小麦抽穗期与麦红吸浆虫成虫发生期的同步性及其受害程度
2015-02-07武予清段爱菊张自启刘长营刘顺通巩中军蒋月丽
武予清, 段爱菊, 张自启, 刘长营, 刘顺通, 苗 进,巩中军, 段 云, 蒋月丽, 李 彤
1 河南省农业科学院植物保护研究所,河南省农作物病虫害防治重点实验室,农业部华北南部作物有害生物综合治理重点实验室, 郑州 450002 2 洛阳市农林科学院, 洛阳 471000
小麦抽穗期与麦红吸浆虫成虫发生期的同步性及其受害程度
武予清1,*, 段爱菊2, 张自启2, 刘长营2, 刘顺通2, 苗 进1,巩中军1, 段 云1, 蒋月丽1, 李 彤1
1 河南省农业科学院植物保护研究所,河南省农作物病虫害防治重点实验室,农业部华北南部作物有害生物综合治理重点实验室, 郑州 450002 2 洛阳市农林科学院, 洛阳 471000
通过2012年在麦红吸浆虫发生田对小麦不同生育期进行套袋保护,明确了小麦抽穗期为麦红吸浆虫侵害的敏感期。采用生态位重叠率的计算方法,统计2012—2013年不同品种的抽穗期与麦红吸浆虫发生期的同步性,结果表明,同步性越高,产量损失越重,具有显著的相关关系。同时,品种抽穗期的成虫累计发生量与产量损失有显著的相关性。成虫发生期与抽穗期同步或吻合导致小麦受害而不是扬花期,所以在缺乏抗虫品种的情况下,应采取抽穗期化学保护进行防治。
小麦抽穗期; 麦红吸浆虫; 成虫发生期; 同步性; 受害程度
麦红吸浆虫(Sitodiplosismosellana(Gehin))是广泛分布于亚洲、北美和欧洲小麦产区的毁灭性的害虫。在我国元明清及民国均有为害小麦的记载,1950年代麦红吸浆虫主发区包括浙江、江西、江苏、安徽、河南、河北、山西、陕西等北纬32°—36°之间,成为由西北到东南的一条带状,横亘在我国东部大平原的中部[1]。自1950年代后期六六六大规模用于土壤处理,直到1983年六六六停止使用,这个时期麦红吸浆虫不再是小麦生产问题。1985年以后,麦红吸浆虫在我国冬小麦主产区黄淮海平原和西北地区再度发生,东部平原以河南、安徽、山东、河北、天津、北京最为严重,发生区向北推移至北纬40°[2]。
早期抗虫性能研究中,发现麦红吸浆虫雌成虫喜欢在扬花前的麦穗上的产卵,在感虫品种302上,着卵量占83%,在抽齐穗、抽出2寸和正开花的穗子接卵,被害率分别为31.46%、6.25%和5.55%,表明抽齐穗越久被害率越低,抽穗期是麦红吸浆虫最喜欢产卵的时期[3]。Reecher、Doane等观察到小麦从露脸到扬花期对麦红吸浆虫产卵都是敏感的[4- 5];尹楚道等在田间观察到破肚后到扬花前套袋,以及扬花后到灌浆期套袋,穗被害率分别为2.94%和70.00%[6]。