天山林区六种灌木生物量的建模及其器官分配的适应性
2015-02-11常顺利张毓涛王文栋王慧杰
仇 瑶, 常顺利,*,张毓涛,王文栋,何 平,王慧杰,谢 锦
1 新疆大学资源与环境科学学院绿洲生态教育部重点实验室,乌鲁木齐 830046 2 新疆林科院森林生态研究所,乌鲁木齐 830063
天山林区六种灌木生物量的建模及其器官分配的适应性
仇 瑶1, 常顺利1,*,张毓涛2,王文栋2,何 平1,王慧杰1,谢 锦1
1 新疆大学资源与环境科学学院绿洲生态教育部重点实验室,乌鲁木齐 830046 2 新疆林科院森林生态研究所,乌鲁木齐 830063
灌木全株生物量估算模型的构建仍存在一定困难,对灌木生物量在器官分配上所体现的适应性研究也不够充分。以天山林区6种常见灌木为研究对象,在天山的东段、中段、西段林区分别设置样地进行群落调查,由此以全株收获法取得6种常见灌木若干标准株的全株、根、枝、叶及各径级根的生物量,将D2H(地径平方与高度的乘积)与V(冠幅面积与高度的乘积)分别选为估测模型的自变量,通过回归分析法建立了各种灌木全株生物量的最优估算模型,然后比较了此6种灌木全株生物量在营养器官上分配差异以及根系生物量在径级上的分配差异。结果表明:(1)天山林区6种常见灌木中,小檗(BerberisheteropodaSchrenk)、忍冬(LonicerahispidaPall. ex Roem. et Schuet.)、栒子(CotoneastermelanocarpusLodd.)的全株生物量约为8.48—9.01 kg,蔷薇(RosaspinosissimaL.)、绣线菊(SpiraeahypericifoliaL.)、方枝柏(JuniperuspseudosabinaFisch. et Mey.)的全株生物量约为2.71—3.20kg;(2)蔷薇、绣线菊、栒子的全株生物量最优估测模型是以V为自变量的函数,小檗、忍冬、方枝柏的全株生物量最优估测模型是以D2H为自变量的函数,各模型R2值均在0.850以上,且在P<0.05水平上达到显著,模型模拟结果达到了较高的准确度;(3)6种灌木全株生物量在根、枝上的分配比重差异不显著,仅在叶上的分配比重有差异(P<0.05);根系生物量在径级上的分配均呈现随根系径级下降而减少的规律,6种灌木在径级大于2 mm根上的分配比重存在差异(P<0.05,径级大于20mm根为P<0.01水平);(4)6种灌木全株生物量在营养器官上的分配差异以及根系生物量在径级上的分配差异均体现了各物种对其生境选择的适应策略。
生物量建模;根系分级;适应策略;灌木;天山
灌木层是森林中的重要片层,灌木生物量是群落的重要数量特征之一[1-3],对灌木生物量的关注有助于群落的物质循环、消耗、分配、转化、积累等研究工作的深入开展[4-5]。在林业传统上,对乔木生物量和蓄积量的研究较为普遍,但对灌木生物量的关注较为欠缺。目前,测定灌木生物量的方法仍然主要是全株收获法[6-7],但该法费时耗力且取样数量又很难满足统计学大样本的要求,存在较大的不确定性。因此,采用易测因子来估算灌木生物量更为可行,成为当前学界较为主流的工作思路。
早期进行的灌木生物量研究方法主要有样方法、平均木法、相对生长法等,随着人们对各种灌木生物量研究的深入,多数研究人员倾向于使用回归模型和数量化的方法[8-10]。以易测因子作为自变量,以生物量为因变量,通过数理统计的回归分析方法,建立起这些因子之间的相对生长方程,以此来推算、预测整体的生物量[4]。对灌木生物量进行建模能够在非破坏性、减小工作量基础上提供较为简单且准确的估算方法[11],使得对林区灌木生物量的连续清查成为可能。
植株生物量在各器官中的分配策略是植物生活史理论研究的主要内容[12-13]。植物在生长发育的过程中,通过不断优化自身的资源分配来提高其适应环境变化的能力[14],故生物量在根、枝、叶之间的分配是植物对环境长期适应的结果[12,15],在很大程度上反映了植物的生态适应对策。当前,对生物量在植株各器官上分配的研究多集中在对不同径级根系的生物量及生理功能的比较上[16-20],对其分配策略的生态适应性研究尚不充分,对灌木生物量的器官分配及其反映出的生态适应性问题更是鲜有报道。
本研究以天山林区6种常见灌木为研究对象,为排除水热空间分布梯度因素的影响,在天山东部、中部、西部分别设置灌木样地进行群落调查,在群落调查基础上取该6种灌木的标准株若干,以全株收获法求算全株生物量和各器官生物量,以实测数据构建天山林区6种灌木的单株生物量估算模型,并深入分析该6种灌木单株生物量在营养器官上的分配规律,探讨各物种在全株生物量的营养器官分配方面以及根系生物量的径级分配方面所体现出的生态适应性,对于更深入了解天山林区灌木的生物学特性具有一定意义。
