中亚热带6种不同更新方式森林0—5mm根系直径频率分布
2015-02-11胡双成熊德成黄锦学王韦韦黄超超陈云玉陈光水
胡双成,熊德成,*,黄锦学,王韦韦,黄超超,邓 飞,陈云玉,陈光水
1 湿润亚热带山地生态国家重点实验室培育基地, 福州 350007 2 福建师范大学地理研究所, 福州 350007
中亚热带6种不同更新方式森林0—5mm根系直径频率分布
胡双成1,2,熊德成1,2,*,黄锦学1,2,王韦韦1,2,黄超超1,2,邓 飞1,2,陈云玉1,2,陈光水1,2
1 湿润亚热带山地生态国家重点实验室培育基地, 福州 350007 2 福建师范大学地理研究所, 福州 350007
运用土芯法研究福建省三明市6种不同更新方式林分(米槠天然林、米槠天然更新林、米槠人促更新林、米槠人工林、马尾松人工林、杉木人工林)根系的(直径≤5 mm)根长随直径分布特点。结果表明:0—5 mm直径根系的根长87%—98%集中于0—2 mm直径范围内,65%—88%集中于0—1 mm直径范围内。6种不同更新方式林分的根长直径分布均呈单峰型分布,且均可用对数正态分布累积函数进行较好的拟合,R2均达0.99以上。函数中拟合参数μ值和σ值的变化代表了不同林分细根在资源获取策略中的差异。拟合参数μ值和σ值之间具有很好的负相关,表明这两个参数呈现协同性变化,反映了细根资源获取策略中的权衡关系。随树种多样性的增加,μ值趋于更小而σ值更大,可能体现了树种间根系对养分和水分竞争的加剧;不同树种人工林μ值、σ值亦存在明显分异,反映了不同树种土壤资源利用策略的差异。结论表明对数正态分布累积函数可以很好地反映林分尺度的根系直径频率分布特点及根系资源利用策略。
森林;细根;根长;直径频率分布;土壤资源利用策略;对数正态分布
植物根系,特别是细根(通常<2 mm)是植物吸收水分和养分的主要器官[1],虽然只占根系总生物量的3%—30%[2],但其生长、死亡和周转在陆地生态系统C分配和养分循环过程中却发挥了重要作用。直径是根系的重要结构特征,其粗细具有不同的生理功能[3-5]。分析不同根系功能指标随直径的变化,对于认识细根结构与功能的关系、建立合理可靠的根系功能分类方法等具有重要意义[6]。然而,按不同直径给细根分类却很少被关注[7]。目前,细根的直径定义饱受批评[6,8],主要因为该定义把特定径级(如<2 mm)细根当作均质功能单元,而忽视细根功能随细根直径的变化[9]。细根周转的准确估计一直是地下生态学研究的难点之一[10-11]。由微根管法获得细根周转速率和由土芯法获得的细根生物量两者的乘积被推荐为目前最合适的细根周转估计方法[12-14]。然而由于两种方法自身存在的缺陷,导致两种方法所观测的细根群体间的不匹配,如微根管法观测的细根直径较土芯法获得的细根偏小[15]。因而,这种方法需要建立在相同的细根群体之上。由于细根直径的测定比细根序级或细根模块的测定更为便携,因而根系直径频率分布(如细根周转速率和生物量随直径的分布)可以为两种方法的整合提供相同的标准,从而对准确估计细根周转具有重要的作用。根长是根系功能的重要指标,根系的直径频率分布携带了非常丰富和有用的信息。如根长的直径频率分布已被作为了解根的形态、养分吸收情况、耕作方式对根系的影响、根之间的相互作用和菌根真菌的入侵的一种有效途径。同时,根长的直径频率分布亦是把根结构、根形态和根生物量纳入到分析模型中的一种简单有效的方法[16]。因而,研究根系的直径频率分布对于深入了解不同森林或不同植物根系的资源获取策略、根系与土壤环境相互作用和模型预测等有十分重要的作用。
然而,目前有关根系直径频率分布研究大多在单株水平或盆栽条件下开展[17-18],而很少研究林分水平的根系直径频率分布特征;湿润亚热带森林的根系直径频率分布特征的研究亦很少。不同森林更新方式常可以形成具有不同树种组成的次生林或人工林,但有关树种组成(包括树种多样性和树种特性)如何影响根系直径频率分布目前亦不清楚。