直到1996年,Elliot and Mann报道了麦红吸浆虫在红春小麦TriticumaestivumL. cv katepwazai的抽穗期(G51- 59)到扬花期(G61- 69)不同阶段产卵后的为害情况[7],在品种Zadoks上,1992年观察到抽穗后期(G57- 59)产卵的麦穗上的幼虫量和被害率是扬花期(G61- 69)产卵的60—90倍;G57- 59(抽穗3/4到齐穗)、G61- 65(扬花前期)和G65- 70(扬花后期)的籽被害率分别是48.2%、3.2%和0.2%。Caafar和Volkmar在德国中部连续3a观察了小麦抽穗期与成虫发生期的情况[8],2007年麦红吸浆虫成虫高峰期在灌浆期(G73- 75),受害最轻(0.2头幼虫/穗);2008年高峰期在抽穗后期(G55- 59)受害最重(4.2头幼虫/穗);2009年高峰期在抽穗末到扬花初(G59- 61)受害为0.3头幼虫/穗。
我国麦红吸浆虫以冬小麦TriticumaestivumL.为主要侵害对象。鉴于小麦抽穗期对麦红吸浆虫侵害的敏感性,Dexter等提出加拿大小麦吸浆虫防治策略以抽穗期为主[9]。倪汉祥等提出了以小麦抽穗期抽穗70%—80%时作为成虫期防治的施药适期[10]。抽穗期喷药防治显著优于扬花期防治[2]。
但是长期以来,我国华北吸浆虫重发区一些生产部门一直认为小麦扬花期与吸浆虫羽化期吻合造成小麦的受害[11- 12],并采取“蛹期和成虫期防治并重,蛹期防治为主”的防治策略,而不是抽穗期化学保护的措施。由于蛹期的撒毒土防治费工、费时、费力,多采取成虫期(而不是抽穗期)喷雾防治,蛹期防治面积逐年减少,防治效果难以保证,造成田间虫源积累、虫情出现反复[13]。因此,研究成虫发生期与抽穗期的同步性、阐明抽穗期成虫的发生与小麦受害程度的关系,直接关系到我国麦红吸浆虫的科学防控。为此我们在河南麦红吸浆虫发生区开展了这方面的调查研究。
1 材料和方法
1.1 小麦不同生育期对麦红吸浆虫侵害的敏感性
(1)试验于2012年在河南省洛阳市洛龙区焦寨村麦红吸浆虫发生比较重的地块进行,试验分别在两块地中进行,小麦品种为洛麦24,2011年10月10日播种。试验田虫口基数分别为每样方(10cm×10cm×20cm)虫量135.5头。在4月20日至30日抽穗至扬花不同的时期开始套袋,套袋保护直到灌浆期:套袋数量打苞(挑旗期)60个;抽穗1/4、抽穗2/4、抽穗3/4和抽齐穗各90个;扬花前期(中间小穗开始吐雄蕊,吐雄蕊的小穗超过小穗总数的5%)和扬花后期(吐雄蕊的小穗超过小穗总数的50%)各90个。
(2)黄板记载成虫动态 同时在吸浆虫成虫发生期(4月中旬至5月中旬)用黄色粘板(100 mm×120 mm,佳多科工贸公司),监测吸浆虫的发生动态,黄板上沿齐于麦穗顶部,监测田设置10块黄板,双行,每行5块,间距10 m。逐日调查黄板上的吸虫数量,并更换粘虫板[ 14]。
