中国温带旱柳物候期对气候变化的时空响应
2015-02-07陈效逑张晴华尉杨平
陈效逑,庞 程,徐 琳,李 静,张晴华,尉杨平
1 北京大学城市与环境学院,地表过程分析与模拟教育部重点实验室,北京 100871 2 中国农业大学资源与环境学院,北京 100193
中国温带旱柳物候期对气候变化的时空响应
陈效逑1,*,庞 程1,徐 琳2,李 静1,张晴华1,尉杨平1
1 北京大学城市与环境学院,地表过程分析与模拟教育部重点实验室,北京 100871 2 中国农业大学资源与环境学院,北京 100193
为了揭示中国温带植物物候随时间变化和植物物候对气候变化响应的空间格局及其生态机制,利用52个站点1986—2005年的旱柳展叶始期、开花始期、果实成熟期、叶变色始期和落叶末期的物候数据,分析其时间序列的线性趋势,并通过建立基于最佳期间日均温的物候时间模型,确定物候发生日期对气温年际变化的响应。在研究的时段内,区域平均旱柳展叶始期、开花始期和果实成熟期的发生日期分别以-4.2 d/10 a、-3.8 d/10 a和-3.3 d/10 a的平均速率显著提前,而区域平均旱柳叶变色始期和落叶末期的发生日期则分别呈不显著推迟和以2.4 d/10 a的平均速率显著推迟的趋势。单站展叶始期、开花始期和果实成熟期发生日期的线性趋势以提前为主,显著提前的站点分别占40%、41%和29%;叶变色始期发生日期呈显著提前和显著推迟趋势的站点数相当,分别占17%和19%;落叶末期发生日期的线性趋势以推迟为主,显著推迟的站点占23%。各站展叶始期、开花始期和果实成熟期发生日期的线性趋势空间序列与相应的最佳期间日均温的线性趋势空间序列之间呈显著负相关,表明一个站点前期气温升高的速率越快,该站这些物候期发生日期提前的速率就越快。在物候期对气温年际变化的响应方面,区域平均春季最佳期间日均温每升高1℃,展叶始期、开花始期和果实成熟期的发生日期分别提前3.08 d、2.83 d和3.54 d;区域平均秋季最佳期间日均温每升高1℃,叶变色始期和落叶末期的发生日期分别推迟1.69 d和2.28 d。单站展叶始期和落叶末期发生日期对气温年际变化的响应表现出在温暖地区的站点比在寒冷地区的站点更为敏感的特点。总体上看,基于日均温的物候时间模型对春、夏季物候期的模拟精度明显高于对秋季物候期的模拟精度。建立了基于最佳期间日均温和日累积降水量的改进秋季物候模型,该模型使旱柳叶变色始期和落叶末期的模拟精度显著提高。由此可见,旱柳叶变色始期和落叶末期的发生日期受到前期气温和降水量的综合影响。
植物物候;旱柳;线性趋势;时间响应;空间格局;敏感性;气候变化
诊断陆地植被的物候变化对于评价生态系统对近期气候变化的快速响应[1- 2],确定植被的碳吸收期[3- 8]和地表与大气之间水分、能量的季节性交换特征[9- 10]及其对全球碳循环和气候变化的反馈作用[11],具有重要的科学意义。大量研究表明,近几十年来,欧洲、北美和亚洲一些植物的春季物候如芽膨大、展叶和开花呈显著提前的趋势,秋季物候如叶变色和落叶呈显著推迟的趋势[12- 18],生长季节则呈显著延长的趋势[13,16- 18]。与上述研究结果不同,在俄罗斯的西北部,春季物候呈不显著提前或推迟的趋势,而秋季物候则呈显著提前或推迟的趋势[19]。在影响植物物候发生期的环境因子方面,大部分研究表明,树木的春季物候发生期主要受到前期气温的控制,通常某年春季气温越高,植物物候发生期越早;春季气温越低,植物物候发生期越晚[12, 16,18,20- 21]。此外,一些研究结果显示,树木的秋季物候发生期也受到前期气温的影响,通常某年秋季气温越高,植物物候发生期越晚;秋季气温越低,植物物候发生期越早[16,18, 20- 21]。诊断植物物候发生期与气温之间关系的传统方法是,计算某一物候发生期平均日期所在月份和此前若干月份的平均气温时间序列与该物候发生日期时间序列之间的简单相关系数和回归系数[12, 20- 21]。利用前期月平均气温作为自变量虽然简单易行,但往往并不确切,因为一个物候现象的发生日期并不一定是由前期完整月份的平均气温所驱动的,而是由与它相关程度最高的一段期间内的日平均气温所决定的,这个相关程度最高的期间可称为最佳期间。为了更加准确和合理地确定植物物候发生日期与气温之间的统计关系,Chen和Xu提出了最佳期间气温-物候时间模型[18],该模型的基本假设是,一个物候现象的发生日期主要受到它发生的最早和最晚日期之间以及最早日期之前一段最佳期间内的日平均气温的影响。据此,他们建立了榆树(Ulmuspumila)展叶始期和落叶末期与日均温之间的统计模型,作为诊断物候对气温变化响应的依据。