本研究通过选择福建省三明市由米槠(Castanopsiscarlesii)天然林经不同更新方式形成的年龄大致相近(约40a)的次生林和人工林,包括经天然更新形成的米槠天然更新次生林;经人促天然更新形成的米槠人促次生林;以及经人工造林形成的米槠人工林、杉木人工林、马尾松人工林,来研究不同的树种多样性和树种特性对根系直径频率分布的影响,为探讨更新方式对森林根系资源利用策略的影响提供依据。
1 研究地区与研究方法
1.1 研究区概况
研究区位于福建省三明市金丝湾森林公园陈大林业采育场和三明格式栲(Castanopsiskawakamii)自然保护区。该区东南面和西北面分别与戴云山脉和武夷山脉相连,地貌以低山丘陵为主。气候属中亚热带季风气候。土壤以花岗岩发育的红壤和黄壤为主,还分布有红黄壤、黄棕壤等,多呈酸性,土壤厚度>1 m。区域内分布着以米槠为代表的中国面积最大的常绿阔叶林,物种多样性十分丰富。
所选树种均为地区代表性乔木,分别为同树种不同更新方式的米槠天然林、米槠天然更新林、米槠人促更新林、米槠人工林和不同树种的马尾松人工林以及杉木人工林。4种米槠林分除米槠天然林外,其它3种林龄相似,且立地条件(如岩性、土壤类型、地形和气候条件等)相同。米槠天然更新林和米槠人促更新林分布于陈大林业采育场内,米槠人工林位于格氏栲自然保护区内,两地相距小于30km。米槠天然更新林由米槠天然林经过强度择伐后封山育林,经过次生演替天然更新形成。其乔木层主要树种有米槠、闽粤栲(Castanopsisfissa)、新木姜子(Neolitseaaurata)和黄丹木姜子(Litseaelongata)等组成;灌木层主要由罗浮栲(Castanopsisfabri)、木荚红豆(Ormosiaxylocarpa)、褐毛石楠(Photiniahirsuta)、米槠等组成;草本层主要由黑莎草(Gahniatristis)、油草(Leptochloachinensis)、狗脊蕨(Woodwardiajaponica)等组成。米槠人促更新林是由米槠天然林经过强度择伐后天然更新,并在更新的过程中,定期砍除生长较差的非目的树种和杂灌,保留生长较快的树种。其乔木主要树种主要有米槠、木荷(Schimasuperba)和东南野桐(Malloyuslianus)等;灌木层主要由毛叶冬青(Ilexpubilimba)、鼠刺(Iteachinensis)和石栎(Lithocarpusglaber)等组成;草本层不发达,主要由鳞子莎(Lepidospermachinense)、狗脊蕨、扇叶铁线蕨(Adiantumflabellulatum)等组成。米槠人工林是经过皆伐、火烧、挖穴造林和幼林抚育形成。其乔木为单一树种米槠;林下灌木层主要以毛冬青(Ilexpubescens)、乌饭树(Vacciniumbracteatum)、桂北木姜子(Litseasubcoriacea)、薄叶山矾(Symplocosanomala)和山姜(Alpiniajaponica)等占优势;草本层以芒萁(Dicranopterisdichotoma)占优势。杉木人工林(Cunninghamialanceolata)和马尾松(Pinusmassoniana)人工林均为1976年次生林皆伐后,营造人工纯林形成。杉木人工林的林下植被主要以狗骨柴(Tricalysiadubia)、毛冬青(Ilexpubescens)、芒萁为主。马尾松人工林的林下植被覆盖度约为90%,灌木较少,主要以芒萁、芒(Miscanthussinensis)和蕨(Pteridiumaquilinum)等草本为主。这些不同的森林类型构成了不同的人为干扰梯度和生物多样性梯度,为研究提供了良好的天然试验地[19]。不同更新方式各试验地详细概况参见表1。
1.2 研究方法
1.2.1 根系取样