(3)剥穗调查 在小麦乳熟期(杨花后15 d)未下雨之前剥穗调查的麦穗的总粒数、受害籽粒数及幼虫量,并计算产量损失率:
(1)
式中,W为穗上总虫数;G为总穗粒数;麦红红吸浆虫幼虫吃完1粒麦粒需4头[10]。
1.2 小麦不同品种抽穗期与麦红吸浆虫成虫的同步性及其受害程度
冬小麦品种来源于国家小麦产业技术体系病虫害防控研究室(依托西北农林科技大学植物保护学院)2011年和2012年秋分别种植417个和601个。试验地点在河南省洛阳市洛龙区焦寨村,10月中旬播种。土壤中麦红吸浆虫幼虫播前淘土,棋盘式取样,每小方平均幼虫量两年依次为135.5头和83.6头。每品种每小区种植1m双行,重复3次,记载每个品种的抽穗期[15]。同时进行成虫监测(方法同前所述)。
损失率调查,在小麦乳熟期(老熟幼虫入土前),每小区随机取10穗,每个品种共取30穗,每穗放入一纸袋内,带回室内逐穗、逐粒剥查麦粒中的幼虫数,计算每个鉴定品种的产量损失率。
采用同步性的计算,按照Geange等捕食者和被捕食者生态位重叠计算方法[16]。在时间的位点(K)上,物种i(麦穗)供吸浆虫利用的比例为pik,且:
(2)
同理,物种j(麦红吸浆虫)可用于有效侵害的成虫的比例为pjk,那么生态位重叠NOijk,即抽穗期与成虫发生期的同步性(Synchronization)为:
(3)
同步性最大值为1,最小值为0。
1.3 统计分析采用SPSS16.0进行。
2 结果与分析
2.1 不同生育期对麦红吸浆虫侵害的敏感性
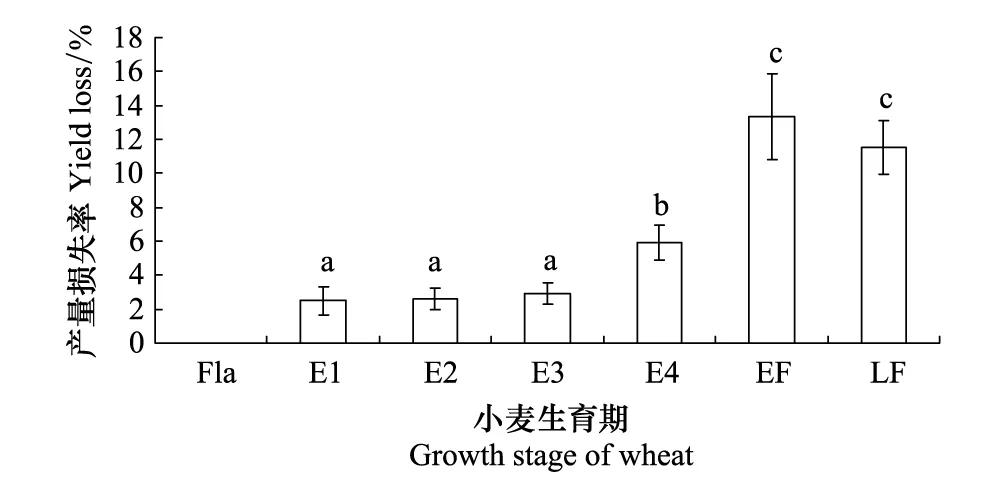
图1 洛麦24不同生育期套袋后麦红吸浆虫所致的产量损失
图1表明,挑旗期套袋保护,小麦没有受害,产量损失为零。抽齐穗期套袋的产量损失率,显著高于抽穗1/4、1/2和3/4三个时期套袋。扬花前期套袋与后期套袋之间的产量损失率无显著差异。扬花期套袋保护的产量损失率显著高于抽穗期,表明扬花期、穗期套袋保护已不能免受吸浆虫的侵害。结果表明抽穗期是麦红吸浆虫侵害的敏感期。
2012年套袋试验所用的品种洛麦24孕穗(挑旗)到扬花期正直吸浆虫成虫活动高峰期。
2.2 小麦不同品种抽穗期与麦红吸浆虫成虫发生期的同步性及其受害程度
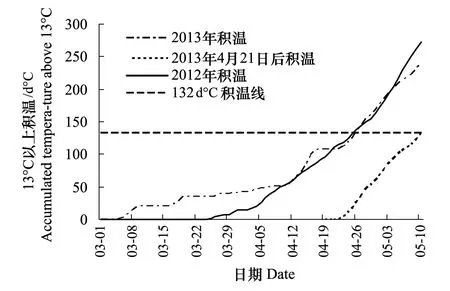
图2 河南省洛阳市洛龙区2012年和 2013年3—5月份13℃以上积温变化线
2013年吸浆虫成虫发生高峰日(5月10日)比2012年的高峰日(4月25日)推迟16d,这与2013年4月19日气温骤降至2℃关系密切。资料表明,在气温达到13℃以上时吸浆虫开始进入化蛹期[17]。图2表明2012年4月25日在13℃以上积温达到132d℃时达到成虫发生高峰。2013年4月19日的降温可能致使吸浆虫蛹遭到大量伤害,4月下旬并未出现成虫发生高峰,并致使小麦抽穗期的黄淮南部麦4区小麦出现大面积严重冻害减产。4月21日气温回升后未化蛹的吸浆虫幼虫重新进入蛹期,在13℃以上积温重新达到132d℃时出现了5月10日的成虫发生高峰。