1 材料与方法
1.1 研究区概况
天山林区以天山云杉(PiceaschrenkianaFisch. et Mey.)纯林为主,林缘、林窗及林下的灌木主要有栒子(CotoneastermelanocarpusLodd.)、小檗(BerberisheteropodaSchrenk及B.nummulariaBge.)、蔷薇(RosaspinosissimaL.)、绣线菊(SpiraeahypericifoliaL.)、方枝柏(JuniperuspseudosabinaFisch. et Mey.)、锦鸡儿(CaraganaleucophloeaPojark.)、忍冬(LonicerahispidaPall. ex Roem. et Schuet.)等,林下草本植物主要有老鹳草(GeraniumrotundifoliumL.)、羽衣草(AlchemillatianschanicaJuz.)、羊角芹(AegopodiumpodagrariaL.)等。林下土壤均为山地灰褐色森林土[21]。
根据云杉森林长势、地形地貌、气候等条件的不同,把整个天山北坡从东到西分为东天山,中天山和西天山三段分别进行群落调查和采样。
天山东段以哈密林场为调查采样区,主要依托巴里坤县白石头景区(93°37′—93°41′E,N43°9′—43°13′)开展工作,该地属温带大陆性干旱半干旱气候,夏季炎热、冬季寒冷、四季分明、干燥少雨,年平均气温为4.7 ℃,1月份极端最低气温为-42.6 ℃,7月份极端最高气温43 ℃;年平均降水量为176 mm,蒸发量2141 mm,无霜期平均156 d,年日照时数2280—3230h。
天山中段以乌鲁木齐县板房沟林场为调查采样区,主要依托中国森林生态系统研究网络(CFERN)天山森林生态系统定位站研究站(87°07′—87°28′E,43°14′—43°26′N)开展工作,该地属温带大陆性气候,年总辐射量达5.85×105 J cm-2a-1),年均气温约为2—3 ℃,历年极端最高温为30.5 ℃,极端最低温为-30.2 ℃;年降水量400—600mm,最大积雪深度为65 cm。
天山西段以昭苏林场为调查采样区,主要依托昭苏县阿克苏乡(80°48′—80°57′E,42°41—42°44′N)开展工作,该地属温带大陆性气候,年平均温度5—7 ℃,1月平均气温为-10℃,7月平均气温为22.6 ℃;年平均降水量为600—800mm,全年60%的降水量集中在5—8月,年平均蒸发量1100—1200mm,年平均相对湿度70%以上,无霜期120d。
从东至西,天山林区呈现出较为明显的水分增多趋势和较不明显的热量减少趋势。
1.2 群落调查与全株生物量收获
2013年7—8月,在天山东段哈密林场、中段板房沟林场、西段昭苏林场分别设置栒子、小檗、蔷薇、绣线菊、方枝柏、忍冬等6种代表性灌木的样地,每种灌木在每个调查采样区至少做1个10m×10m的调查样方。在样方调查数据的基础上,以地径(D, cm)、株高(H, m)、冠幅(CA, m2)为指标,分别对应选取6种灌木的标准株。每个物种在3个调查采样区各全株采样1—2个标准株,共5—6株。样方设置与标准株采样情况见表1。
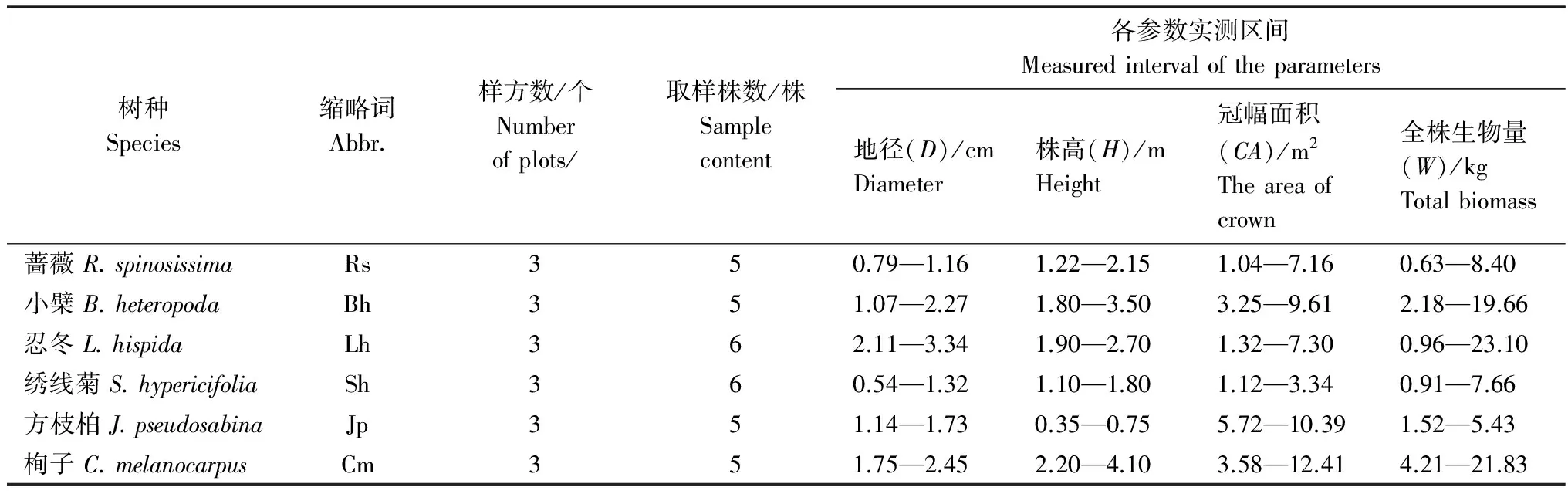
表1 样方调查与标准株取样统计表Table 1 The results of sample plots survey
用全收获法将各标准株整株挖出,深度为根系分布所达范围。全株分为根、枝、叶3个营养器官,其中又按径级将根分为根桩(>20mm)、粗根(10—20mm)、中根(5—10mm)、小根(2—5 mm)和细根(<2 mm)等5个层级。分别称取每个采样株各营养器官以及各径级根的鲜质量,再按比例称取各组份样品带回实验室,于105 ℃条件下烘干至恒重称重。通过各采样株各器官的鲜重、干重、含水率,计算各采样株灌木全株生物量以及各器官的干物质量。
1.3 生物量建模方法
根据前人的研究,本研究先选取变量地径(D,cm)、株高(H,cm)、冠幅面积(CA=π×C1×Cw/4,m2)、地径平方与高度的乘积(D2H,cm3)、植株体积(V=CA×H,m3)、全株生物量(W,kg)等6个因素进行相关分析,筛选出与全株生物量相关性较好的状态因子,再以相关性较好的几个状态因子(地径(D)、高度(H)、冠幅面积(CA=π×C1×Cw/4)、植株体积(V=CA×H)、D2H)为自变量,全株生物量(W)为因变量进行回归分析,最后选用以下几种生物量研究中常用的回归模型(见式1至式7)建立预测林下灌木生物量的回归模型:
y=b0+b1x
(1)
y=b0+b1x+b2x2
(2)
y=b0(b1)x
(3)
y=b0+b1lnx
(4)
y=b0+b1x+b2x2+b3x3
(5)
y=b0e(b1x)
(6)
y=b0xb1
(7)
通过判定系数(R2)和标准误(SEE)的大小及回归检验显著水平来评价每个物种的各方程的优劣,从中选出拟合度最好、相关最密切的数学模型作为该种灌木全株生物量的估算模型。
1.4 数据分析方法
本研究中的数据采用Excel进行数据统计,并用SPSS17.0统计软件进行相关分析、回归分析和多重比较(LSD)。
2 结果与分析
2.1 6种灌木的生物量模型构建
2.1.1 生物量回归方程自变量的选择
对样地内选取的各灌木全株生物量(W)与其地径(D)、高度(H)、冠幅面积(CA)等因子之间进行相关分析(表2)。
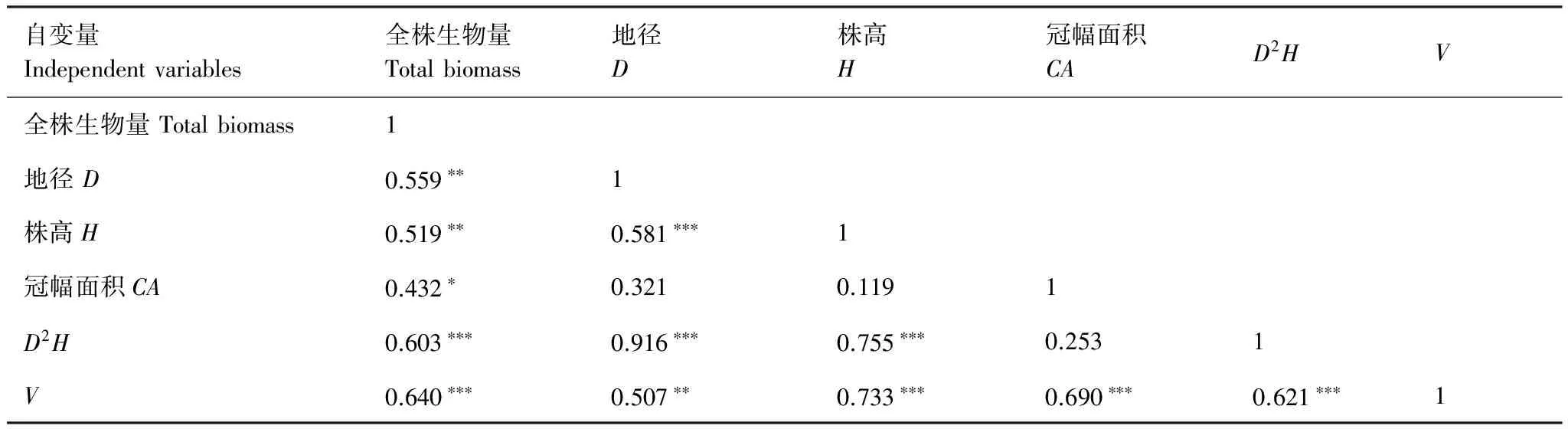
表2 6种灌木重要变量间的相关分析Table 2 The correlation analysis among independent variables for six shrubs
D:地径 Diameter;H:株高 Height;CA:冠幅面积The area of crown(CA=π×C1×Cw/4)C1:crown major axis;Cw:crown minor axis;D2H:地径平方与高度的乘积 Square of diameter(D2) multiplied by height(H);V:植株体积 The volume of shrub(V=CA×H); *P<0.05;**P<0.01;***P<0.001
表2可知,各个变量与全株生物量之间都有较强的相关关系,其中,包含地径平方与高度的乘积(D2H)、植株体积(V=CA×H)与全株生物量的相关系数最大,相关性在P<0.001水平上达到显著。因此,建立天山林区6种灌木生物量回归模型时,分别选用D2H、V作为自变量。
2.1.2 生物量模型的确立
为了提高建立的回归方程的准确度,分别用V和D2H两个自变量与几个常用的生物量回归方程进行拟合,通过筛选判定系数(R2)较高、标准误(SEE)较小以及通过回归检验的方程,找到拟合度最好的方程,结果见表3。
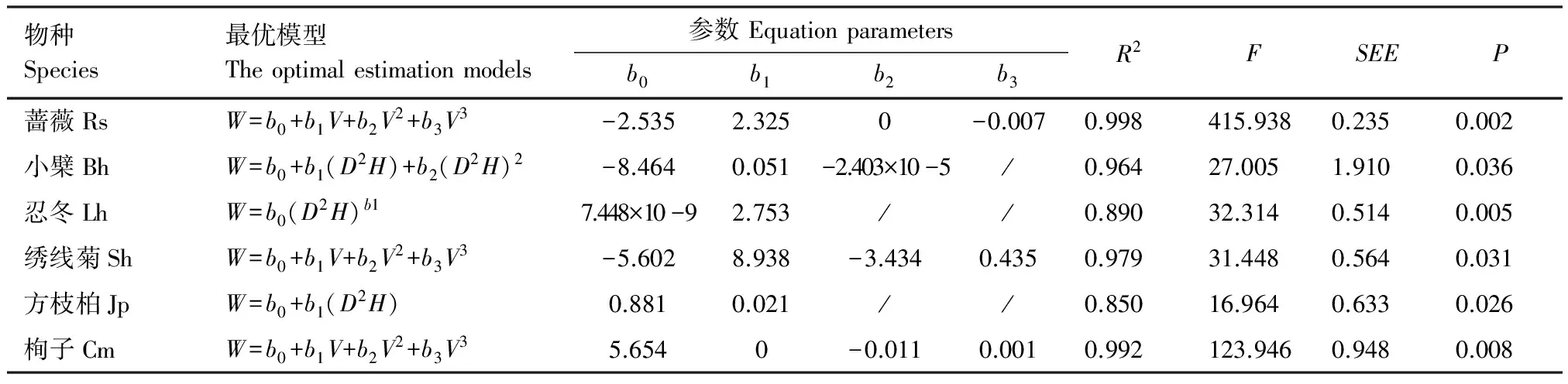
表3 6种灌木全株生物量的最优估算模型Table 3 The optimal estimation models for total biomass of six shrub species
蔷薇Rs:R.spinosissima;小檗Bh:B.heteropoda;忍冬Lh:L.hispida;绣线菊Sh:S.hypericifolia;方枝柏Jp:J.pseudosabina;栒子Cm:C.melanocarpus;R2为判别系数;F为方差分析的F值the value of ANOVA;SEE为估计值的标准误差the standard error of estimate;P为显著性水平
蔷薇、绣线菊和栒子的全株生物量与V相关性最好,最优模型为y=b0+b1V+b2V2+b3V3,R2分别为0.998、0.979 和 0.992。小檗、忍冬和方枝柏与D2H相关性最好,小檗的最优模型为y=b0+b1(D2H)+b2(D2H)2,R2为0.964。忍冬的最优模型为y=b0(D2H)b1,R2为0.890。方枝柏的最优模型为y=b0+b1(D2H),R2为0.850。蔷薇、忍冬和栒子在P<0.01水平上达到显著,小檗、绣线菊和方枝柏在P<0.05水平上达到显著。天山林区6个灌木物种的全株生物量估算模型结果见式8—式13。
蔷薇Rs:
W=-2.535+2.325V-0.007V3
(8)
绣线菊Sh:
W=-5.602+8.938V-3.434V2+0.435V3
(9)
栒子Cm:
W=5.654-0.011V2+0.001V3
(10)
小檗Bh:
W=-8.464+0.051(D2H)-(2.403×10-5)(D2H)2
(11)
忍冬Lh:
W=(7.448-9)(D2H)2.753
(12)
方枝柏Jp:
W=0.881+0.021(D2H)
(13)
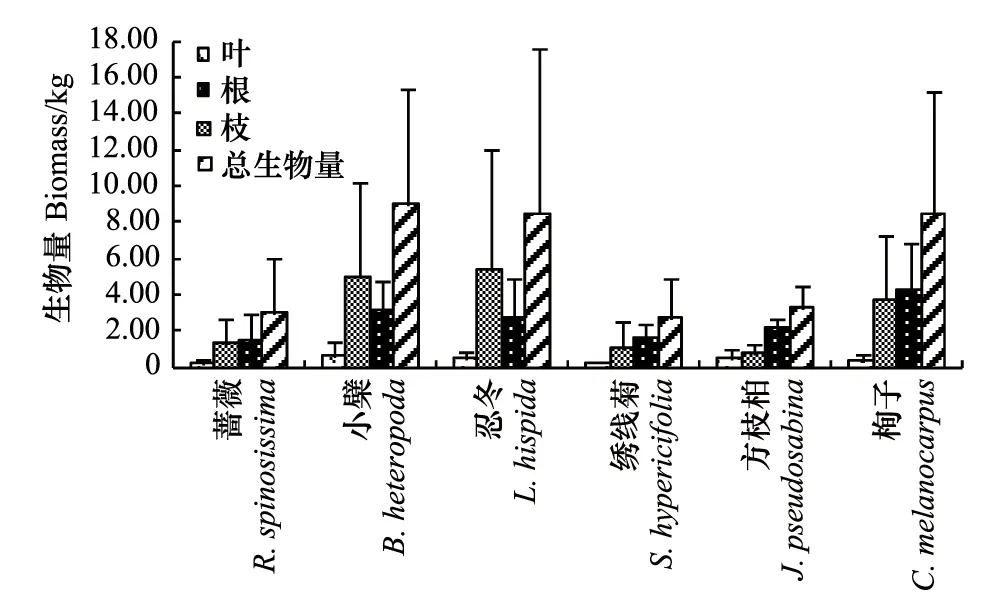
图1 6种灌木全株及营养器官的生物量 Fig.1 The biomass of leaf, branch, root and total biomass of six shrubs