结果显示,与基于月均温的模型相比,该模型显著提高了物候期对气温年际变化响应诊断的准确性。本文在上述研究的基础上,将该方法用于旱柳(Salixmatsudana)展叶始期、开花始期、果实成熟期、叶变色始期和落叶末期与气温关系的统计分析,以验证该模型对于其他植物种和物候期的适用性,并尝试改进其对秋季物候期的模拟。本文的主要目的是:(1)揭示1986—2005年旱柳物候期的线性趋势及其与气温线性趋势的空间相关性;(2)建立基于最佳期间日均温的物候时间模型,揭示旱柳物候期对气温年际变化的响应及其空间差异的成因;(3)建立基于最佳期间日均温和日累积降水量的改进物候时间模型,以揭示旱柳秋季物候期对前期气温和降水量的综合响应特征。
1 材料与方法
1.1 研究区域与物种选择
北半球温带地区是落叶阔叶植被的主要分布区,植物的物候变化非常显著,因此,通常作为研究植物物候对全球气候变化响应的重点区域[12- 16, 18- 24]。本文的研究区域包括中国气候区划中的中温带和暖温带两个气候带以及湿润、半湿润、半干旱和干旱四个气候大区[25]。受东部季风气候和西北干旱气候的共同影响,该地区水热条件具有显著的季节变化和空间差异,植物物候的时空变化亦非常明显[18,26- 27],因此,适于进行大尺度、连续时间序列的植物物候对气候变化的响应研究。
旱柳(Salixmatsudana)隶属杨柳科(Salicaceae)柳属(Salix),为落叶乔木。该树种喜光,喜湿润的土壤,且能耐干旱、水湿和寒冷,是中国温带地区分布广泛的常见树种和造林树种,一般生长在海拔小于1600 m的平原、山地溪流两侧、水分条件较好的沙丘及丘间低地[28]。鉴于在中国气象局农业气象观测网的物候观测记录中,旱柳的物候观测站点较多且观测数据也比较完整,因此,本文选择旱柳作为指示植物种,来揭示中国温带植物物候随时间变化和植物物候对气候变化响应的空间格局及其生态机制。
1.2 物候与气象数据
所用旱柳物候数据取自中国气象局农业气象观测网的自然物候观测数据集,该物候观测网始建于1980年,是中国地面站点最多的物候观测网络系统[29],观测内容包括49种木本植物、10种草本植物和11种动物的物候期以及气象和水文现象[30]。为了揭示旱柳生长发育不同阶段的时序变化特征及其对气候变化的响应,选择展叶始期、开花始期、果实成熟期、叶变色始期和落叶末期作为指示物候期,其中,展叶始期的观测标准是“观测树上有个别枝条上的芽出现第一批平展的叶片”;开花始期的观测标准是“观测树上有一朵或同时几朵花的花瓣开始完全开放”;果实成熟期的观测标准是“当观测的树木上有一半的果实变为成熟的颜色”;叶变色始期的观测标准是“观测的树木叶子在秋天第一批开始变色”;落叶末期的观测标准为秋季“树上的叶子几乎全部脱落”[30]。
为了保证数据的质量,根据物候现象发生的顺序相关性规律[31],对研究区域内各站点旱柳物候发生日期的数据进行了系统的审查,在对错误数据进行剔除的基础上,以1986—2005年期间各物候期的时间序列大于或等于16a为标准,选出52个样本站点。各物候期符合此标准的站点数不等,分别为展叶始期50个站、开花始期49个站、果实成熟期31个站、叶变色始期42个站和落叶末期48个站,这些站点覆盖了除沙漠和高山区以外的大部分中温带和暖温带地区且分布比较均匀,具有较好的区域代表性(图1)。
气象数据来源于中国气象数据共享服务网(http://cdc.cma.gov.cn/),包括研究区域内1986—2005年343个站点的逐日平均气温和降水量数据。由于在少量物候观测站点附近没有平行的气象观测,为了获得这些站点的日均温和日降水量数据,利用气象插值软件ANUSPLIN 4.2[32]和8 km×8 km分辨率的数字高程(DEM)数据,对1986—2005年逐日地面观测的日均温和日降水量进行空间插值,进而,根据缺失气象数据的物候站点经、纬度信息提取当地逐日日均温和日降水量数据,从而获得全部物候站点所在地1986—2005年的完整日均温和日降水量时间序列数据。
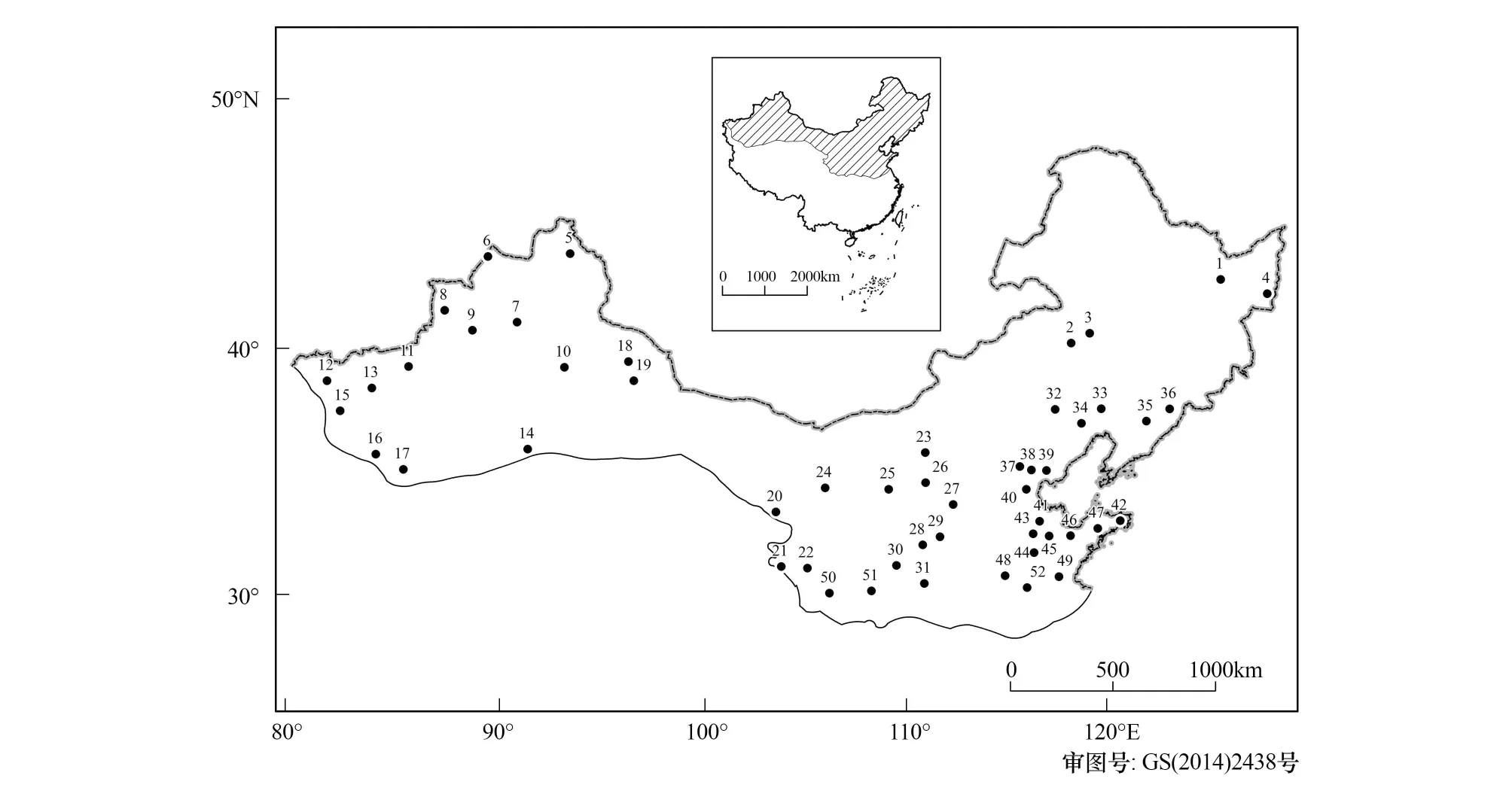
图1 旱柳物候站点的空间分布
1.3 统计方法