中亚热带地区细根生物量一般在春季(3—5月)达到最大值[20-22],故选择在2011年4月进行细根取样。在每个林分类型中,依上、中、下坡位分别设置3块20m×20m样地,用内径4.05 cm的土钻按S形钻取土芯,深度为80cm。6个样地分别钻取6、6和8个土芯,每种林分均为20个土芯。按0—10cm、10—20cm、20—40cm、40—60cm和60—80cm这5个间隔把土芯分割为5个土层,每个土层样品分别装入标记好的塑料袋中,带回实验室并存放于4 ℃冰箱内。
将土样从塑料袋中取出,在盆中用自来水浸泡、漂洗、待根与土壤分离后过0.5 mm尼龙筛。用镊子、剪刀、游标卡尺等工具按小于1 mm、1—2 mm和大于2—5 mm三个径级分捡根,其中大于5 mm的根舍去。根据根颜色、外形、弹性、根皮与中柱分离的难易程度挑出活根。鉴于用单一径级定义细根且认为所有<2 mm的根在结构和生理上都相同的认识忽视了根系内部结构和功能上的异质性[23],在估计细根周转时可能造成较大误差[24],也不利于认识根系内部结构根和吸收根之间的变化,故而本研究选取了0—5 mm根系作为研究对象。由于本研究是群落水平的物种根系研究,因此不对各物种的细根进行具体区分。
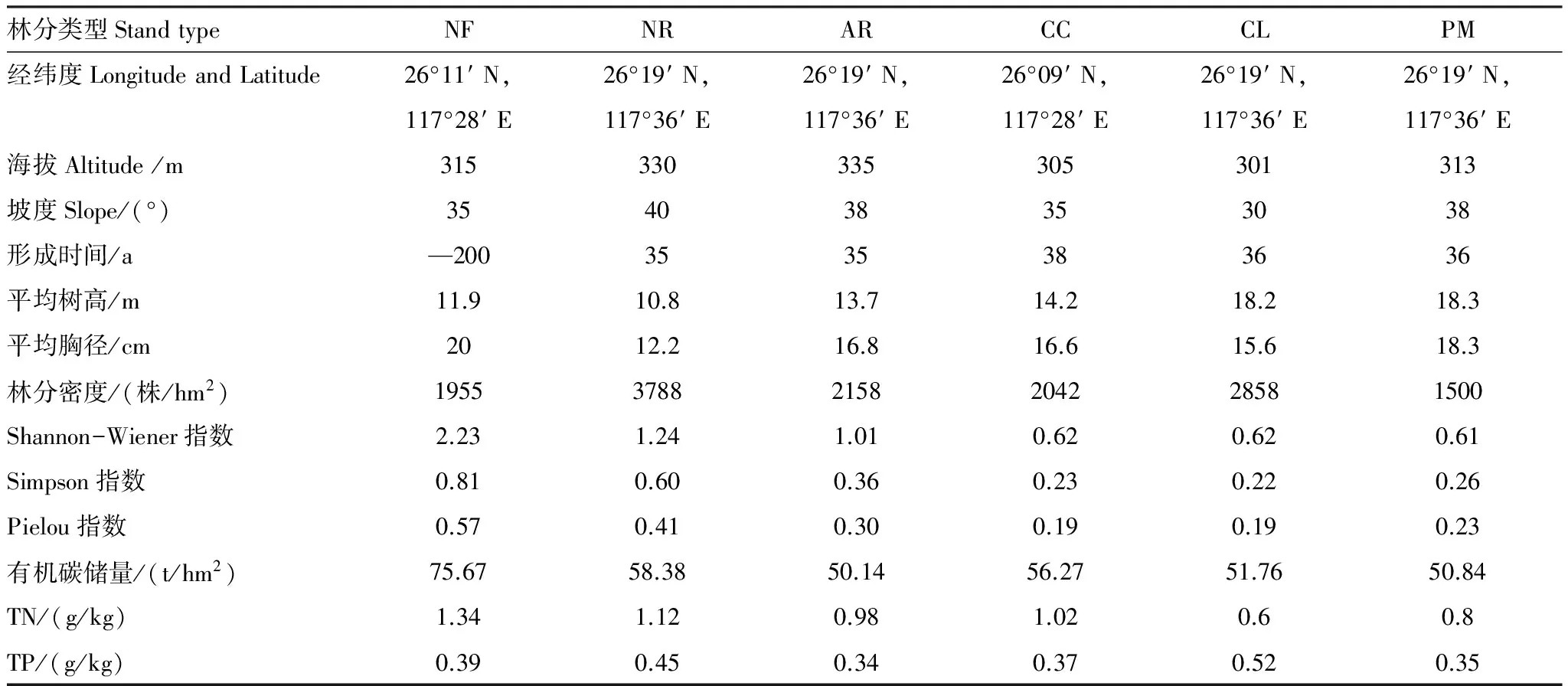
表1 研究区6种林分的基本特征和土壤表层(0—20cm)理化性质Table 1 Main stand characteristic and top soil (0—20cm) properties in the six studied stands
NF:米槠天然林 Nature forest ofCastanopsiscarlesii; NR:米槠天然更新林Secondary forest ofCastanopsiscarlesiithrough natural regeneration; AR:米槠人促更新林 Secondary forest ofCastanopsiscarlesiithrough natural regeneration with anthropogenic promotion; CC:米槠人工林Castanopsiscarlesiiplantation; PM:马尾松人工林Pinusmassonianaplantation; CL:杉木人工林Cunninghamialanceolataplantation; 形成时间 formation time; 平均树高 Mean tree heigh; 平均胸径 Mean tree diameter at breast height; 林分密度Stand density; Shannon-Wiener指数 Shannon-Wiener index; Simpson指数 Simpson index; Pielou指数 Pielou index; 有机碳储量 Soil organic carbon reserve; TN:全氮Total nitrogen; TP:全磷Total phosphorus
1.2.2 根系指标测定
将处理好的根系样品放置在扫描仪的透明托盘中,注入水,使根系完全舒展,采用数字化扫描仪Epson scanner对根系样品进行扫描。使用Win-RHIZO(Pro 2009b)根系扫描分析系统对根系图像进行扫描,得到根系的长度、表面积和根长分布等数据。再将根样品放置于65℃烘箱中烘干48h至恒重后称干重。
计算细根根长密度(RLD) 、生物量密度(RBD)数据。计算公式如下:
根长密度(m/m3)=每土芯平均细根长×10-6/ [π(d/2)2×h];
生物量密度(g/m3)=每土芯平均细根干重×10-6/[π(d/2)2×h];
式中,d为使用土钻的内径(cm),h为所用土钻的高度(cm)。
在Win-RHIZO(Pro 2009b)根系扫描分析系统扫描后的结果中,把细根直径以0.2 mm间隔依次分成25个间隔(即0—0.2 mm,0.2—0.4 mm,…,4.6—4.8 mm,≥4.8 mm),以此来计算6种林分根系的直径频率分布。
1.3 数据处理