2.2 6种灌木全株生物量的基本情况
对6种灌木标准木的全株生物量和根、枝、叶生物量分别在多个重复中求均值进行比较分析,结果见图1。图1表明,6种灌木中,小檗、忍冬、栒子的全株生物量较大,在8.48—9.01 kg之间;蔷薇、绣线菊、方枝柏全株生物量较小,在2.71—3.20kg之间。6种灌木的根生物量在1.38—4.20kg之间;枝生物量在0.70—5.37 kg之间,其中小檗、忍冬的略大为5.00kg和5.37 kg,蔷薇、绣线菊、方枝柏、栒子略小,在0.70—3.73 kg之间;叶生物量均在0.70kg以内。
2.3 6种灌木全株生物量在营养器官上的分配
对6种灌木生物量在营养器官根、枝、叶间的分配进行比较分析,进行物种间的差异比较,得到图2。图2表明,6种灌木在营养器官根、枝、叶间的生物量分配呈现出大体相同的趋势,但同一器官在不同灌木全株生物量中所占比例不同。
图2可见,各营养器官生物量分配总体上表现为根生物量>枝生物量>叶生物量。6种灌木根、枝、叶生物量占全株生物量的比例各不相同,根生物量所占比例在47.8%—72.9%之间,物种间差异不显著;枝条生物量所占比例在20.6%—46.5%之间,物种间差异也不显著;而叶生物量所占比例在2.4%—10.1%之间,其中方枝柏为10.1%、栒子为3.5%、绣线菊为2.4%,仅呈现出方枝柏与绣线菊、栒子有显著性差异(P<0.05),而其他物种间差异不显著。即,6种灌木间生物量在根、枝上的分配差异不显著,仅在叶上的分配有差异。
2.4 6种灌木根系生物量在径级上的分配
对6种灌木生物量在根系不同径级间的分配进行比较分析,进行物种间的差异比较,得到图3。图3表明,6种灌木在根系不同径级间的生物量分配呈现出大体相同的趋势,但同一径级的根在不同灌木总根系中生物量所占比例不同。
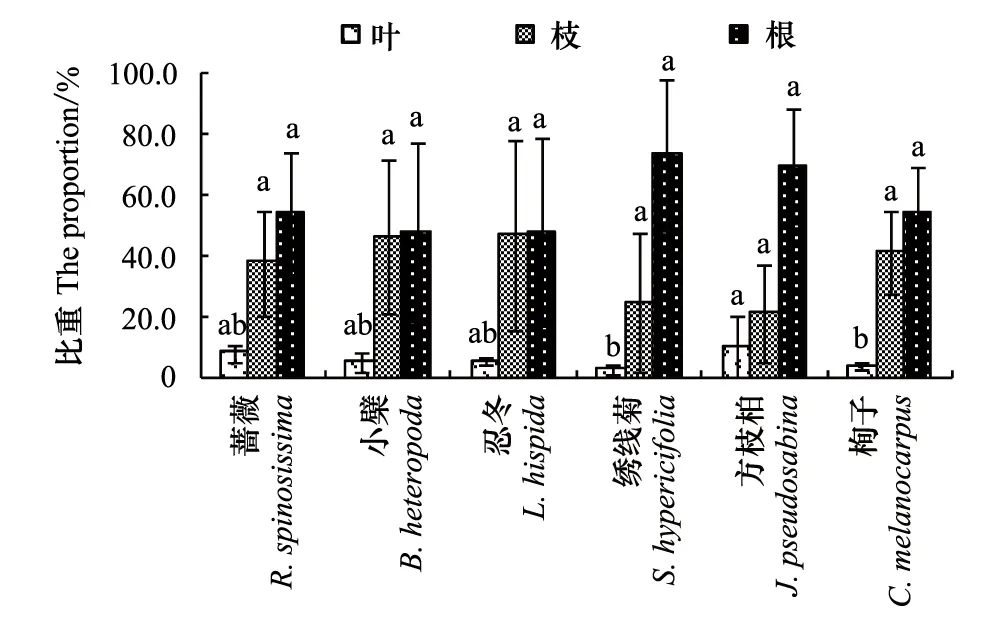
图2 6种灌木全株生物量在各营养器官中的分配Fig.2 The allocation of six shrubs` total biomass in vegetative organs
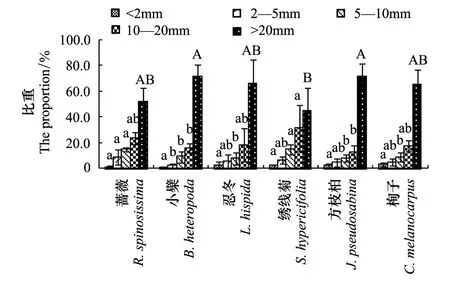
图3 6种灌木根系生物量在径级上的分配Fig.3 The allocation of six shrubs` underground biomass in organs小写字母a、b表示统计量的差异在 P<0.05水平上;大写字母A、B表示统计量的差异在P <0.01的水平上
图3可见,灌木根系各不同径级生物量分配大小均表现为根桩(>20mm)>粗根(10—20mm)>中根(5—10mm)>小根(2—5 mm)>细根(<2 mm)。通过方差分析和LSD多重比较可以得出,6种灌木间根系生物量在各径级根上的分配有所区别。
在根桩(>20mm)生物量的分配上,方枝柏最大为71.6%、绣线菊最小为44.9%,方枝柏、小檗与绣线菊有极显著性差异(P<0.01),而其他物种间均无显著性差异。
在粗根(10—20mm)生物量的分配上,绣线菊最大为31.2%、方枝柏最小为13.1%,绣线菊与小檗、方枝柏间差异显著(P<0.05),而其他物种间均无显著性差异。
在中根(5—10mm)生物量的分配上,蔷薇和绣线菊最大均为14.9%、忍冬最小为8.0%,蔷薇、绣线菊与其他4种灌木间均有显著差异(P<0.05)。
在小根(2—5 mm)生物量的分配上,蔷薇最大为8.9%、小檗最小为2.5%,蔷薇与小檗间有显著差异(P<0.05),其他物种间均无显著性差异。
在细根(<2 mm)生物量的分配上,6种灌木均不超过3.3%,且6种灌木间无显著性差异。
3 结论与讨论
3.1 关于灌木生物量模型构建的探讨
本研究分别在天山东段、中段和西段各自设立林下灌木样地,对天山云杉森林林区常见的6种林下灌木进行全株生物量的取样,利用回归分析的方法建立了这6种灌木全株生物量的最优估测模型。
在生物量估算建模中,回归模型自变量的选取是难点所在[1,5-6,22-26]。本研究通过对地径(D)、株高(H)、冠幅面积(CA)、D2H、V等易测因子与全株生物量进行相关分析发现,D2H、V与全株生物量相关性最大,均在P<0.001水平上达到显著,与前人同类的研究较为一致[6]。这表明利用二因子复合变量比单因子变量进行模型拟合的效果要好,故本文分别用D2H和V对6种灌木进行回归模型的拟合,从中筛选出每种灌木的最优模型。