本文采用最佳期间日均温的计算方法[18]进行旱柳物候期的统计模拟,其基本计算过程为:首先,将某站点(或区域平均)1986—2005年旱柳物候发生日期(以从1月1日算起的顺序天数表示,简称序日)时间序列中最早和最晚日期之间的时段定义为基本长度期间(basic length period,bLP);其次,从基本长度期间的最早日期向后(向更早的时间方向)以1 d为步长取一系列滑动长度期间(moving length period,mLP),设定滑动长度期间以60 d为限,因为60 d以前的气温状况一般对植物物候的发生影响较小[18];第三,依次计算所有完整长度期间内(bLP+1 d,bLP+2 d,bLP+3 d,…,bLP+60 d)日平均气温的时间序列,完整长度期间(length period,LP)定义为:
LP=bLP+mLP
(1)
第四,分别计算所有完整长度期间内的日平均气温时间序列与旱柳物候发生期时间序列之间的相关系数;第五,将最大相关系数(绝对值)所对应的日平均气温时间序列定义为最佳长度期间的日均温时间序列,并据此建立该站(或区域平均)最佳期间气温-物候时间模型:
Phenophase=a+bToptimum
(2)
式中,Phenophase为物候发生日期,Toptimum为最佳期间日均温,a为回归方程截距,b为斜率。
在分析秋季物候期对温度和水分的综合响应方面,引入了日降水量因子,其滑动长度期间也以60 d为限。在寻找日均温和日累积降水量的最佳长度期间组合时,需依次计算所有完整长度期间内(bLP+1 d,bLP+2 d,bLP+3 d,…,bLP+60 d)日均温和日累积降水量的时间序列。进而,计算所有日均温和日累积降水量时间序列组合(60×60=3600)与物候发生日期时间序列的复相关系数。最后,将最大复相关系数所对应的日均温和日累积降水量时间序列定义为最佳长度期间的日均温和日累积降水量时间序列,并据此建立最佳期间气温、降水-物候时间模型:
Phenophase=a+b1Toptimum+b2Poptimum
(3)
式中,Poptimum为最佳期间日累积降水量,b1和b2为偏回归系数,其他符号同公式(2)。
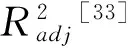
(4)
(5)
式中,obsi和prei为第i个年份的观测值和模拟(预测)值;n为年数。
2 结果
2.1 旱柳物候期的线性趋势变化
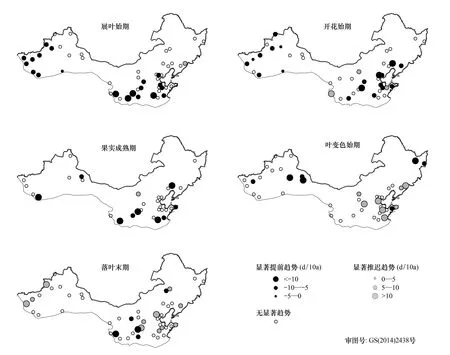
图2 1986—2005年单站旱柳物候期线性趋势的空间格局
从单站来看(图2),旱柳展叶始期呈显著提前趋势(P<0.05)的站点有20个,占站点总数的40%,其倾向率介于-3.8 d/10 a至-16.8 d/10 a之间,呈显著推迟趋势的站点有1个,其倾向率为7.5 d/10 a;开花始期呈显著提前趋势的站点也有20个(41%),其倾向率介于-3 d/10 a至-16.6 d/10 a之间,呈显著推迟趋势的站点有2个,其倾向率为5.1 d/10 a和12.1 d/10 a;果实成熟期呈显著提前趋势的站点有9个(29%),其倾向率介于-7.1 d/10 a至-16.8 d/10 a之间,呈显著推迟趋势的站点有1个,其倾向率为7.8 d/10 a;叶变色始期呈显著提前趋势的站点有7个(17%),其倾向率介于-5.9 d/10 a至-14.2 d/10 a之间,呈显著推迟趋势的站点有8个(19%),其倾向率介于5.1 d/10 a至15.6 d/10 a之间;落叶末期呈显著提前趋势的站点有5个(10%),其倾向率介于-5.6 d/10 a至-14 d/10 a之间,呈显著推迟趋势的站点有11个(23%),其倾向率介于5.2 d/10 a至16.7 d/10 a之间。综上所述,在1986—2005年期间,旱柳物候期在大部分站点的线性趋势变化并不显著,相比之下,展叶始期和开花始期呈显著线性趋势变化的站点较多,且以显著提前的趋势为主;而果实成熟期、叶变色始期和落叶末期呈显著线性趋势变化的站点较少,其中,果实成熟期以显著提前的趋势为主,叶变色始期呈显著提前和推迟的站点数相近,落叶末期以显著推迟的趋势为主。
从区域平均来看,1986—2005年旱柳展叶始期、开花始期和果实成熟期均呈显著提前的趋势,其倾向率分别为-4.2 d/10 a(P<0.01)、-3.8 d/10 a(P<0.01)和-3.3 d/10 a(P<0.05);叶变色始期呈不显著推迟的趋势(P>0.1),落叶末期则呈显著推迟的趋势(P<0.05),其倾向率为2.4 d/10 a。
2.2 旱柳物候期对气温年际变化的响应