利用SPSS 17. 0软件对数据进行统计分析。采用单因素方差分析和最小显著差数法(LSD)检验各径级范围不同树种的根长密度和生物量密度的差异,显著性水平设定为α=0.05。采用非线性回归中的对数正态分布累积函数(CDF log-normal):
(1)
式中,y为根系在直径为x时的累积根长比重;μ、σ为函数的拟合参数。
对6种不同林分根系的累积直径频率分布建立模型,拟合出各林分的参数μ和σ,并得出预测值。利用决定系数R2对以上回归方程的拟合程度进行F检验。采用Excel 2007软件作图。
2 结果与分析
2.1 根长密度和生物量密度
由表2可知,6种林分中,0—1 mm细根和0—2 mm细根的根长密度占0—5 mm根系根长密度的比值从米槠天然林到米槠天然更新林、米槠人促更新林、米槠人工林、马尾松林和杉木林分别为:87.8%、83.3%、65.0%、81.3%、79.3%、81.8%和97.7%、92.8%、87.2%、94.8%、92.6%、95.3%。这说明0—2 mm,特别是0—1 mm细根在总的根长密度中占主体。然而,从米槠天然林到米槠天然更新林、米槠人促更新林、米槠人工林、马尾松林和杉木林各林分0—2 mm细根生物量密度的比值占0—5 mm细根生物量密度分别为60.4%、48.0%、48.7%、64.0%、49.4%和58.5%。从米槠天然林、米槠天然更新林、米槠人促更新林到米槠人工林,3个径级范围的根长密度和根生物量密度均显著降低,表明随着人为干扰程度的增加,细根生产力降低。在3种人工林,以杉木林的根长密度和生物量密度最大,而马尾松林的根长密度和生物量密度则最小。
6种林分中,米槠天然林和杉木林的根长密度在3个径级范围内,都与其它林分间差异显著。杉木林的生物量密度在0—1 mm和0—2 mm径级范围内与其它林分间差异显著。其它各林分各径级间关于根长密度和生物量密度的差异性见表2。
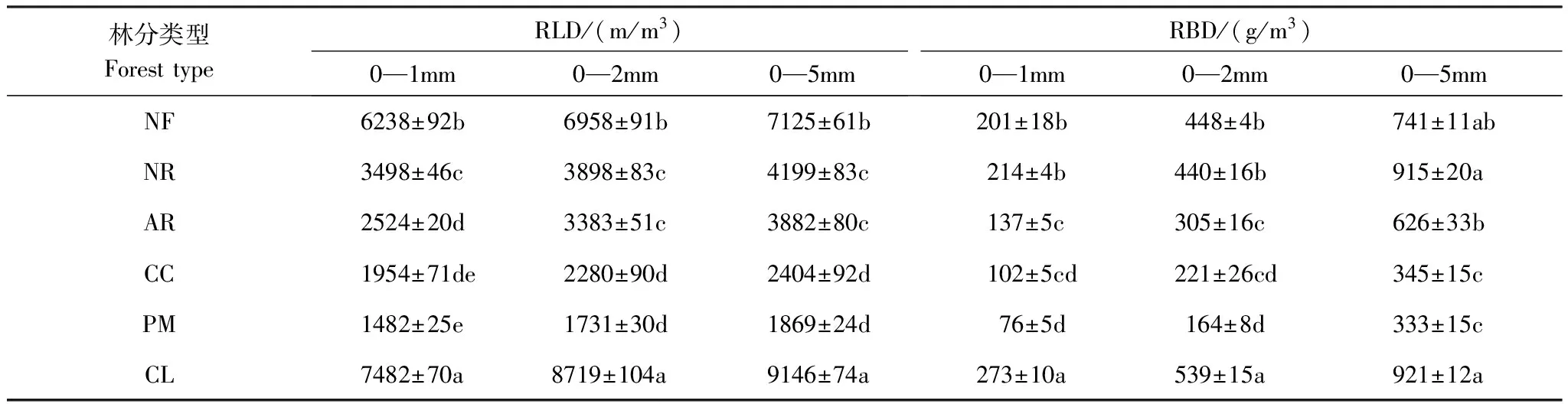
表2 6种林分不同径级根系的根长密度(RLD)和生物量密度(RBD)Table 2 the RLD and RBD of different diameter roots in the six stands
同一列相同小写字母表示两个林分间无显著差异(P﹥0.05); NF:米槠天然林Nature forest ofCastanopsiscarlesii; NR:米槠天然更新林Secondary forest ofCastanopsiscarlesiithrough natural regeneration; AR:米槠人促更新林 Secondary forest ofCastanopsiscarlesiithrough natural regeneration with anthropogenic promotion; CC:米槠人工林Castanopsiscarlesiiplantation; PM:马尾松人工林Pinusmassonianaplantation; CL:杉木人工林Cunninghamialanceolataplantation; RLD:根长密度 Root length density;RBD:根生物量密度 Root biomass density
2.2 根系直径频率分布
由图1可以看出不论是天然林、次生林,还是人工林;不论是多乔木树种林分(米槠天然林、米槠天然更新林、米槠人促更新林),还是单一乔木树种林分(米槠人工林、杉木人工林、马尾松人工林),林分水平的根系直径频率分布均呈明显的单峰型分布。0—5 mm直径细根的根长分布主要集中于0—2 mm直径范围内,6种林分细根在2 mm直径处的累积频率除米槠天然林接近95%外,其它5片林分均达到95%以上。由此可以看出,≤2 mm细根的累积根长在0—5 mm直径范围内占绝对优势。而这其中又以0—1 mm直径范围的累积频率最大,米槠天然林、米槠天然更新林、米槠人促更新林、米槠人工林、杉木人工林、马尾松人工林0—1 mm直径范围累积频率均在70%以上,分别为74.7%、77.9%、81.3%、75.9%、74.7%和78.9%。
