青海云杉无性系木材性状的遗传变异1)
2015-02-07李青粉王军辉李登平胡长寿祁生秀
李青粉 王军辉 李登平 胡长寿 祁生秀
(国家林业局林木培育重点实验室(中国林业科学研究院林业研究所),北京,100091) (青海省大通县东峡林场)
青海云杉无性系木材性状的遗传变异1)
李青粉 王军辉 李登平 胡长寿 祁生秀
(国家林业局林木培育重点实验室(中国林业科学研究院林业研究所),北京,100091) (青海省大通县东峡林场)
采用相关分析、主成分分析和聚类分析方法对18个青海云杉无性系的生长性状和材性性状进行遗传变异研究,并通过多性状选择筛选出综合性状优良的无性系。结果表明:材性性状无性系间均存在较大的变异,除早材管胞长度与早材双壁厚度外,其他性状晚材的变异系数均大于早材;树脂道比量、生长轮宽度和晚材径向管胞腔径的变异系数较大(21.0%~34.2%),管胞比量和基本密度的变异系数较小(0.50%和3.9%)。基本密度与管胞形态(晚材胞壁率、早材双壁厚度和晚材径向管胞腔径等)及组织比量(除树脂道比量)在无性系之间的差异均达到极显著或显著水平。22个性状中有9个性状的重复力在0.5以上。早材管胞长度与其它材性性状间的正负相关关系基本上和晚材管胞长度与其它性状的正负相关关系一致;基本密度与早材弦向管胞直径、管胞数量间有极显著的负相关,与生长轮宽度、晚材宽度、早材弦向管胞腔径、早材径向管胞直径间有显著的负相关。早材胞壁率与晚材胞壁率间亦呈极显著正相关,晚材率与早、晚材管胞长度间呈极显著正相关。管胞比量与木射线比量间呈极显著正相关关系。采用生长性状与材性性状联合选择的方法筛选出3个综合性状优良的无性系(3号、11号和17号)。
青海云杉;无性系;木材性状;主成分分析;聚类分析
We measured genetic variation in growth and wood traits inPiceacrassifoliaclones and conducted genetic variation analysis, the correlation analysis, principal component analysis and cluster analysis. The superior clones were selected by simple index selection. There were variations within the clones. Latewood had greater coefficient of variation than earlywood, excepting the tracheid length and wall thickness. There were abundant variations in the proportion of resin canal, annual ring width and radial lumen diameter of latewood and the coefficients of variation were in 21.0%-34.2%. There was small variation in the tracheid proportion and basal density and the coefficient of variation was 0.50% and 3.9%, respectively. There were significant differences in basal density and tracheid traits (cell wall percentage of latewood, wall thickness of earlywood, radial lumen diameter of latewood etc.) among clones. There were (very) significant differences in tissue proportion (besiding the proportion of resin canal) among clones. These repeatabilities of nine traits were higher than 0.5. By correlation analysis, the correlation of in earlywood with other wood traits was basically same