2008—2012年的抗虫谱中品种抗性鉴定表明,除荆麦66表现高抗、中农28表现中抗外,其余均为感虫或高感[15],这使得感虫品种不同抽穗期与麦红吸浆虫成虫发生期的同步性及其受害程度的分析成为可能。
2012年成虫发生高峰期在4月下旬(图3),因此与抽穗期在4月22—28日的23个品种与麦红吸浆虫成虫发生期的同步性最高,为0.628,平均损失率78.1%:与抽穗期4月28日至5月2日的3个品种的同步性最低,为0.307,平均损失率11.7%。
2013年成虫发生高峰期在5月上旬(图4),因此与抽穗期在5月4—11日的3个品种的同步性最高,为0.783,平均损失率2.42%;与抽穗期在4月15—25日的11个品种同步性仅为0.062,平均损失率0.04%。
图5表明了品种抽穗期和成虫发生期的同步性,与小麦品种的产量损失率之间的显著相关性。同步性越高,品种的受害程度越重,这暗示着吸浆虫发生区吸浆虫发生的轻重与同步性关系密切。
2012和2013年的试验结果表明,在品种抽穗期,每个黄色粘板诱捕到的成虫累积量,与品种的产量损失率具有显著的正相关关系(图6)。因此,在小麦被吸浆虫侵害的抽穗期,成虫的发生量决定了品种的受害,即可以用抽穗期的成虫发生量决定是否进行吸浆虫防治。
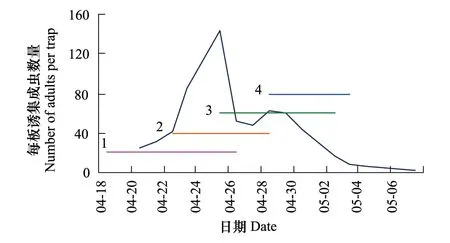
图3 2012年麦红吸浆虫成虫发生期与品种抽穗期
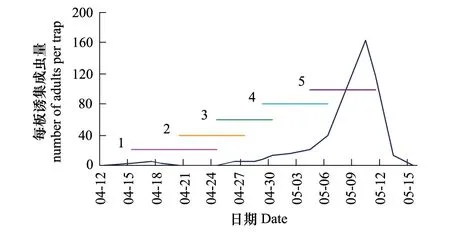
图4 2013年麦红吸浆虫成虫发生期与品种抽穗期

图5 品种的抽穗期与麦红吸浆虫成虫发生期的同步性,以及同步性与品种产量损失的相关性
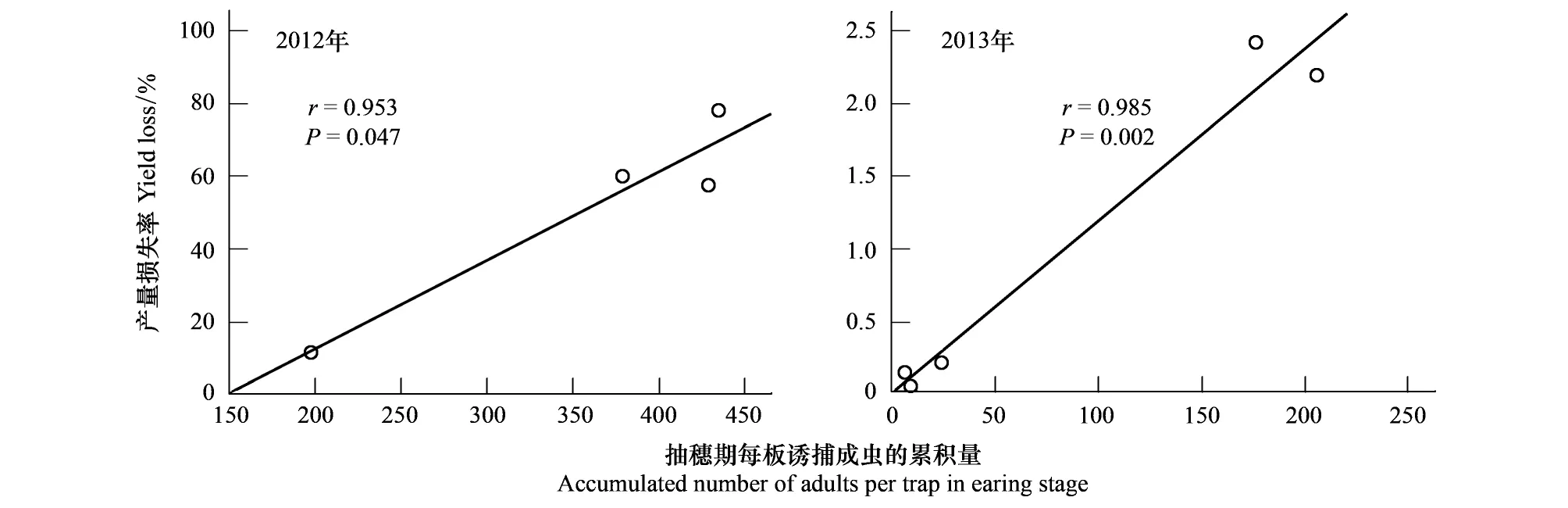
图6 品种抽穗期每黄板诱捕到的成虫累积量与产量损失的相关性
3 讨论
本文研究结果表明,麦红吸浆虫成虫发生期与抽穗期(而不是扬花期)的同步性或吻合程度的差异,导致小麦受害程度的不同。由于年度间气候的波动引起的同步性差异,从而导致小麦受害程度的发生较大的变化,是我国小麦主产区的普遍现象。定量研究同步性与吸浆虫发生程度的关系,对于研究气候变化下植物抗性以及昆虫与寄主植物相互作用关系的变化、作物病虫害发生趋势的预测具有重要的生态学意义和生产应用价值[18]。