米槠天然林、米槠人工林以及杉木人工林三者根长的直径分布相似,均在直径0.7 mm处根长比重最大;而米槠人促更新林、米槠天然更新林和马尾松人工林三者的根系直径频率分布亦较相似,均在0.3 mm直径处的根长比重达到最大(图1)。
2.3 累积频率分布函数的拟合
采用对数正态分布累积频率分布函数(CDF log-normal)对6种林分在0—5 mm直径范围内直径频率分布的累积频率进行回归分析后发现(表2),决定系数R2均达到0.99以上,说明该函数可以很好地拟合本研究6种不同更新方式林分根长的直径分布。
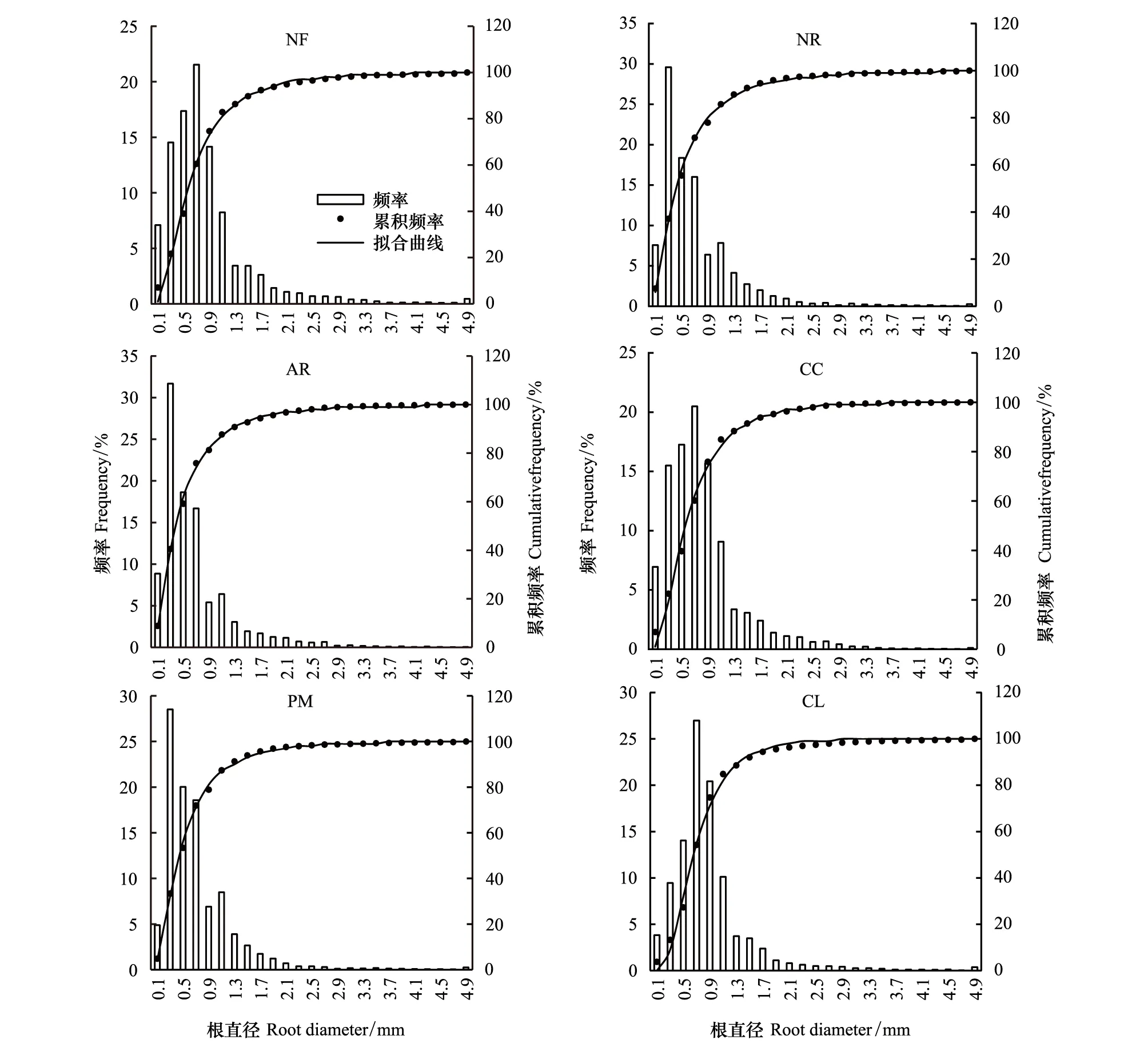
图1 6种林分根系的直径频率分布、累积频率及拟合曲线Fig.1 The root diameter frequency, the cumulative frequency and the fitted curve in the six standsNF:米槠天然林Nature forest of Castanopsis carlesii;NR:米槠天然更新林Secondary forest of Castanopsis carlesii through natural regeneration;AR:米槠人促更新林Secondary forest of Castanopsis carlesii through natural regeneration with anthropogenic promotion;CC:米槠人工林Castanopsis carlesii plantation;PM:马尾松人工林Pinus massoniana plantation; CL:杉木人工林 Cunninghamia lanceolata plantation
由图2可以看出,6种林分的μ值和σ值呈良好的线性负相关,R2=0.8787,P=0.006。其中,μ值小、σ值大时,反映在对数正态分布累积频率分布函数中的情况就是根系的根长分布更多的集中在径级较小细根范围,且比重较大。随着μ值增大、σ值减小,根系的分布不在过于集中于径级较小的细根部分,这表明这两个参数呈现出很好的协同性变化,代表着根系资源获取策略可能存在着一个权衡轴,即μ值和σ值的协同变化反映着根系资源获取中广布与精细之间的权衡变化。同时,从该权衡轴上亦可以很好地看出不同林分细根形态和分支结构特征及养分获取策略的差异,即米槠人促更新林、米槠天然更新林和马尾松人工林较相似,米槠天然林、米槠人工林和杉木人工林较类似。

表3 6种林分的μ值、σ值及R2 值Table 3 The value of μ, σ and R2 for the six stands
NF:米槠天然林 Nature forest ofCastanopsiscarlesii; NR:米槠天然更新林Secondary forest ofCastanopsiscarlesiithrough natural regeneration; AR:米槠人促更新林Secondary forest ofCastanopsiscarlesiithrough natural regeneration with anthropogenic promotion; CC:米槠人工林Castanopsiscarlesiiplantation; PM:马尾松人工林Pinusmassonianaplantation; CL:杉木人工林Cunninghamialanceolataplantation

图2 6种林分μ值和σ值线性拟合图Fig.2 The linear fit for the value of μ and σ of the six stands
3 讨论
3.1 林分尺度的根系直径频率分布
