图像重现等图像理解问题中的应用。 [1] Maini R, Aggarwal H. Study and comparison of various image edge detection techniques [J]. International Journal of Image Processing (IJIP), 2009, 3(1): 1-11. [2] Sadagopan S, Ferster D. Feedforward origins of response variability underlying contrast invariant orientation tuning in cat visual cortex [J]. Neuron, 2012, 74(5): 911-923. [3] Kerr D, Coleman S, Mcginnity M,etal. Biologically inspired edge detection [C] //2011 11th International Conference on Intelligent Systems Design and Applications (ISDA). Cordoba: IEEE, 2011: 802-807. [4] Petkov N, Subramanian E. Motion detection, noise reduction, texture suppression, and contour enhancement by spatiotemporal Gabor filters with surround inhibition [J]. Biological Cybernetics, 2007, 97(5-6): 423-439. [5] Wei Hui, Lang Bo, Zuo Qingsong. Contour detection model with multi-scale integration based on non-classical receptive field [J]. Neurocomputing, 2013, 103(0): 247-262. [6] Yang Kaifu, Gao Shaobing, Li Chaoyi,etal. Efficient color boundary detection with color-opponent mechanisms [C] //2013 IEEE Conference on Computer Vision and Pattern Recognition (CVPR). Portland: IEEE, 2013: 2810-2817. [7] Yang Kaifu, Li Chaoyi, Li Yongjie. Multi-feature based surround inhibition improves contour detection in natural images [J]. IEEE Transactions on Image Processing, 2014, 23(12): 5020-5032. [8] 罗佳骏,武薇,范影乐,等. 基于视觉感光层功能的菌落图像多强度边缘检测研究 [J]. 中国生物医学工程学报, 2014, 33(6): 677-686. [9] 廖进文,范影乐,武薇,等. 基于点阵神经元响应时空信息的菌落图像边缘检测 [J]. 航天医学与医学工程, 2014, 27(2): 94-100. [10] Rivlin-Etzion M, Wei Wei, Feller MB. Visual stimulation reverses the directional preference of direction-selective retinal ganglion cells [J]. Neuron, 2012, 76(3): 518-525. [11] Naito T, Okamoto M, Sadakane O,etal. Effects of stimulus spatial frequency, size, and luminance contrast on orientation tuning of neurons in the dorsal lateral geniculate nucleus of cat [J]. Neuroscience Research, 2013, 77(3): 143-154. [12] Chouhan AS. An analytical study of leaky integrate-and-fire neuron model using MATLAB simulation [J]. Int J Eng Res Technol, 2013, 2(4): 2242-2245. [13] Venkataramani S, Taylor WR. Orientation selectivity in rabbit retinal ganglion cells is mediated by presynaptic inhibition [J]. The Journal of Neuroscience, 2010, 30(46): 15664-15676. [14] 寿天德. 视觉信息处理的脑机制 [M]. 2版. 合肥: 中国科学技术大学出版社, 2010. [15] Zhao Xinyu, Chen Hui, Liu Xiaorong,etal. Orientation-selective responses in the mouse lateral geniculate nucleus [J]. The Journal of Neuroscience, 2013, 33(31): 12751-12763. [16] Suematsu N, Naito T, Miyoshi T,etal. Spatiotemporal receptive field structures in retinogeniculate connections of cat [J]. Frontiers in Systems Neuroscience, 2013, 7(103): 1-13. [17] Tanabe S. Population codes in the visual cortex [J]. Neuroscience Research, 2013, 76(3): 101-105. [18] Gollisch T, Meister M. Rapid neural coding in the retina with relative spike latencies [J]. Science, 2008, 319(5866): 1108-1111. [19] Isaacson JS, Scanziani M. How inhibition shapes cortical activity [J]. Neuron, 2011, 72(2): 231-243. [20] Zi Fang, Zhao Dawei, Zhang Ke. Image pre-processing algorithm based on lateral inhibition [C] // The 8th International Conference on Electronic Measurement and Instruments. Xi’an: IEEE, 2007: 2-701-2-705. [21] Govindarajani B, Panetta KA, Agaian S. Image reconstruction for quality assessment of edge detectors [C] //IEEE International Conference on Systems, Man and Cybernetics. Singapore City: IEEE, 2008: 691-696. [22] Wang Zhou, Bovik AC, Sheikh HR,etal. Image quality assessment: from error visibility to structural similarity [J]. IEEE Transactions on Image Processing, 2004, 13(4): 600-612. [23] Meer P, Georgescu B. Edge detection with embedded confidence [J]. IEEE Transactions on Pattern Analysis and Machine Intelligence, 2001, 23(12): 1351-1365. [24] 磨少清. 边缘检测及其评价方法的研究 [D]. 天津: 天津大学, 2011. A Hierarchical Image Edge Detection Method Based on Orientation Sensitivity of Visual Pathway with Synaptic Connections Wang Dian Fan Yingle*Zhang Mengnan Wu Wei (LaboratoryofPatternRecognitionandImageProcessing,HangzhouDianZiUniversity,Hangzhou310018) The orientation sensitivity of human visual pathway plays a key role in contour perception, and this feature provides vital information for image understanding. In this paper, a new method of image edge detection based on visual direction sensitive mechanism was proposed. Using the physical structure feature of ganglion cells and LGN neurons receptive field distributing centripetal, a sub-cortex multi-direction sensitive function layer was constructed to transform visual incentive to pulse sequence, and neural spiking information were fused to get an edge sensitive image; then a primary visual cortex function layer with removing optical direction receptive field was built to code on the spike sequence generated by the former layer according to first spike time. The edge detection result was obtained through lateral inhibition and threshold processing. In this paper, colony images with fuzzy hierarchy and rich details were taken for processing. The results of hierarchical edge detection were assessed by the confidence of edge, reconstruction similarity and weighted sum of them. It was proved that our method can completely detect image edge and effectively filter out texture noise. And the mean value of weighted sum index was 0.7468, significantly higher than other methods compared. The new method of edge detection proposed in the paper provides a new idea for the image processing and understanding based on orientation sensitivity of visual pathway. edge detection; visual pathway; synaptic connections; orientation sensitivity; lateral inhibition 10.3969/j.issn.0258-8021. 2015. 05.002 2015-04-08, 录用日期:2015-05-25 国家自然科学基金(60872090);浙江省大学生科技创新活动计划项目(2014R407013) R318 A 0258-8021(2015) 05-0522-11 *通信作者(Corresponding author), E-mail: fan@hdu.edu.cn2 结果





3 讨论
4 结论