研究结果同样表明了小麦品种对麦红吸浆虫具有“生态抗虫性”。如2008—2012年抗虫鉴定中大多数对吸浆虫感虫或高感的品种[15],在2013年由于吸浆虫成虫的出土高峰严重滞后而避开了吸浆虫的危害,均未表现出感虫;而抽穗较晚的品种,如冬性很强的蓝天系列品种,在洛阳的一般年份抽穗期比普通品种晚4—7 d而表现出“生态抗性或避虫性”,但在2013年与成虫发生期同步性较强的情况下,不再表现“生态抗性”。在我国小麦主产区,推迟的生育期会遭受更多的后期病虫害的危害,并影响秋作物的适期播种,因此这种以抽穗期推迟为标志的“生态抗性”被作为不良性状在育种过程中淘汰。
麦红吸浆虫作为麦类植物的专性寄生昆虫[19],有着一年一代或多年一代专性滞育的特征[20],成虫的出土期受温湿度条件影响极大[2]。在土壤湿度满足幼虫化蛹的前提下,年度间成虫的出土期和发生高峰期更多地受到温度的影响。加拿大Elliott等利用去除5月份小于20mm降雨的年份数据后,在气温5℃以上的积温与成虫出土多年数据的相关关系预测麦红吸浆虫的出土期,出土50%时的正负偏差为5.4 d,利用气温6和9℃以上的积温偏差更大[21]。因此,尽管积温控制着出土期的早晚,但是由于我国冬小麦抽穗期一般为7—10 d,就目前的研究报道来说,用积温预测出土期的误差,仍然容易错过小麦的抽穗期。吸浆虫成虫发生与气候调节的关系导致发生期不确定性为预测预报增加了难度。比利时Jacquemin等利用分段预测法,即越冬后从脱茧到活动幼虫在3℃以上满足250 d℃的积温后,当温度达到13℃(化蛹的起点温度)并且土壤湿度满足的情况下,在7 ℃以上160 d℃积温后为羽化高峰期,误差只有1 d。
由于小麦抽穗期是吸浆虫侵害的适期,在吸浆虫发生趋势的预测中,当年主栽品种小麦抽穗期的预测变得同样重要。此外,吸浆虫发生区北扩东移的原因,是否与同步性有关?目前已有的研究表明,江淮和黄淮南部地区麦红吸浆虫可随气流向华北长距离扩散,华北新发生区的吸浆虫来自江淮和黄淮南部[[22- 25]。鉴于1951—2009年59年间华北地区的气温上升了1.5℃[26],同时华北小麦品种的历史演化中,抽穗期的性状并没有太大的改变[27],那么是因为气候变暖引起华北北部吸浆虫成虫发生期提前,导致与小麦抽穗期由不同步性转为同步性?更深入的研究将为我国小麦吸浆虫发生区域的预测,以及研究气候变化对农作物病虫害发生规律的影响有着更为深刻的意义。
陈浩等报道麦红吸浆虫成虫发生总量与产量损失的存在线性关系[28]。本文的结果进一步表明,抽穗期成虫累计发生量与产量损失率存在显著的相关性。因为大田抽穗期一般为7—10 d,防治窗口非常“窄”,所以穗期的防治决策必须在抽穗初期做出。
在我国目前华北地区吸浆虫防治策略所采用“主攻蛹期(毒土处理),成虫扫残”的方针,由于蛹期毒土处理费工费力,不适于大面积“统防统治”的要求。因此,在我国目前极度在缺乏抗虫品种的情况下[15],穗期化学保护应该成为小麦吸浆虫发生区的主要措施。
[1] 杨平澜. 小麦吸浆虫的研究与防治 //昆虫学集刊.北京:科学出版社, 1959: 193- 211.