单站最佳期间气温-物候模型的建模结果显示(表1),旱柳展叶始期、开花始期和果实成熟期时间序列与相应的最佳期间日均温时间序列多呈负相关,即某个站点某一年的最佳期间日均温越高,则该站点该年的物候发生日期就越早。其中,展叶始期有48个站点(96%)的相关系数通过了显著性检验(P<0.05),对气温年际变化的响应速率为-1.85 d/℃至-9 d/℃,其余2个站点的负相关不显著;开花始期有40个站点(82%)的相关系数通过了显著性检验,对气温年际变化的响应速率为-1.98 d/℃至-7.93 d/℃,其余9个站点中有7个为负相关,2个为正相关,但均不显著;果实成熟期有21个站点(68%)的相关系数通过了显著性检验,对气温年际变化的响应速率为-1.93 d/℃至-9.54 d/℃,其余10个站点中有6个为负相关,4个为正相关,也均不显著。相反,旱柳叶变色始期和落叶末期时间序列与相应的最佳期间日均温时间序列多呈正相关,即某个站点某一年的最佳期间日均温越高,则该站点该年的物候发生日期就越晚。然而,大部分站点的相关系数均没有通过显著性检验,其中,叶变色始期仅有7个站点(17%)的相关系数通过了显著性检验,对气温年际变化的响应速率为3.2 d/℃至7.85 d/℃,落叶末期有12个站点(25%)的相关系数通过了显著性检验,对气温年际变化的响应速率为2.49 d/℃至7.91 d/℃。
表1 1986—2005年单站旱柳物候期与最佳期间日均温之间的回归分析
Table 1 Regression analyses betweenSalixmatsudanaphenophases and mean daily air temperatures within the optimum length periods at each station during 1986 to 2005
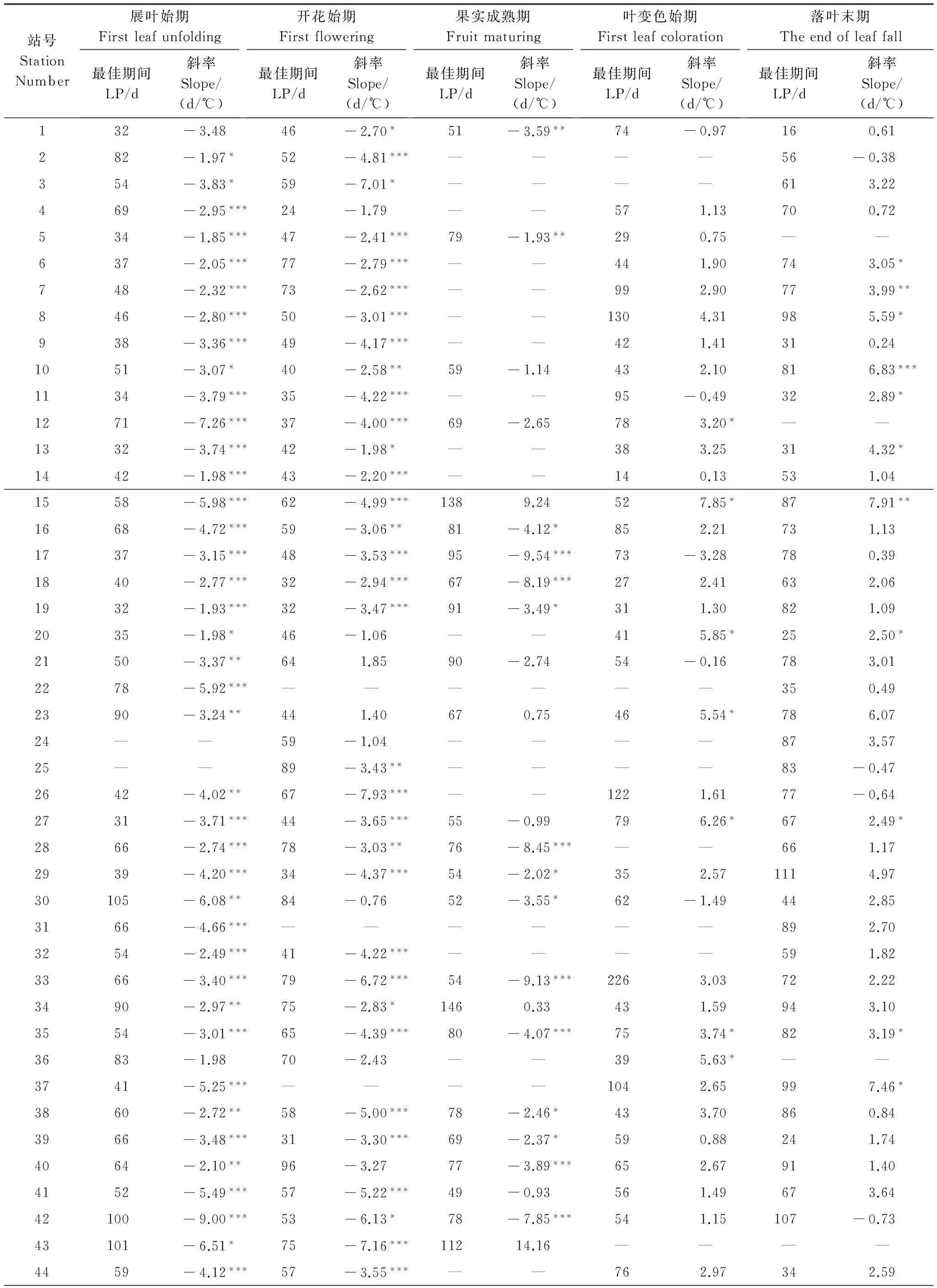
站号StationNumber展叶始期Firstleafunfolding最佳期间LP/d斜率Slope/(d/℃)开花始期Firstflowering最佳期间LP/d斜率Slope/(d/℃)果实成熟期Fruitmaturing最佳期间LP/d斜率Slope/(d/℃)叶变色始期Firstleafcoloration最佳期间LP/d斜率Slope/(d/℃)落叶末期Theendofleaffall最佳期间LP/d斜率Slope/(d/℃)132-3.4846-2.70*51-3.59**74-0.97160.61282-1.97*52-4.81***————56-0.38354-3.83*59-7.01*————613.22469-2.95***24-1.79——571.13700.72534-1.85***47-2.41***79-1.93**290.75——637-2.05***77-2.79***——441.90743.05*748-2.32***73-2.62***——992.90773.99**846-2.80***50-3.01***——1304.31985.59*938-3.36***49-4.17***——421.41310.241051-3.07*40-2.58**59-1.14432.10816.83***1134-3.79***35-4.22***——95-0.49322.89*1271-7.26***37-4.00***69-2.65783.20*——1332-3.74***42-1.98*——383.25314.32*1442-1.98***43-2.20***——140.13531.041558-5.98***62-4.99***1389.24527.85*877.91**1668-4.72***59-3.06**81-4.12*852.21731.131737-3.15***48-3.53***95-9.54***73-3.28780.391840-2.77***32-2.94***67-8.19***272.41632.061932-1.93***32-3.47***91-3.49*311.30821.092035-1.98*46-1.06——415.85*252.50*2150-3.37**641.8590-2.7454-0.16783.012278-5.92***——————350.492390-3.24**441.40670.75465.54*786.0724——59-1.04————873.5725——89-3.43**————83-0.472642-4.02**67-7.93***——1221.6177-0.642731-3.71***44-3.65***55-0.99796.26*672.49*2866-2.74***78-3.03**76-8.45***——661.172939-4.20***34-4.37***