从模型模拟的结果来看,每种灌木选取的自变量和最优方程并不完全相同,这是因为不同种灌木的形态是不同的,而生物量估测参数的选择与灌木形态关系密切相关[27],因此本文不能用同一种变量对6种不同灌木去构建形式上相对统一的方程。
有些灌木的生长形态类似于圆柱型或圆锥形,以D2H为自变量进行模型的模拟有较高的准确性。而有些较为矮小的多分枝丛生灌木,以横向发展冠幅来增加光合作用面积[28],故以V为自变量来估算生物量具有较高的精度[24]。本研究中灌木生物量最优模型形式多数为三次方程W=b0+b1x+b2x2+b3x3,这与前人同类工作较为一致[6]。另外,有些前人同类工作也采用W=b0(X)b1模型对生物量进行描述[22-23,29-32],本研究中只有忍冬的生物量最优模型为W=b0(D2H)b1。
虽然本研究限于条件对每种灌木的取样仍然偏少,但在实测数据的基础上,6种灌木模型模拟的R2值均在0.850以上,在P<0.05水平上达到显著。因此,仍然可以认为模拟结果达到了较高的准确度。
林下灌木的生长状况会因林分、土壤等立地条件存在一定的差异,可能会出现同物种在不同立地条件下估算模型不同的情况。虽然天山林区从东至西呈现出较为明显的水分增多和较不明显的热量减少趋势,且本研究在天山东、中、西段分别得到了6种灌木的全株生物量,各有5—6个重复,但若在东、中、西三段天山间进行6种灌木单株生物量及其各自估算模型的比较的话,则在每段研究区的取样重复数都是不足的(表1)。若开展此分析,工作量将继续成倍增加。因此,本文是将每种灌木在三段天山的取样均作为重复,分别构建了6种灌木在整个天山林区统一的生物量估测模型。在天山林区以外,应用本文的6种灌木生物量估算模型时,需进行进一步验证,当存在冠幅、植株高度变量等的外推时也需做进一步的验证。
3.2 天山林区灌木全株生物量在营养器官上分配策略的适应性分析
在植物的整个生长发育过程中,可利用的光合固定产物是有限的,因此植物必需对有限的资源进行分配,在获取光合固定产物的地上部分的器官和吸收土壤中水分和养分的地下部分的器官之间进行权衡[33]。
单株生物量在营养器官上分配格局的不同,反映了不同物种的对其生境的适应策略。最优分配理论认为,植物通过调节生物量在各器官中的分配来适应外界的环境条件,更多的生物量总是分配给那些获取最受限制资源的器官[12,34]。当光照资源成为最主要的限制因子时,植物将更多的生物量分配给地上器官(枝、叶)以尽可能多的获取光照;当水分或养分成为最主要的限制因子时,植物将更多的生物量分配给地下器官(根系)以尽可能多的获取土壤水分或养分[35]。
从总体来看,天山林区6种代表性灌木全株生物量均呈现出根>枝>叶的规律,与前人研究结果一致[36-37],但这6种灌木全株生物量在营养器官的分配上又可以分为三类较有差异的生境适应类型(图4)。
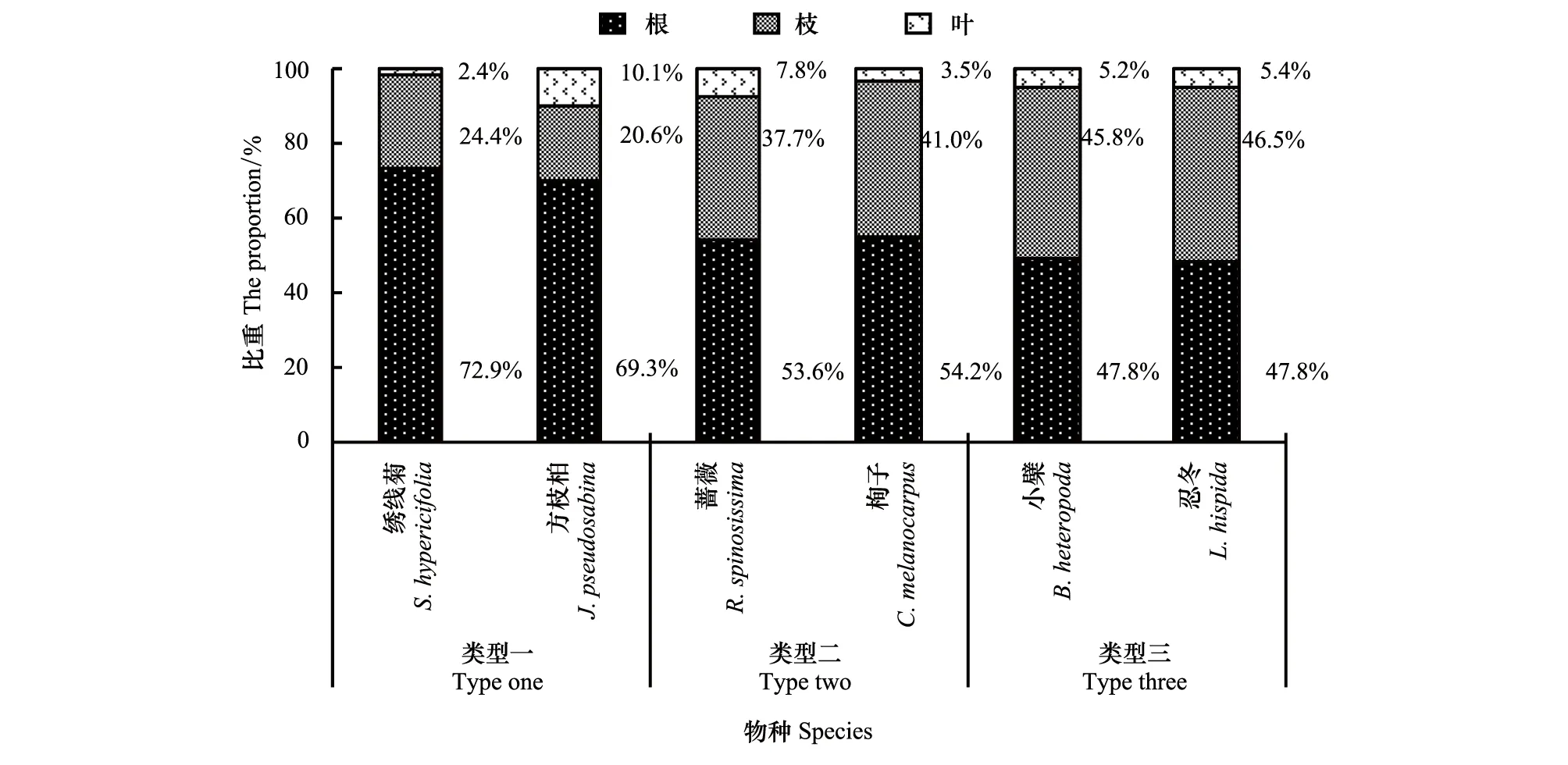
图4 6种灌木全株生物量在营养器官分配上的适应性Fig.4 Adaptability of the allocation of six shrubs` total biomass in vegetative organs
类型一包括绣线菊、方枝柏。该类型的两个物种主要分布在阳坡半阳坡、土层浅、砾石含量高的生境,它们的株形普遍矮小,全株生物量较低(图1),根占全株生物量的大部分(70%左右),枝占全株生物量的超过20%(图4)。此种类型表明,从土壤中获取水分和养分是这两个物种最严峻的生态压力,对根系生物量的大投入体现了它们对较干旱和贫瘠生境的适应。
类型二包括蔷薇、栒子。该类型的两个物种主要分布在半阳坡、阴坡的林缘和林窗,它们株形较为高大,植株密度较高,全株生物量较高(图1),根占全株生物量的略超过50%,枝占全株生物量的40%左右,叶生物量占全株生物量比例波动较大(图4)。此种类型表明,种内和种间对光照的竞争在一定程度上是这两个物种较主要的生存压力,对营养器官生物量的分配体现了它们对水分养分条件较好、光照相对充足生境的适应特点。
类型三包括小檗、忍冬。该类型的两个物种主要分布在林下、阴坡半阴坡的林缘或林窗,它们的株形相对较为纤细,全株生物量较高(图1),根占全株生物量的略低于50%,地上部分的枝、叶生物量占全株生物量的50%以上,且叶生物量占全株生物量比例较为稳定(图4)。此种类型表明,对光照的竞争是这两个物种最主要的生态压力,对营养器官生物量的分配体现了它们对光照条件稀缺生境的适应特点。
3.3 天山林区灌木根系生物量在径级上分配策略的适应性分析