to that in latewood. Basal density was very negatively correlated with chordwise mid-diameter of earlywood and tracheid number, which was negatively related to annual ring width, latewood width, chordwise lumen diameter and radial mid-diameter of earlywood tracheid. Cell wall percentage of earlywood was very positively correlated with cell wall percentage of latewood, latewood percentage was very positively related to tracheid length of earlywood and latewood, tracheid proportion wood was very positively related to wood ray proportion. Three superior clones were selected by simple index selection.
青海云杉(PiceacrassifoliaKom.)是祁连山东部地区的特有树种,分布于青海、甘肃、宁夏、内蒙等省、区,青海云杉材质优良、抗旱性强,是祁连山、贺兰山和大青山等水源涵养林的主要组成树种[1]。因其单位面积年生长量高、木材品质良好、用途广泛,已成为重要的工业用材树种[2],但目前对其材质的研究甚少[3]。树木的管胞形态特征与木材的用途存在一定的关系,不同用途的木材原料对管胞形态有不同的要求。管胞形态特征与纸浆质量和纸张强度密切相关[4-5],管胞形态受多种环境因子影响,管胞形态特征还存在巨大的遗传变异性,不同树种及同一树种不同种源间、家系间、无性系间均存在明显的变异,即使同一树种同一植株不同部位也有差别[6-7]。因为管胞是衡量木材工艺成熟的重要指标,所以针叶树种的管胞特性、微纤丝角的相关研究已有大量报道[8-11],但是目前对云杉材性的报道并不多,且多集中在管胞及基本密度的径向变异、人工林和天然林材性比较、不同立地条件对管胞形态、基本密度等方面[12-15],而对青海云杉管胞特征的研究罕见报道[16]。无性系利用正成为青海云杉纸浆材经营的方式之一,因此研究不同无性系木材基本密度和管胞形态的变异,对青海云杉新品种的选育和人工林的定向培育具有指导意义。
笔者以31年生青海云杉嫁接无性系种子园为对象,选取其中18个无性系各3个分生株的试材,研究木材基本密度和管胞形态等特征在无性系间的遗传变异规律,并进行主成分分析和聚类分析,探讨青海云杉无性系材质改良的潜力,综合研究青海云杉生长与材性的遗传变异规律和相关关系,并选育出生长和材性兼优的无性系,为青海云杉无性系的早期选择提供参考。
1 试验地概况
试验地设在青海省大通县东峡林场东峡营林区246林班3小班,E101°50′27″,N37°05′42″,海拔2 715 m,年均气温2.9 ℃,最暖月均气温16 ℃,最冷月均气温-11.7 ℃,年均降水量556 mm。
2 材料与方法
2.1 材料
青海云杉初级种子园建于1974年,采用8年生超级苗做砧木,用髓心形成层嫁接法,砧木初植株行距为1 m×3 m,嫁接成活率在95%左右,共嫁接19个无性系。2005年(31年生)取样18个无性系,每个无性系选取3个分生株作标准木,测定每个单株的树高、胸径、冠幅等生长指标,计算各无性系的平均表现。同时利用生长锥在南向胸高处钻取木芯(7 mm),带回实验室进行各材性指标测定。
2.2 测定指标和方法
木材基本密度和管胞长度的测定。木材基本密度的测定采用最大饱和含水量法。管胞长度的测定采用硝酸和铬酸混合液离析的Tayler染色法。经番红液染色,每处理样本在投影显微镜下随机测定50根完整管胞的长度,取其平均值作为样品的管胞长度。
管胞其它形态特征的测定。将软化后的木样,从髓心到树皮方向做斜口连续切片。采用滑走式木材切片机切横面切片,厚度为10~15 μm的斜口式连续生长轮切片,然后分别染色。乙醇梯度脱水,二甲苯透明,施胶封盖玻片。采用日本Nikon 80i研究显微镜、美国Pixera的Penguin 600CL摄像传感器、P4计算机、TDY-5.2彩色图像处理分析软件组成的显微图像检测系统。将制作好的横切面切片,按测定指标要求逐生长轮采集清晰的早、晚材图像,测量取点不低于2处,每个生长轮被测有效管胞数量不低于50个。测定工作由东北林业大学材料科学与工程学院完成。