54-2.02*352.571114.9730105-6.08**84-0.7652-3.55*62-1.49442.853166-4.66***——————892.703254-2.49***41-4.22***————591.823366-3.40***79-6.72***54-9.13***2263.03722.223490-2.97**75-2.83*1460.33431.59943.103554-3.01***65-4.39***80-4.07***753.74*823.19*3683-1.9870-2.43——395.63*——3741-5.25***————1042.65997.46*3860-2.72**58-5.00***78-2.46*433.70860.843966-3.48***31-3.30***69-2.37*590.88241.744064-2.10**96-3.2777-3.89***652.67911.404152-5.49***57-5.22***49-0.93561.49673.6442100-9.00***53-6.13*78-7.85***541.15107-0.7343101-6.51*75-7.16***11214.16————4459-4.12***57-3.55***——762.97342.59
区域平均最佳期间气温-物候模型的建模结果显示,旱柳展叶始期、开花始期和果实成熟期时间序列与相应最佳期间日均温时间序列均呈显著负相关,最佳期间日均温每升高1℃,展叶始期提前3.08 d,开花始期提前2.83 d,果实成熟期提前3.54 d;而叶变色始期和落叶末期时间序列与相应最佳期间日均温时间序列均呈显著正相关,最佳期间日均温每升高1℃,叶变色始期推迟1.69 d,落叶末期推迟2.28 d。
2.3 旱柳物候期与气温线性趋势之间的空间相关
从图2可以看出,单站旱柳物候发生日期线性趋势的空间差异显著,为了揭示这种空间差异的影响因素,对每个站点上的旱柳物候发生日期线性趋势与相应的最佳期间气温线性趋势进行空间相关分析。结果表明,各站展叶始期、开花始期和果实成熟期发生日期的线性趋势与其最佳期间气温的线性趋势之间存在显著的负相关关系(P<0.05)。图3(a)—(c)中水平虚线以下的散点显示,最佳期间气温升温速率较快的站点,春季物候发生日期的提前速率(负值)也较快。与此相反,各站叶变色始期和落叶末期发生日期的线性趋势与其最佳期间气温的线性趋势之间相关关系均不显著(图3(d)和(e))。由此可见,1986—2005年旱柳春季物候发生日期线性趋势的空间格局主要由气温线性趋势的空间格局所决定,而旱柳秋季物候发生日期线性趋势的空间格局并非主要由气温线性趋势的空间格局所决定,其成因需要进一步研究和论证。
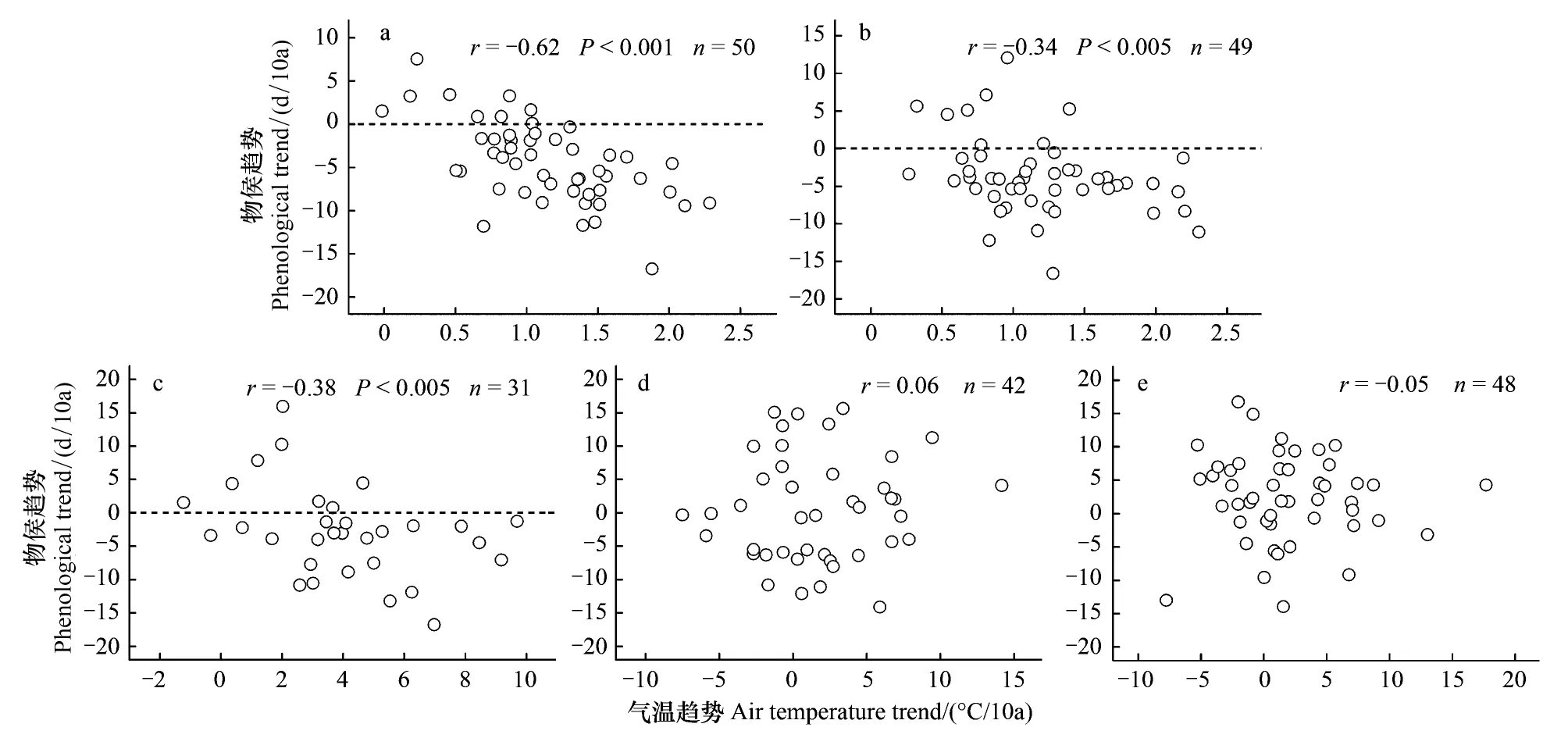
图3 1986—2005年单站旱柳物候期与气温线性趋势之间的空间相关分析
2.4 旱柳物候期对气温年际变化响应的空间差异归因
从表1可以看出,单站旱柳物候期对最佳期间气温年际变化响应速率(回归方程斜率)的空间差异显著,为了揭示这种空间差异的影响因素,以各站的多年平均气温作为衡量其热量状况的指标,计算了各站物候期对最佳期间气温的响应速率与当地多年平均气温的空间相关系数。结果表明,各站展叶始期对最佳期间气温的响应速率与多年平均气温呈显著负相关(P<0.001,图4(a)),落叶末期对最佳期间气温的响应速率与多年平均气温呈显著正相关(P<0.05,图4(e)),其余3个物候期对最佳期间气温的响应速率与多年平均气温的空间相关关系均不显著(图4(b)—(d))。由此可见,1986—2005年旱柳展叶始期和落叶末期(虚线以上的散点)对气温年际变化响应速率(绝对值)的空间格局主要受到多年平均气温空间格局的影响,表现为气候较暖站点的响应速率一般大于气候较冷站点的响应速率,而开花始期、果实成熟期和叶变色始期对气温年际变化响应速率的空间格局与多年平均气温的空间格局之间却不存在显著的统计关系,其控制因素有待进一步研究和论证。
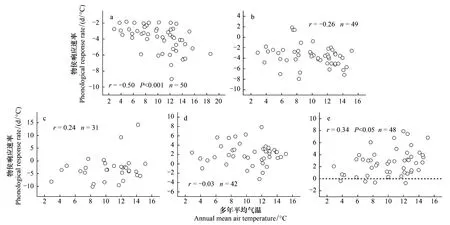
图4 1986—2005年单站旱柳物候期对气温的响应速率与多年平均气温的空间相关分析
3 讨论