[2] 武予清, 苗进, 段云, 蒋月丽, 巩中军. 麦红吸浆虫的研究与防治. 北京:科学技术出版社, 2011: 1- 10, 56- 59,175- 175.
[3] 朱象三. 小麦抗吸浆虫性能的研究//中国植物保护科学. 北京:科学出版社, 1961: 374- 385.
[4] Reecher M M. The wheat midge in the Pacific Northwest. Circular United States Developmental Agriculture, 1945, 732: 1- 8.
[5] Doane J F, Olfert O O, Mukerji M K. Extraction precision of sieving and brine flotation for removal of wheat midge,Sitodiplosismosellana(Diptera: Cecidomyiidae) cocoons and larvae from soil. Journal of Economic Entomology, 1987, 80(1): 268- 271.
[6] 尹楚道, 潘锡康, 宋社吾, 张道环, 黄琦, 储常连. 小麦红吸浆虫种群动态、消长因素及为害调查研究. 安徽农学院学报, 1987, (3): 43- 48.
[7] Elliott R H, Mann L W. Susceptibility of red spring wheat,Triticumaestivuml. Cv. Katepwa, during heading and anthesis to damage by wheat midge,Sitodiplosismosellana(Géhin) (Diptera: cecidomyiidae). The Canadian Entomologist, 1996, 128(3): 367- 375.
[8] Gaafar N, Volkmar C. Evaluation of wheat ear insects in large scale field in central Germany. Agricultural Sciences, 2010, 1(2):68- 75.
[9] Dexter J E, Preston K R, Cooke L A, Morgan B C, Kruger J E, Kilborn R H, Elliott R H. The influence of orange wheat blossom midge (SitodiplosismosellanaGéhin) damage on hard red spring wheat quality and the effectiveness of insecticide treatments. Canadian Journal of Plant Sciences,1987, 67(3): 697- 712.
[10] 倪汉祥, 丁红建, 郭予元. 小麦红吸浆虫种群动态及综合治理技术体系成果研究回顾与展望 //成卓敏. 植物保护科技创新与发展.北京:中国农业科学技术出版社, 2008: 37- 44.
[11] 王书芳, 霍春玲, 李金凤, 申景梅, 王瑞海, 杨淑敏. 魏县小麦吸浆虫为害上升原因分析及综防措施. 中国植保导刊, 2012, 32(2): 39- 41.
[12] 王维莲, 江彦军, 李光. 2013年石家庄市小麦吸浆虫重发原因及防治对策. 现代农村科技, 2013, (11): 33- 34.
[13] 解丽娜, 张小龙, 李虎群, 张艳刚. 安新县2012年小麦吸浆虫发生特点及防治对策. 中国农技推广, 2013, 29(7): 45- 45, 9- 9.
[14] 武予清, 赵文新, 蒋月丽, 段云. 小麦红吸浆虫成虫的黄色粘板监测. 植物保护学报, 2009, 36(4): 381- 382.
[15] 武予清, 段爱菊, 张自启, 刘长营, 刘顺通, 苗进, 蒋月丽, 段云, 巩中军, 李彤. 小麦品种的麦红吸浆虫抗性分级方法及抗性评价. 作物学报, 2013, 39(12): 1271- 1276.