植物根系起着固定植株、摄取运输和贮存水分与营养等重要作用[26]。不同径级的根的功能是有一定区别的[38-39],其中,粗根通常在根系生物量中占主要组分,是主要的贮藏器官并起着固定植株的作用,但在水分和养分吸收、根土相互作用等方面的功能较小[26];而细根通常在根系生物量中占比较小[40],但细根是植物根系获得水分和养分的最重要的途径[19]。因此,植物对特定生境的适应不仅通过调节根系生物量占全株生物量比例来体现,也往往通过各径级根系生物量占根系整体生物量比例来体现[41]。
从总体来看,天山林区6种代表性灌木均呈现出各径级根系生物量随径级减小而占比减小的规律,这符合乔灌木物种通常的根系构型。但这6种灌木的各径级根系生物量占比分配上又可以分为两类较有差异的生境适应类型(图5)。
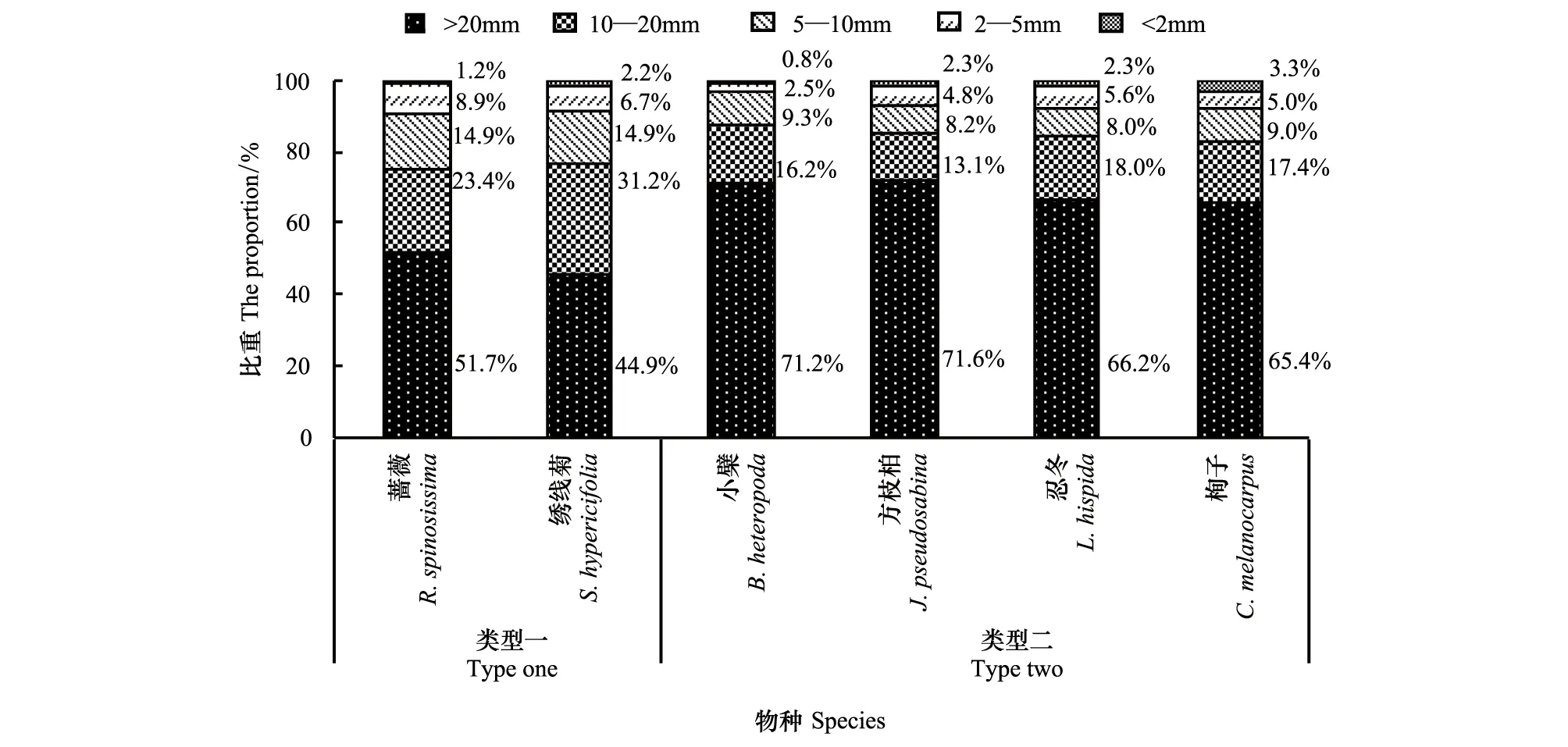
图5 6种灌木根系生物量在径级上分配的适应性Fig.5 Adaptability of the allocation of six shrubs` underground biomass in organs
类型一包括蔷薇、绣线菊。该类型的两个物种中,起固定植株作用的根桩(>20mm)占根系总生物量的50%左右,起延展根系空间作用的粗根(10—20mm)占有20%以上的比重,而在水分、养分的吸收方面发挥重要作用的中根(5—10mm)、小根(2—5 mm)、细根(<2 mm)占25%左右的较大比重(图5)。此种类型表明,这两个物种在各自的生境中要么存在较强的种内或种间的竞争压力(例如蔷薇),要么面对较为贫瘠干旱的土壤环境(例如绣线菊),因此需要大量更富有活性的径级在10mm以下的根系来满足植株旺盛生长对营养和水分的需求。因此,此类型属于在竞争生境(蔷薇)或胁迫生境(绣线菊)中适应的较强者。
类型二包括小檗、方枝柏、忍冬、栒子。该类型的4个物种中,起固定植株作用的根桩(>20mm)占根系生物量的70%左右,起延展根系空间作用的粗根(10—20mm)占有15%—20%的较稳定比重,而在水分、养分的吸收方面发挥重要作用的中根(5—10mm)、小根(2—5 mm)、细根(<2 mm)仅占15%左右的比重(图5)。此种类型表明,这4个物种在各自的生境中要么处在光照条件稀缺的生境中(例如小檗、忍冬),要么面对较为贫瘠干旱的群落演替早期的土壤环境(例如方枝柏),要么面对较强的种内或种间竞争压力的抑制(例如栒子),从而导致了它们的生长速率相对低下,其正常生长过程对营养和水分的需求相对较低,使得富有活性的径级小于10mm的根系在根系总生物量中的占比较小。因此,此类型属于适应胁迫生境(小檗、忍冬)、先锋生境(方枝柏)或竞争生境(栒子)中适应的较弱者。
3.4 结论
(1)将D2H与V选为天山林区6种常见灌木全株生物量估测模型的自变量,得到各物种的全株生物量估测模型,各模型R2值均在0.850以上,且在P<0.05水平上达到显著,表明模型模拟结果达到了较高的准确度。
(2)天山林区6种灌木的全株生物量在营养器官根、枝、叶上的分配均呈现出根>枝>叶的现象,且在根、枝上的分配比重差异不显著,仅在叶上的分配比重有差异(P<0.05)。根系生物量在不同径级上的分配均呈现为根桩(>20mm)>粗根(10—20mm)>中根(5—10mm)>小根(2—5 mm)>细根(<2 mm),6种灌木间仅在细根上的分配差异不显著(P<0.05),在其他径级根上的分配均有差异(P<0.05,其中根桩为P<0.01)。
(3)天山林区6种常见灌木全株生物量在营养器官上分配差异体现出各物种对生境差异的适应性,类型一为绣线菊、方枝柏,它们对较干旱和贫瘠生境较为适应;类型二为蔷薇、栒子,它们对水分养分条件较好、光照相对充足生境较为适应;类型三为小檗、忍冬,它们对光照条件稀缺的生境较为适应。
(4)天山林区6种常见灌木根系生物量在径级上的分配差异也体现出各物种对生境选择的适应策略,类型一为蔷薇、绣线菊,它们属于在竞争生境(蔷薇)或胁迫生境(绣线菊)中适应的较强者;类型二为小檗、方枝柏、忍冬、栒子,它们属于适应胁迫生境(小檗、忍冬)、先锋生境(方枝柏)或竞争生境(栒子)中适应的较弱者。
[1] 张峰, 上官铁梁, 李素珍. 关于灌木生物量建模方法的改进. 生态学杂志, 1993, 12(6):67- 69.
[2] 赵振勇, 王让会, 张慧芝, 王雷. 天山南麓山前平原柽柳灌丛地上生物量. 应用生态学报, 2006, 17(9):1557- 1562.
[3] 刘长成, 魏雅芬, 刘玉国, 郭柯. 贵州普定喀斯特次生林乔灌层地上生物量. 植物生态学报, 2009, 33(4):698- 705.