(1)将本文的研究结果与同一时域空间内榆树物候期时间变化特征及其对气温变化的响应[18]进行比较可以看出,旱柳展叶始期和落叶末期的线性趋势与榆树展叶始期和落叶末期的线性趋势大致相当:二者展叶始期呈显著提前趋势的站点分别占总数的40%和44%,区域平均展叶始期的线性趋势分别为-4.2 d/10 a和-4 d/10 a;二者落叶末期呈显著推迟趋势的站点分别占总数的23%和24%,区域平均落叶末期的线性趋势分别为2.4 d/10 a和2.2 d/10 a。与榆树物候的研究结果相似,单站旱柳展叶始期线性趋势的倾向率与各地最佳期间气温线性趋势的倾向率呈显著负相关,即在升温速率较快的站点,树木展叶始期的提前速率也较快。应该指出,与榆树仅分析展叶始期和落叶末期的线性趋势相比,本文增加了旱柳开花始期、果实成熟期和叶变色始期时间序列的线性趋势分析,从而丰富了对不同植物物候期时间序列线性趋势差异性的认识。
(2)旱柳和榆树[18]对气温年际变化的响应及其空间差异特征也具有相似性。在96%和89%的站点上,旱柳和榆树展叶始期与最佳期间气温呈显著负相关,二者区域平均展叶始期对最佳期间气温的响应速率分别为-3.08 d/℃和-2.8 d/℃;在25%和26%的站点上,旱柳和榆树落叶末期与最佳期间气温呈显著正相关,二者区域平均落叶末期对最佳期间气温的响应速率分别为2.28 d/℃和2.1 d/℃。由此可见,基于日均温的最佳期间气温-物候时间模型具有描述气温与树木物候期之间统计关系的某种普适性,并且在准确性和合理性方面明显优于基于月均温的气温-物候时间模型。此外,各站旱柳和榆树展叶始期对最佳期间气温的响应速率(负值)均与当地多年平均气温呈显著负相关,而各站旱柳和榆树落叶末期对最佳期间气温的响应速率(正值)则均与当地多年平均气温呈显著正相关,表明气候较暖站点的展叶始期和落叶末期对气温年际变化的响应速率一般大于气候较冷站点。同样,与榆树仅分析展叶始期和落叶末期对气温年际变化的响应相比,本文增加了旱柳开花始期、果实成熟期和叶变色始期对气温年际变化的响应分析,从而丰富了不同植物物候期对气温年际变化响应差异性的认识。
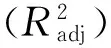
4 结论
(1)在1986—2005年期间,中国北方温带地区旱柳平均展叶始期、开花始期和果实成熟期分别以-4.2 d/10 a、-3.8 d/10 a和-3.3 d/10 a的平均速率显著提前,叶变色始期呈不显著推迟的趋势,落叶末期则以2.4 d/10 a的平均速率显著推迟。就单站而言,展叶始期、开花始期和果实成熟期以显著提前的趋势占优势,呈显著提前趋势的站点分别占40%、41%和29%;叶变色始期呈显著提前和显著推迟趋势的站点分别占17%和19%;落叶末期以显著推迟的趋势占优势,呈显著推迟趋势的站点占23%。
(2)各地展叶始期、开花始期和果实成熟期的线性趋势与其最佳期间气温的线性趋势之间负相关显著(P<0.05),即最佳期间气温升温速率较快的站点,这些物候期的提前速率也较快。与此相反,各地叶变色始期和落叶末期的线性趋势与其最佳期间气温的线性趋势之间相关均不显著。
(3)最佳期间气温-物候时间模型的模拟结果显示,区域平均最佳期间气温每升高1℃,展叶始期、开花始期和果实成熟期分别提前3.08 d、2.83 d和3.54 d;叶变色始期和落叶末期分别推迟1.69 d和2.28 d。就单站而言,大部分站点的展叶始期(96%)、开花始期(82%)和果实成熟期(68%)与最佳期间气温呈显著负相关,表明春季物候期的年际变化主要受到前期气温年际变化的影响;而大部分站点的叶变色始期和落叶末期与最佳期间气温的相关系数均未通过显著性检验,说明秋季物候期的早晚除受到前期气温年际变化的影响外,还与其他气象因子的状况有关。通过建立最佳期间气温、降水-秋季物候模型发现,前期气温和降水量对旱柳叶变色始期和落叶末期产生显著的综合影响,引入降水量明显地提高了模型对旱柳秋季物候期的模拟精度。
(4)各地展叶始期对最佳期间气温的响应速率与当地多年平均气温呈显著负相关(P<0.001),各地落叶末期对最佳期间气温的响应速率与当地多年平均气温呈显著正相关(P<0.05),其余3个物候期对最佳期间气温的响应速率与多年平均气温的相关关系均不显著。由此可见,气候较暖站点的展叶始期和落叶末期对气温年际变化的响应速率一般大于气候较冷站点。
致谢:感谢中国气象局气候资料室提供植物物候数据。
[1] Walther G R, Post E, Convey P, Menzel A, Parmesan C, Beebee T J C, Fromentin J M, Guldberg O H, Bairlein F. Ecological responses to recent climate change. Nature, 2002, 416(6879): 389- 395.