[16] Geange S W, Pledger S, Burns K C,Shima JS. A unified analysis of niche overlap incorporating data of different types. Methods in Ecology and Evolution, 2011, 2(2): 175- 184.
[17] Jacquemin G, Chavalle S, De ProftMi. Forecasting the emergence of the adult orange wheat blossom midge,Sitodiplosismosellana(Géhin) (Diptera: Cecidomyiidae) in Belgium. 2014, Crop Protection, 58 (1): 6- 13.
[18] Parmesan C. Ecological and evolutionary responses to recent climate change. Annual Review of Ecology, Ecology,and Systematics,2006,37(1): 637- 669.
[19] 武予清, 刘顺通, 段爱菊, 刘长营, 张自启, 蒋月丽, 段云. 河南西部小麦红吸浆虫禾本科寄主植物的记述. 植物保护, 2010, 36(5): 138- 140.
[20] Gong Z J, Wu Y Q, Miao J, Duan Y, Jiang Y L, Li L. Global transcriptome analysis of orange wheat blossom midge,Sitodiplosismosellana(Gehin) (Diptera: Cecidomyiidae) to identify candidate transcripts regulating diapause. PLOS One, 2013, 8(8): e71564.
[21] Elliott R H, Mann L, Olfert O. Calendar and degree-day requirements for emergence of adult wheat midge,Sitodiplosismosellana(Géhin) (Diptera: Cecidomyiidae) in Saskatchewan, Canada. Crop Protection, 2009, 28(7): 588- 594.
[22] 苗进, 武予清, 郁振兴, 陈华爽, 刘顺通, 蒋月丽, 段云. 麦红吸浆虫随气流远距离扩散的轨迹分析. 昆虫学报, 2011, 54(4): 432- 436.
[23] Miao J, Wu Y Q, Gong Z J, He Y Z, Duan Y, Jiang Y L. Long-distance wind-borne dispersal ofSitodiplosismosellanaGéhin (Diptera: Cecidomyiidae) in northern China. Journal of Insect Behavior, 2013, 26(1): 120- 129.
[24] Hao Y N, Miao J, Wu Y Q, Gong Z J, Jiang Y L, Duan Y, Li T, Cheng W N, Cui J X. Flight performance of the orange wheat blossom midge (Diptera: Cecidomyiidae). Journal of Economic Entomology, 2013, 106(5): 2043- 2047.
[25] Duan Y, Wu Y Q, Luo L Z, Miao J, Gong Z J, Jiang Y L, Li T. Genetic diversity and population structure ofSitodiplosismosellanain northern China. PLOS One, 2013, 8(11): e78415.
[26] 张一驰, 吴凯, 于静洁, 夏军. 华北地区1951—2009 年气温、降水变化特征. 自然资源学报,2011, 26(11): 1931- 1940.
[27] 庞春明, 郭北海. 用灰色关联度分析河北省冬小麦品种的演变规律. 国外农学-麦类作物,1994, (1): 36- 39.
[28] 陈浩, 李怡萍, 高宏利, 薛改妮, 肖智辉, 刘彦飞, 仵均祥. 小麦吸浆虫发生与小麦产量损失间的关系. 西北农业学报, 2011, 20(7): 37- 40.