[4] 郑绍伟, 唐敏, 邹俊辉, 慕长龙. 灌木群落及生物量研究综述. 成都大学学报:自然科学版, 2007, 26(3):189- 192.
[5] 曾慧卿, 刘琪璟, 马泽清, 曾珍英. 基于冠幅及植株高度的檵木生物量回归模型. 南京林业大学学报:自然科学版, 2006, 30(4):101- 104.
[6] 赵蓓, 郭泉水, 牛树奎, 孙武, 张志旭. 大岗山林区几种常见灌木生物量估算与分析. 东北林业大学学报, 2012, 40(9):28- 33.
[7] 袁素芬, 陈亚宁, 李卫红, 刘加珍, 孟丽红, 张丽华. 新疆塔里木河下游灌丛地上生物量及其空间分布. 生态学报, 2006, 26(6):1818- 1824.
[8] 卢振龙, 龚孝生. 灌木生物量测定的研究进展. 林业调查规划, 2009, 34(4):37- 40, 45- 45.
[9] 刘恩斌, 周国模, 姜培坤, 葛宏立, 杜华强. 生物量统一模型构建及非线性偏最小二乘辩识—以毛竹为例. 生态学报, 2009, 29(10):5561- 5569.
[10] 刘恩斌, 李永夫, 周国模, 施拥军, 莫路锋. 生物量精确估算模型与参数辨识方法及应用. 生态学报, 2010, 30(10):2549- 2561.
[11] 黎燕琼, 郑绍伟, 宿以明, 龚良春, 慕长龙. 岷江上游干旱河谷引种番麻的生态适应性及生物量预测模型. 生态学报, 2009, 29(9):4820- 4826.
[12] 杨昊天, 李新荣, 刘立超, 贾荣亮, 王增如, 李小军, 李刚. 荒漠草地4种灌木生物量分配特征. 中国沙漠, 2013, 33(5):1340- 1348.
[13] Weiner J. Allocation, plasticity and allometry in plants. Perspectives in Plant Ecology, Evolution and Systematics, 2004, 6(4):207- 215.
[14] Fabbro T, Korner C. Altitudinal differences in flower traits and reproductive allocation. Flora - Morphology, Distribution, Functional Ecology of Plants, 2004, 199(1):70- 81.
[15] 冯丽, 张景光, 张志山, 郭群, 李新荣. 腾格里沙漠人工固沙植被中油蒿的生长及生物量分配动态. 植物生态学报, 2009, 33(6):1132- 1139.
[16] 王政权, 郭大立. 根系生态学. 植物生态学报, 2008, 32(6):1213- 1216.
[17] 刘兴良, 马钦彦, 杨冬生, 史作民, 宿以明, 周世强, 刘世荣, 杨玉坡. 川西山地主要人工林种群根系生物量与生产力. 生态学报, 2006, 26(2):542- 551.
[18] 岳永杰, 余新晓, 慕长龙, 刘兴良, 宿以明, 郑绍伟. 岷江上游干旱河谷区人工林根系生物量及分布格局. 东北林业大学学报, 2008, 36(2):16- 18.
[19] 王向荣, 王政权, 韩有志, 谷加存, 郭大立, 梅莉. 水曲柳和落叶松不同根序之间细根直径的变异研究.植物生态学报, 2005, 29(6):871- 877.
[20] 王力朋, 晏紫伊, 李吉跃, 王军辉, 何茜, 苏艳, 陈博, 马建伟, 董菊兰. 指数施肥对楸树无性系生物量分配和根系形态的影响. 生态学报, 2012, 32(23):7452- 7462.
[21] 阿米娜木·艾力, 常顺利, 张毓涛, 仇瑶, 何平. 天山云杉森林土壤有机碳沿海拔的分布规律及其影响因素. 生态学报, 2014, 34(7):1626- 1634.
[22] 曾慧卿, 刘琪璟, 冯宗炜, 马泽清, 胡理乐. 红壤丘陵区林下灌木生物量估算模型的建立及其应用. 应用生态学报, 2007, 18(10):2185- 2190.
[23] 陈遐林, 马钦彦, 康峰峰, 曹文强, 张国华, 陈宗伟. 山西太岳山典型灌木林生物量及生产力研究. 林业科学研究, 2002, 15(3):304- 309.
[24] 杨昆, 管东生. 森林林下植被生物量收获的样方选择和模型. 生态学报, 2007, 27(2):705- 714.
[25] 曾慧卿, 刘琪璟, 马泽清, 张海清, 蔡哲, 曾珍英, 李轩然. 千烟洲灌木生物量模型研究. 浙江林业科技, 2006, 26(1):13- 17.