[2] Parmesan C. Influences of species, latitudes and methodologies on estimates of phenological response to global warming. Global Change Biology, 2007, 13(9): 1860- 1872.
[3] Goulden M L, Munger J W, Fan S M, Daube B C, Wofsy S C. Exchange of carbon dioxide by a deciduous forest: response to interannual climate variability. Science, 1996, 271(5255): 1576- 1578.
[4] Black T A, Chen W J, Barr A G, Arain M A, Chen Z, Nesic Z, Hogg E H, Neumann H H, Yang P C. Increased carbon sequestration by a boreal deciduous forest in years with a warm spring. Geophysical Research Letters, 2000, 27(9): 1271- 1274.
[5] White M A, Nemani R R. Canopy duration has little influence on annual carbon storage in the deciduous broad leaf forest. Global Change Biology, 2003, 9(7): 967- 972.
[6] Barr A G, Black T A, Hogg E H, Kljun N, Morgenstern K, Nesic Z. Inter-annual variability in the leaf area index of a boreal aspen-hazelnut forest in relation to net ecosystem production. Agricultural and Forest Meteorology, 2004, 126(3/4): 237- 255.
[7] Churkina G, Schimel D, Braswell B H, Xiao X M. Spatial analysis of growing season length control over net ecosystem exchange. Global Change Biology, 2005, 11(10): 1777- 1787.
[8] Baldocchi D. Breathing of the terrestrial biosphere: lessons learned from a global network of carbon dioxide flux measurement systems. Australian Journal of Botany, 2008, 56(1): 1- 26.
[9] Wilson K B, Baldocchi D D. Seasonal and interannual variability of energy fluxes over a broadleaved temperate deciduous forest in North America. Agricultural and Forest Meteorology, 2000, 100(1): 1- 18.
[10] Kljun N, Black T A, Griffis T J, Barr A G, Gaumont-Guay G, Morgenstern K, McCaughey J H, Nesic Z. Response of net ecosystem productivity of three boreal forest stands to drought. Ecosystems, 2007, 10(6): 1039- 1055.
[11] Sellers P J, Hall F G, Kelly R D, Black A, Baldocchi D, Berry J, Ryan M, Ranson K J, Crill P M, Lettenmaier D P, Margolis H, Cihlar J, Newcomer J, Fitzjarrald D, Jarvis P G, Gower S T, Halliwell D, Williams D, Goodison, B, Wickland D E, Guertin F E. BOREAS in 1997: Experiment overview, scientific results, and future directions. Journal of Geophysical Research: Atmospheres, 1997, 102(D24): 28731- 28769.
[12] Bradley N L, Leopold A C, Ross J, Huffaker W. Phenological changes reflect climate change in Wisconsin. Proceedings of the National Academy of Sciences of the United States of America, 1999, 96(17): 9701- 9704.
[13] Menzel A, Fabian P. Growing season extended in Europe. Nature, 1999, 397(6721): 659- 659.
[14] Beaubien E G, Freeland H J. Spring phenology trends in Alberta, Canada: links to ocean temperature. International Journal of Biometeorology, 2000, 44(2): 53- 59.
[15] Fitter A H, Fitter R S R. Rapid changes in flowering time in British plants. Science, 2002, 296(5573): 1689- 1691.
[16] Matsumoto K, Ohta T, Irasawa M, Nakamura T. Climate change and extension of the Ginkgo biloba L. growing season in Japan. Global Change Biology, 2003, 9(11): 1634- 1642.
[17] Gordo O, Sanz J J. Long-term temporal changes of plant phenology in the western Mediterranean. Global Change Biology, 2009, 15(8): 1930- 1948.
[18] Chen X Q, Xu L. Phenological responses of Ulmus pumila (Siberian elm) to climate change in the temperate zone of China. International Journal of Biometeorology, 2012, 56(4): 695- 706.
[19] Kozlov M V, Berlina N G. Decline in length of the summer season on the Kola Peninsula, Russia. Climatic Change, 2002, 54(4): 387- 398.
[20] Menzel A. Plant phenological anomalies in Germany and their relation to air temperature and NAO. Climatic Change, 2003, 57(3): 243- 263.
[21] Gordo O, Sanz J J. Impact of climate change on plant phenology in Mediterranean ecosystems. Global Change Biology, 2010, 16(3): 1082- 1106.
[22] Wolfe D W, Schwartz M D, Lakso A N, Otsuki Y, Pool R M, Shaulis N J. Climate change and shifts in spring phenology of three horticultural woody perennials in northeastern USA. International Journal of Biometeorology, 2005, 49(5): 303- 309.
[23] Schwartz M D, Chen X Q. Examining the onset of spring in China. Climate Research, 2002, 21(2): 157- 164.
[24] 陈效逑, 韩建伟. 我国东部温带植物群落的季相及其时空变化特征. 植物生态学报, 2008, 32(2): 336- 346.
[25] 中国气象局. 中华人民共和国气候图集. 北京: 中国地图出版社, 1979: 222- 223.