The synchronization of ear emerging stages of winter wheat with occurrent periods of the orange wheat blossom midge,Sitodiplosismosellana(Gehin)(Diptera: Cecidomyiidae) adults and its damaged level
WU Yuqing1,*, DUAN Aiju2, ZHANG Ziqi2, LIU Changying2, LIU Shuntong2, MIAO Jin1, GONG Zhongjun1,DUAN Yun1, JIANG Yueli1, LI Tong1
1InstituteofPlantProtection,HenanAcademyofAgriculturalSciences,KeyLaboratoryofCropPestsControlofHenanProvince,KeyLaboratoryofPestManagementinSouthofNorth-China,MinistryofAgriculture,Zhengzhou450002,China2LuoyangMunicipalAcademyofAgriculturalandForestrySciences,Luoyang471000,China
The orange wheat blossom midge (OWBM),Sitodiplosismosellana(Gehin), is one of the most important pests of winter wheat in North China. It often feeds on developing wheat kernels and can cause a serious yield loss of this cereal. There were 2.4 million hectares with OWBM occurrence in 2013 in China. On the other hand, from the 1950s to the present time, the northern boundary of the OWBM occurrence area has shifted towards the pole by about four degrees of latitude (440 km) in North China. The northern area of North China, including Hebei, Beijing, Tianjin and northwestern Shandong, has become a novel occurrence region.This investigation was carried out on bagged ears of flag leaf, ear emergent and flowering stages of the winter wheat cultivar, Luomai 24, protected from the OWBM in fields in 2012 at Jiaozhaivillage, Luolong district, Luoyang city, Henan. The OWBM adults were monitored in April and May with yellow sticky traps, and the adult period of OWBM was found to overlap with the ear emergent and flowering stages of the cultivar. There were no damaged bagged ears in the flag leaf stage. The yield losses in bagged ears from 1/4 to 3/4 ear emergence were significantly less than in the fully ear emergent stage, but there were no significant differences between bagged early and late flowering stages. However, the yield loss in bagged flowering ears (average 10.79%) was significantly higher than in the bagged ear emergent stage (average 3.48%).The result revealed that emerging ears are susceptible to OWBM infestation. The damage level was evaluated in terms of synchronization of ear emergence stages of winter wheat cultivars with occurrence periods of OWBM adults in the 2012 and 2013 crop growing seasons. More than 400 susceptible cultivars were evaluated. Because of a 2℃ lower temperature on 19 April 2013, the adult period ran from 4 May to 13 May that year. Synchronization was calculated using the niche overlap formula (Geange et al 2011). In the 2012 growing season, the maximum synchronization value for 23 cultivars was 0.628 when the ear emergent stage ran from 22 April to 28 April, and the average yield loss was 78.1%. The minimum synchronization value was 0.307 for three cultivars when the ear emergent stage ran from 28 April to 2 May, and the average yield loss was 11.7%. In the 2013 growing season, the maximum synchronization value for three cultivars was 0.783 when the ear emergent stage ran from 4 May to 11 May, and the average yield loss was 2.41%. The minimum value for 11 cultivars was 0.062 when the ear emergent stage ran from 15 April to 25 April, and the average yield loss was 0.04%. The results obtained indicate that higher synchronization values led to more serious yield losses and there was a significant relationship between them. The yield losses of wheat cultivars showed a significant positive correlation with the total number of OWBM adults during the ear emergent stage. This conclusion indicates that the control strategy should be aimed at chemical protection of the ear emergent stage in China until resistant wheat cultivars are planted, because the infestive OWBM occurrence period is in the ear emergent stages of winter wheat, and not the flowering stage. At the same time, the result may help to explain why the OWBM occurrence area has shifted towards the pole because the average winter and spring temperature has risen by 1.5℃ since the 1950s in the northern area of North China, and there might now be an synchronous or matched status between wheat ear emergent stage and OWBM adults in the northern area of North China.
wheat ear emerging stage;Sitodiplosismosellana; adult occurrence period; synchronization; damage level
国家现代农业产业技术体系(CARS- 03); 国家自然科学基金(31201513)
2013- 08- 11;
2014- 06- 12
10.5846/stxb201308112060
*通讯作者Corresponding author.E-mail: yuqingwu36@hotmail.com
武予清, 段爱菊, 张自启, 刘长营, 刘顺通, 苗进,巩中军, 段云, 蒋月丽, 李彤.小麦抽穗期与麦红吸浆虫成虫发生期的同步性及其受害程度.生态学报,2015,35(11):3548- 3554.
Wu Y Q, Duan A J, Zhang Z Q, Liu C Y, Liu S T, Miao J, Gong Z J,Duan Y, Jiang Y L, Li T.The synchronization of ear emerging stages of winter wheat with occurrent periods of the orange wheat blossom midge,Sitodiplosismosellana(Gehin) (Diptera: Cecidomyiidae) adults and its damaged level.Acta Ecologica Sinica,2015,35(11):3548- 3554.