[26] 何列艳, 亢新刚, 范小莉, 高延, 冯启祥. 长白山区林下主要灌木生物量估算与分析. 南京林业大学学报:自然科学版, 2011, 35(5):45- 50.
[27] Paton D, Nunez J, Bao D, Munoz A. Forage biomass of 22 shrub species from Monfrague Natural Park (SW Spain) assessed by log-log regression models. Journal of Arid Environments, 2002, 52(2):223- 231.
[28] 孙书存, 钱能斌. 刺旋花种群形态参数的通径分析与亚灌木个体生物量建模. 应用生态学报, 1999, 10(2):155- 158.
[29] 林伟, 李俊生, 郑博福, 郭建明, 胡理乐. 井冈山自然保护区12种常见灌木生物量的估测模型. 武汉植物学研究, 2010, 28(6):725- 729.
[30] 张倩媚, 温达志, 叶万辉, 孔国辉. 南亚热带常绿阔叶林林下层植物的生物量及其测定方法的探讨. 生态科学, 2000, 19(4):62- 66.
[31] 王新云, 郭艺歌, 陈林, 刘任涛, 杨明秀, 谢腾腾. 荒漠草原不同林龄柠条灌丛生物量模型研究. 生物数学学报, 2013, 28(2):377- 383.
[32] 董道瑞, 李霞, 万红梅, 林海军. 塔里木河下游柽柳灌丛地上生物量估测. 西北植物学报, 2012, 32(2):384- 390.
[33] 曾凡江, 郭海峰, 刘波, 曾杰, 邢文娟, 张晓蕾. 多枝柽柳和疏叶骆驼刺幼苗生物量分配及根系分布特征. 干旱区地理, 2010, 33(1):59- 64.
[34] Schenk H J, Jackson R B. Rooting depths, lateral root spreads and below-ground/above-ground allometries of plants in water-limited ecosystems. Journal of Ecology, 2002, 90(3):480- 494.
[35] Osone Y, Tateno M. Applicability and limitations of optimal biomass allocation models:a test of two species from fertile and infertile habitats. Annals of Botany, 2005, 95(7):1211- 1220.
[36] 黎燕琼, 郑绍伟, 龚固堂, 陈俊华, 唐波, 朱自芳, 吴雪仙, 慕长龙. 不同年龄柏木混交林下主要灌木黄荆生物量及分配格局. 生态学报, 2010, 30(11):2809- 2818.
[37] 刘兴良, 郝晓东, 杨冬生, 刘世荣, 宿以明, 蔡小虎, 何飞, 马钦彦. 卧龙巴郎山川滇高山栎灌丛地上生物量及其模型. 生态学杂志, 2006, 25(5):487- 491.
[38] 刘瑞, 靳虎甲, 马全林, 王耀琳, 李银科, 孙涛, 宋德伟, 朱国庆. 甘肃景电灌区不同栽植年限枸杞生物量分配特征. 生态学杂志, 2012, 31(10):2493- 2500.
[39] Salas E, Ozier-Lafontaine H, Nygren P. A fractal root model applied for estimating the root biomass and architecture in two tropical legume tree species. Annals of Forest Science, 2004, 61(4):337- 345.
[40] 郭忠玲, 郑金萍, 马元丹, 韩士杰, 李庆康, 于贵瑞, 范春楠, 刘万德, 邵殿坤. 长白山几种主要森林群落木本植物细根生物量及其动态. 生态学报, 2006, 26(9):2855- 2862.
[41] 牛存洋, 阿拉木萨, 宗芹, 骆永明, 押田敏雄, 孙贵凡, 刘谦. 科尔沁沙地小叶锦鸡儿地上-地下生物量分配格局. 生态学杂志, 2013, 32(8):1980- 1986.
Biomass estimation modeling and adaptability analysis of organ allocation in six common shrub species in Tianshan Mountains forests, China
QIU Yao1, CHANG Shunli1,*, ZHANG Yutao2, WANG Wendong2, HE Ping1, WANG Huijie1, XIE Jin1
1KeyLaboratoryofOasisEcology,CollegeofResourceandEnvironmentScience,XinjiangUniversity,Urumqi830046,China2InstituteofForestEcology,XinjiangAcademyofForestry,Urumqi830063,China
Estimation modeling of the total biomass of shrubs is associated with certain difficulties. Moreover, merely studying the effect of organ allocation adaptability on shrub biomass is not sufficient. Thus, to investigate the effectiveness of biomass estimation modeling on organ allocation in plants, we focused on six common shrub species found in the forests of Tianshan Mountains, northwest China. To conduct a community investigation, we selected three sample areas in the eastern, central, and western regions of Tianshan Mountains. A number of whole standard plants from six shrub species were harvested to determine total biomass, which was divided into the, biomass of the roots, branches, and leaves, along with different classes of root diameter.D2H(the square of the diameter multiplied by height) andV(the area of the crown multiplied by height) were selected as the independent variables in the estimation models. Optimal estimation models for the total biomass of the six shrub species were constructed using regression analysis. These models were used to compare differences in total biomass allocation to vegetative organs and in biomass allocation to roots of each root diameter class in the shrub species. The following results were obtained. First, the total biomass ofBerberisheteropoda,Lonicerahispida, andCotoneastermelanocarpuswas approximately 8.48 to 9.01 kg, whereas that ofRosaspinosissima,Spiraeahypericifolia, andJuniperuspseudosabinawas approximately 2.71 to 3.20kg. Second, the optimal estimation models for the total biomass ofR.spinosissima,S.hypericifolia, andC.melanocarpuswere functions based onVas an independent variable, whereas those forB.heteropoda,L.hispida, andJ.pseudosabinawere functions based onD2Has an independent variable. The value ofR2for each estimation model was greater than 0.850, and all the models reached a high degree of accuracy at the 0.05 significance level. Third, differences in the proportional allocation of total biomass to roots and branches of the six shrub species were not significant, whereas that to leaves was significant at the 0.05 significance level. The root biomass of all six shrub species decreased with decreasing root diameter class, with significant differences in the proportional allocation of biomass to roots in roots with a diameter greater than 2 mm (significant at the 0.05 and 0.01 significance levels for roots with diameters greater than 20mm). For all examined shrub species, we conclude that the differences in total biomass allocation to vegetative organs and in biomass allocation to the roots of each diameter class reflect adaptive strategies to their respective habitats.
biomass modeling; root classification; adaptive strategy; shrub; Tianshan Mountains
新疆维吾尔自治区财政林业科技专项资金项目(xjlk(2013)001); 国家自然科学基金项目(31060075); 新疆自治区公益性科研院所基本科研业务费专项资金项目(《天山东部与西部灌木林生物量及生产力研究》201414)
2014- 04- 28; < class="emphasis_bold">网络出版日期:
日期:2015- 05- 19
10.5846/stxb201404280841
*通讯作者Corresponding author.E-mail:ecocsl@163.com
仇瑶, 常顺利,张毓涛,王文栋,何平,王慧杰, 谢锦.天山林区六种灌木生物量的建模及其器官分配的适应性.生态学报,2015,35(23):7842- 7851.
Qiu Y, Chang S L, Zhang Y T, Wang W D, He P, Wang H J, Xie J.Biomass estimation modeling and adaptability analysis of organ allocation in six common shrub species in Tianshan Mountains forests, China.Acta Ecologica Sinica,2015,35(23):7842- 7851.