在植物个体水平上,一些研究发现植物细根直径分布频率呈单峰型分布,且符合对数正态分布[16]。本研究发现,在林分尺度上细根的直径频率分布亦主要呈单峰型分布,并亦能用对数正态分布很好地拟合。虽然少数研究认为细根的频率直径分布有时可以呈双峰型分布[25],但在林分尺度上这种双峰型分布并不明显。这些结果为从林分尺度上描述细根直径频率分布提供了依据。由于根系的直径频率分布整合了根系结构与功能很多方面的信息,如根长、生物量的直径分布、根形态特征和分支特点等,加上根系直径分布测定相对简单易行,因而研究林分尺度的根系直径频率分布,对于细根周转、碳吸存和养分通量的估算,揭示森林地下竞争状况和对土壤资源有效性的响应,以及生态系统模型模拟上均具有十分重要的作用。
对数正态分布累积分布函数的两个参数(μ值和σ值)可以很好地指示根系的分支结构和养分利用特点,亦可反映根系的拓扑结构[24]。如低的μ值和高的σ值,反映了根系具有高的拓扑指数,表明具有更类似于鲱鱼骨形的分支结构;而大的μ值和低的σ值,表明根系具有低的拓扑指数,分支结构更随机或更像二叉分支结构。根的分支结构可以影响其养分获取的效率[26],如Bouma 等[27]的研究表明,鲱鱼骨形分支结构的养分吸收效率更高,即单位碳投入所获取的养分更高,这在养分较高的土壤中可带来竞争上的优势。然而,二叉分支结构可获得更大的土壤资源空间利用潜力,在土壤养分缺乏的生境可以带来竞争上的优势。
本研究发现,对数正态分布累积分布函数的μ值和σ值呈现很好的负相关,呈现出协同性变化,表明这两个参数可能反映了根系资源利用策略的一个权衡轴。这可能与一些研究者提出的根系资源利用中存在权衡的理论相吻合。如Campbell 等[28]通过研究8种植物根系的觅食行为后发现根系觅食行为中可能存在着对资源利用广度和对资源利用精度之间的一个权衡,即有些植物能够在富养斑块中分配相对多的细根生物量,具有较高的觅食精确性;而有些植物则表现出尽可能多占有土壤空间,而在富养斑块中并没有多分配细根生物量,觅食精确性较低。相似的,Berntson[29]亦提出了一个类似的权衡假设,即资源利用效率和资源利用潜力之间的权衡。前者指单根体积所能利用的土壤体积,后者指根系所能全部利用的土壤体积。因而,本研究表明,考察μ值和σ值的变化,可以较直观地反映林分水平根系资源利用策略的变化。
3.2 不同更新方式根系直径频率分布的差异
本研究表明,利用根系直径频率分布拟合出的μ值和σ值,可以很好地反映不同更新方式林分根系资源利用策略的差异。
本研究中从米槠人工林到米槠次生林(米槠人促更新林和米槠天然更新林),树种多样性增加。从图2可见,随着这个树种多样性增加,μ-σ权衡轴上从右下端向左上端移动,反映了林分小径级根的比重增大、径级间的分布变异增加。这与伴随树种多样性增加,对地下资源的种间竞争更为强烈有关。另外,从表1可见,随树种多样性增加,根长密度和生物量密度亦相应增加,表明树种多样性增加引起了地下“超产”现象[30],这除了与生态位互补有关外,还可能与对地下资源的种间竞争加强而引起的对地下投入增加有关。
从图2亦可见,米槠人工林、马尾松人工林和杉木人工林在μ-σ权衡轴上处于明显不同的位置,表明这3种人工林具有不同的地下资源利用策略。这可能与人工林乔木层树种特性差异引起。马尾松人工林的μ值最小,这可能与其为强阳性先锋树种,比较适应干旱环境条件有关。在干旱的条件下,较小的细根直径(和导管直径)有利于保障树木水力结构传输的安全(即避免导管出现空穴现象)[31],虽然以牺牲水分运输效率为代价[32]。而米槠为演替后期树种,杉木原为常绿阔叶林林内伴生树种,林内环境相对比较阴湿,水分条件较高且较稳定,因而树木优先选择提高水分运输效率(即增大细根及导管直径)。
比较有意思的是,米槠天然林在μ-σ权衡轴上处于与米槠人工林相近的位置,而与米槠天然更新林和米槠人促更新林差异很大。天然林中虽然乔木树种丰富,但林木分化非常严重,导致乔木层中米槠等少数树种占绝对优势,而林下植被较少,从而种间的竞争可能比较缓和,导致根系对土壤资源利用策略类似于米槠人工林。
[1] 张小全, 吴可红. 森林细根生产和周转研究. 林业科学, 2001, 37(3):126- 138.