[26] Xu L, Chen X Q. Spatial modeling of the Ulmus pumila growing season in China′s temperate zone. Science China Earth Sciences, 2012, 55(4): 656- 664.
[27] Chen X Q, Xu L. Temperature controls on the spatial pattern of tree phenology in China′s temperate zone. Agricultural and Forest Meteorology, 2012, 154- 155: 195- 202.
[28] 华北树木志编写组. 华北树木志. 北京: 中国林业出版社, 1984: 89- 90.
[29] Hudson I L, Keatley M R. Phenological Research: Methods for Environmental and Climate Change Analysis. Dordrecht-Heidelberg-London-New York: Springer, 2009: 35- 37.
[30] 国家气象局. 农业气象观测规范. 北京: 气象出版社, 1993: 136- 141.
[31] 陈效逑, 杨国栋. 北京地区的物候日历及其应用. 北京: 首都师范大学出版社, 1995: 7- 8.
[32] Hutchinson M F. Anusplin Version 4. 2 User Guide, 2002.
[33] 陈彦光. 地理数学方法: 基础和应用. 北京: 科学出版社, 2011: 61- 61.
[34] Chen X Q, Pan W F. Relationships among phenological growing season, time-integrated normalized difference vegetation index and climate forcing in the temperate region of eastern China. International Journal of Climatology, 2002, 22(14): 1781- 1792.
[35] Chen X Q, Hu B, Yu R. Spatial and temporal variation of phenological growing season and climate change impacts in temperate eastern China. Global Change Biology, 2005, 11(7): 1118- 1130.
[36] Dufrêne E, Davi H, François C, Le Maire G, Le Dantec V, Granier A. Modelling carbon and water cycles in a beech forest: Part I: model description and uncertainty analysis on modelled NEE. Ecology Modelling, 2005, 185(2/4): 407- 436.
[37] Heide O, Prestrud A K. Low temperature, but not photoperiod, controls growth cessation and dormancy induction and release in apple and pear. Tree Physiology, 2005, 25(1): 109- 114.
[38] Delpierre N, Dufrêne E, Soudani K, Ulrich E, Cecchini S, Boé J, François C. Modelling interannual and spatial variability of leaf senescence for three deciduous tree species in France. Agricultural and Forest Meteorology, 2009, 149(6/7): 938- 948.
Spatiotemporal response ofSalixmatsudana′s phenophases to climate change in China′s temperate zone
CHEN Xiaoqiu1*, PANG Cheng1, XU Lin2, LI Jing1, ZHANG Qinghua1, YU Yangping1
1CollegeofUrbanandEnvironmentalSciences,LaboratoryforEarthSurfaceProcessesoftheMinistryofEducation,PekingUniversity,Beijing100871,China2CollegeofResourcesandEnvironmentalSciences,ChinaAgriculturalUniversity,Beijing100193,China
Simulating temporal and spatial relationships between occurrence dates of plant phenophases and climatic factors is crucial not only for predicting phenological responses to climate change but also for identifying the carbon-uptake period and examining the seasonal exchanges of water and energy between land surface and atmosphere. The latter in turn affects the global carbon cycle and climate change. In order to reveal the spatial pattern and its ecological mechanism of temporal variation of plant phenology and plant phenology response to climate change in China′s temperate zone, we usedSalixmatsudana′s phenology data of first leaf unfolding (LU), first flowering (FF), fruit maturing (FM), first leaf coloration (LC) and the end of leaf fall (LF) at 52 stations during 1986—2005 to analyze the linear trend of phenological time series and identify the response of phenological occurrence dates to interannual temperature variations by establishing daily mean air temperature-based temporal phenology models. During the research period, regional mean occurrence dates ofSalixmatsudana′s LU, FF and FM significantly advanced at average rates of -4.2, -3.8 and -3.3 days per decade, respectively, whereas regional mean occurrence dates ofSalixmatsudana′s LC and LF indicated a nonsignificant delayed trend and a significant delayed trend at an average rate of 2.4 days per decade, respectively. At single stations, occurrence dates ofSalixmatsudana′s LU, FF and FM significantly advanced at 40%, 41% and 29% of stations, respectively; occurrence dates ofSalixmatsudana′s LC significantly advanced at 17% of stations and delayed at 19% of stations; occurrence dates ofSalixmatsudana′s LF significantly delayed at 23% of stations. Spatial series of linear trends in occurrence dates ofSalixmatsudana′s LU, FF and FM at all stations correlate negatively with spatial series of linear trends in corresponding daily mean air temperatures during the optimum length periods, namely, the quicker the preceding air temperatures increased at a station, the quicker the phenological occurrence dates advanced at the station. With respect to phenological response to interannual temperature variations, a 1°C increase in regional mean spring air temperatures during the optimum length periods may induce an advancement of 3.08 days, 2.83 days and 3.54 days in regional mean occurrence dates ofSalixmatsudana′s LU, FF and FM, respectively, whereas a 1°C increase in regional mean autumn air temperatures during the optimum length periods may cause a delay of 1.69 days and 2.28 days in regional mean occurrence dates ofSalixmatsudana′s LC and LF, respectively. At single stations, the response of occurrence dates ofSalixmatsudana′s LU and LF to interannual temperature variations was more sensitive at warmer locations than at colder locations. Overall, simulation precision of daily mean air temperature-based temporal phenology models for spring and summer phenophases was obviously higher than that for autumn phenophases. Therefore, we constructed daily mean air temperature and daily accumulative precipitation-based autumn phenology models. The revised models significantly enhanced simulation precision ofSalixmatsudana′s LC and LF. This indicates that occurrence dates ofSalixmatsudana′s LC and LF were triggered by the combined influence of preceding air temperature and precipitation.
plant phenology;Salixmatsudana; linear trend; temporal response; spatial pattern; sensitivity; climate change
国家自然科学基金项目(41071027,40871029)
2013- 10- 08;
2014- 06- 09
10.5846/stxb201310082411
*通讯作者Corresponding author.E-mail: cxq@pku.edu.cn
陈效逑,庞程,徐琳,李静,张晴华,尉杨平.中国温带旱柳物候期对气候变化的时空响应.生态学报,2015,35(11):3625- 3635.
Chen X Q, Pang C, Xu L, Li J, Zhang Q H, Yu Y P.Spatiotemporal response ofSalixmatsudana′s phenophases to climate change in China′s temperate zone.Acta Ecologica Sinica,2015,35(11):3625- 3635.