[2] Vogt K A, Vogt D J, Palmiotto P A, Boon P, O′Hare J, Asbjornsen H. Review of root dynamics in forest ecosystems grouped by climate, climatic forest type and species. Plant and Soil, 1995, 187(2):159- 219.
[3] 王向荣, 王政权, 韩有志, 谷加存, 郭大立, 梅莉. 水曲柳和落叶松不同根序之间细根直径的变异研究. 植物生态学报, 2005, 29(6):871- 877.
[4] Williamson M H, Fitter A. The characters of successful invaders. Biological Conservation, 1996, 78(1- 2):163- 170.
[5] Pregitzer K S, Laskowski M J, Burton A J, Lessard V C, Zak D R. Variation in sugar maple root respiration with root diameter and soil depth. Tree Physiology, 1998, 18 (10):665- 670.
[6] 常文静, 郭大立. 中国温带、亚热带和热带森林45个常见树种细根直径变异. 植物生态学报, 2008, 32(6):1248- 1257.
[7] Zobel R W, Kinraide T B, Baligar V C. Fine root diameters can change in response to changes in nutrient concentrations. Plant and Soil, 2007, 297(1- 2):243- 254.
[8] 黄石竹, 张彦东, 王政权. 树木细根养分内循环. 生态学杂志, 2006, 25(11):1395- 1339.
[9] Pregitzer K S. Fine roots of trees—a new perspective. New Phytologist, 2002, 154(2):267- 270.
[10] Eissenstat D M, Wells C E, Yanal R D, Whitbeck J L. Building roots in a changing environment:implications for root longevity. New Phytologist, 2000, 147(1):33- 42.
[11] 李俊英, 王孟本, 史建伟. 应用微根管法测定细根指标方法评述. 生态学杂志, 2007, 26(11):1842- 1848.
[12] 史建伟, 于水强, 于立忠, 韩有志, 王政权, 郭大立. 微根管在细根研究中的应用. 应用生态学报, 2006, 17(4):715- 719.
[13] Hansson A C, Zhao A F, André O. Fine-root production and mortality in degraded vegetation in Horqin sandy rangeland in Inner Mongolia, China. Arid soil research and rehabilitation, 1995, 9(1):1- 13.
[14] 张小全, 吴可红, Murach D. 树木细根生产与周转研究方法评述. 生态学报, 2000, 20(5):875- 883.
[15] 袁一丁. 亚热带杉木人工林细根寿命研究[D]. 福建:福建师范大学, 2009.
[16] Scanlan C A, Hinz C. Using radius frequency distribution functions as a metric for quantifying root systems. Plant and Soil, 2010, 332(1- 2):475- 493.
[17] 曲秋玲, 王国梁, 刘国彬, 刘涛, 曹艳春, 袁子成. 施氮对白羊草细根形态和生长的影响. 水土保持通报, 2012, 32(2):74- 79.
[18] 马雪红, 周志春, 张一, 金国庆. 杉木不同家系对异质养分环境的适应性反应差异. 植物生态学报, 2008, 32(1):189- 196.
[19] 王超. 常绿阔叶林不同更新方式下土壤呼吸的动态及机制[D]. 福建:福建师范大学, 2009.
[20] Yang Y S, Chen G S, Lin P, Xie J S, Guo J F. Fine root distribution, seasonal pattern and production in four plantations compared with a natural forest in Subtropical China. Annals of Forest Science, 2004, 61(7):617- 627.
[21] 杨玉盛, 陈光水, 林鹏, 黄荣珍, 陈银秀, 何宗明. 格氏栲天然林与人工林细根生物量、季节动态及净生产力. 生态学报, 2003, 23(9):1719- 1730.
[22] 杨玉盛, 陈光水, 何宗明, 陈银秀, 谢锦升. 杉木观光木混交林群落细根净生产力及周转. 林业科学, 2001, 37(S1):35- 41.
[23] Pregitzer K S, DeForest J L, Burton A J, Allen M F, Ruess R W, Hendrick R L. Fine root architecture of nine North American trees. Ecological Monographs, 2002, 72(2):293- 309.
[24] Guo D L, Li H, Mitchell R J, Han W X, Hendricks J J, Fahey T J, Hendrick R L. Fine root heterogeneity by root branch order:exploring the discrepancy in root longevity and turnover estimates between minirhizotron and carbon isotope methods. New Phytologist, 2008, 177(2):443- 456.
[25] Anderson T M, Starmer W T, Thorne M. Bimodal root diameter distributions in Serengeti grasses exhibit plasticity in response to defoliation and soil texture:implications for nitrogen uptake. Functional Ecology, 2007, 21(1):50- 60.
[26] Fitter A H, Stickland T R. Architectural analysis of plant root systems. 2. Influence of nutrient supply on architectur in contrasting plant species. New Phytologist, 1991, 118(3):383- 389.
[27] Bouma T J, Nielsen K L, Van Hal J, Koutstaal B. Root system topology and diameter distribution of species from habitats differing in inundation frequency. Functional Ecology, 2001, 15(3):360- 369.
[28] Campbell, B D, Grime J P, Mackey J M L. A trade- off between scale and precision in resource foraging. Oecologia, 1991, 87(4):532- 538.
[29] Berntson G M. Modelling root architecture:are there tradeoffs between efficiency and potential of resource acquisition?. New Phytologist, 1994, 127(3):483- 493.
[30] 刘聪, 项文化, 田大伦, 方晰, 彭长辉. 中亚热带森林植物多样性增加导致细根生物量“超产”. 植物生态学报, 2011, 35(5):539- 550.
[31] Hacke U G, Sperry J S, Pittermann J. Drought experience and cavitation resistance in six shrubs from the Great Basin, Utah. Basic and Applied Ecology, 2000, 1(1):31- 41.
[32] 申卫军. 木本植物木质部空穴和栓塞化研究(综述). 热带亚热带植物学报, 1999, 7(3):257- 266.
Diameter frequency distribution for 0—5mm roots in six mid-subtropical forests subject to different regeneration approaches
HU Shuangcheng1,2, XIONG Decheng1,2,*, HUANG Jinxue1,2, WANG Weiwei1,2, HUANG Chaochao1,2, DENG Fei1,2, CHEN Yunyu1,2, CHEN Guangshui1,2
1StateKeyLaboratoryofHumidSubtropicalMountainEcology,Fuzhou350007,China2InstituteofGeographicalSciences,FujianNormalUniversity,Fuzhou350007,China
Diameter is an important structural trait for root systems, with roots of different thickness havings different physiological functions. Root length is a significant index of root function, while root diameter frequency distribution provides useful information. Thus, studies on the characteristics of root diameter frequency distributionenhance our understanding about the root exploitation strategy for soil resources in different plant or forest systems, as well as the interaction between roots and the soil environment, allowing us to model projections. Using the soil coring method, the diameter frequency distribution characteristics of roots (≤5 mm in diameter) in six forests was determined using different regeneration approaches. The forests included:the nature forest ofCastanopsiscarlesii(NF), the secondary forest ofCastanopsiscarlesiithrough natural regeneration (NR), the secondary forest ofCastanopsiscarlesiithrough natural regeneration with anthropogenic promotion (AR), theCastanopsiscarlesiiplantation (CC), thePinusmassonianaplantation (PM), and theCunninghamialanceolataplantation (CL), in Sanming, Fujian Province. Because it is different to differentiate between the roots of different species, the study was conducted at the community level. There were three main conclusions in this study. First, root length density (RLD) and root biomass density (RBD) for all the three diameter ranges (≤1 mm, ≤2 mm, and ≤5 mm) decreased from NF, NR, AR, to CC. Out of the three plantations, the highest RLD and RBD wereobtained in CC, while the lowest RLD and RBD was obtained in PM. Second, out of roots with 0—5mm diameter, 87%—98%and 65%—88%were represented by 0—2 mm and 0—1 mm diameter roots, respectively. The roots of NF, CC, and CL had the similar root diameter frequency distributions, with a maximum frequency of 0.7 mm diameter. The root diameter frequency distribution of AR, NR, and PM was also similar, with a maximum frequency of 0.3mm diameter. The root diameter frequency distribution for all six stands was unimodal, and fitted well with the cumulative lognormal distribution function, with the determination coefficientR2being above 0.99 for all stands. Third, the fitting parameters ofμandσin the cumulative lognormal distribution function represented different foraging strategies for soil resources in different stands.μandσwere strongly negatively correlated, which showed a trade-off in the fine root resource acquisition strategy. With the increasing tree species diversity,μtends to be smaller, whileσtends to be larger, reflecting the growing competition for nutrition and water among the roots of different trees species.μandσwere distinctly different among different plantation tree species, showing differences in the exploitation strategy for soil resources among plantation tree species. In conclusion, the cumulative lognormal distribution function well reflects stand level root diameter frequency distribution, in addition to root exploitation strategies for soil resources.
forest; fine root; root length; diameter frequency distribution; exploitation strategy for soil resources; log-normal distribution
国家重大科学研究计划课题(2014CB954003);国家自然科学基金重点项目(31130013);福建省杰出青年科研基金项目(2010J06009)
2014- 05- 05; < class="emphasis_bold">网络出版日期:
日期:2015- 05- 18
10.5846/stxb201405050888
*通讯作者Corresponding author.E-mail:xdc104@163.com
胡双成,熊德成,黄锦学,王韦韦,黄超超,邓飞,陈云玉,陈光水.中亚热带6种不同更新方式森林0—5mm根系直径频率分布.生态学报,2015,35(23):7812- 7820.
Hu S C, Xiong D C, Huang J X, Wang W W, Huang C C, Deng F, Chen Y Y, Chen G S. Diameter frequency distribution for 0—5mm roots in six mid-subtropical forests subject to different regeneration approaches.Acta Ecologica Sinica,2015,35(23):7812- 7820.