2.3 数据统计
运用SAS 9.0的Mixed模型进行方差分析,主成分分析、聚类分析以及相关分析,并采用指数选择法进行优良无性系的多性状选择[13]。
3 结果与分析
3.1 无性系间材性性状的变异
如表1所示,管胞比量的变异系数最小,基本密度次之。针叶材管胞长度大于1.8 mm(早晚材管胞长度均大于0.2 mm),管胞总体上分布集中合理,细小管胞含量小。无论是管胞腔径还是管胞直径、无论是径向还是弦向均是早材值较晚材值大,且管胞直径要比管胞腔径大,弦向直径要较径向直径大,其中径向直径的变异系数最大。以上结果均表明管胞表型性状和组织比量在无性系之间的选择潜力较大。此外,如表2所示,重复力大于0.7的性状有基本密度、晚材胞壁率、晚材管胞长度和早材管胞双壁厚度。晚材管胞双壁厚度、晚材径向管胞腔径、晚材径向管胞直径、管胞比量和木射线比量的重复力也都在0.5以上。
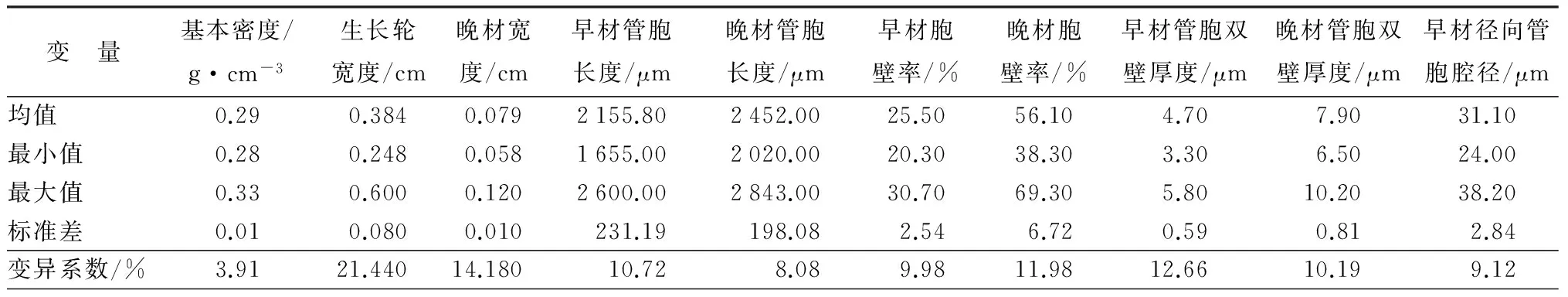
表1 青海云杉无性系间材性性状的遗传变异
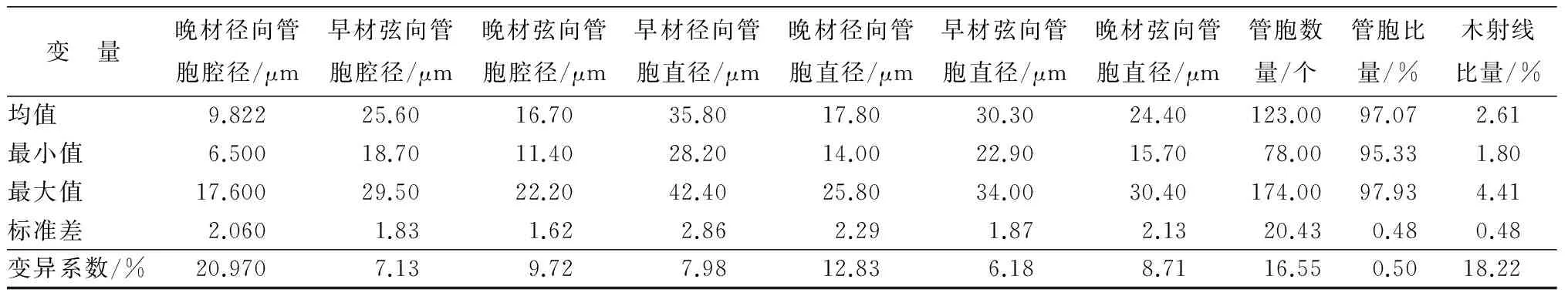
变 量晚材径向管胞腔径/μm早材弦向管胞腔径/μm晚材弦向管胞腔径/μm早材径向管胞直径/μm晚材径向管胞直径/μm早材弦向管胞直径/μm晚材弦向管胞直径/μm管胞数量/个管胞比量/%木射线比量/%均值9.82225.6016.7035.8017.8030.3024.40123.0097.072.61最小值6.50018.7011.4028.2014.0022.9015.7078.0095.331.80最大值17.60029.5022.2042.4025.8034.0030.40174.0097.934.41标准差2.0601.831.622.862.291.872.1320.430.480.48变异系数/%20.9707.139.727.9812.836.188.7116.550.5018.22
方差分析(表2)结果表明:基本密度、晚材胞壁率、早材双壁厚度(相邻2个细胞的壁厚)和晚材径向管胞腔径在无性系间存在极显著差异;晚材宽度、早材胞壁率、晚材双壁厚度和晚材径向管胞直径在无性系间差异显著;管胞比量和木射线比量在无性系间差异极显著。

表2 青海云杉无性系间材性性状的方差分析
3.2 材性性状间的相关分析
由表3可知:基本密度与早材弦向管胞直径、管胞数量间有极显著的负相关,与生长轮宽度、晚材宽度、早材弦向管胞腔径、早材径向管胞直径间显著负相关;生长轮宽度与晚材宽度、早材管胞长度、早材径向管胞腔径、早材弦向管胞腔径、早材径向管胞直径、早材弦向管胞直径及管胞数量有极显著或显著的相关关系。此外,早材管胞长度与晚材管胞长度、晚材率极显著正相关,与早材径向管胞直径(管胞腔径)及管胞数量间呈显著的负相关;晚材管胞长度与晚材率极显著正相关,与管胞数量间极显著负相关,与早材管胞直径(管胞腔径)间显著负相关。
3.3 材性性状的主成分分析
所有的主成分构成中(表4),入选的3个主成分对综合材性的累计贡献率为85%。其中,对第Ⅰ主成分值影响较大的性状有生长轮宽度、晚材宽度、晚材胞壁率和管胞数量;对第Ⅱ主成分值影响较大的性状有晚材宽度、早材管胞长度、早材胞壁率和晚材胞壁率;对第Ⅲ主成分值影响较大的性状有基本密度、生长轮宽度、早材管胞长度、早材胞壁率和管胞数量。

表3 材性性状间的相关关系

表4 材性性状的主成分分析
3.4 基本密度、生长轮宽度等7个性状的聚类分析
本研究利用青海云杉的7个木材性状的遗传相关矩阵计算遗传距离,然后采用系统聚类的类平均法将青海云杉的18个无性系分为3类:第1类包括无性系1、6、9、10、12、14和15;第2类包括无性系3、4和11;第3类包括无性系2、5、7、8、13、16、17和18。
3.5 生长性状优良无性系选择
对青海云杉18个无性系树高和胸径进行初评(表5):树高大于群体平均值的家系从大到小排序是:16、12、11、15、8、3、4、5;胸径大于群体平均值的家系从大到小排序是:8、16、12、11、2、3、4。再根据树高生长量超过群体均值的6.5%和胸径超过群体均值的12%选出了16、12、11号共3个优良无性系。其中16、12、11号无性系树高分别比群体平均值高15.1%、7.4%、6.73%,胸径分别比群体平均值高16.74%、13.49%、13.49%。
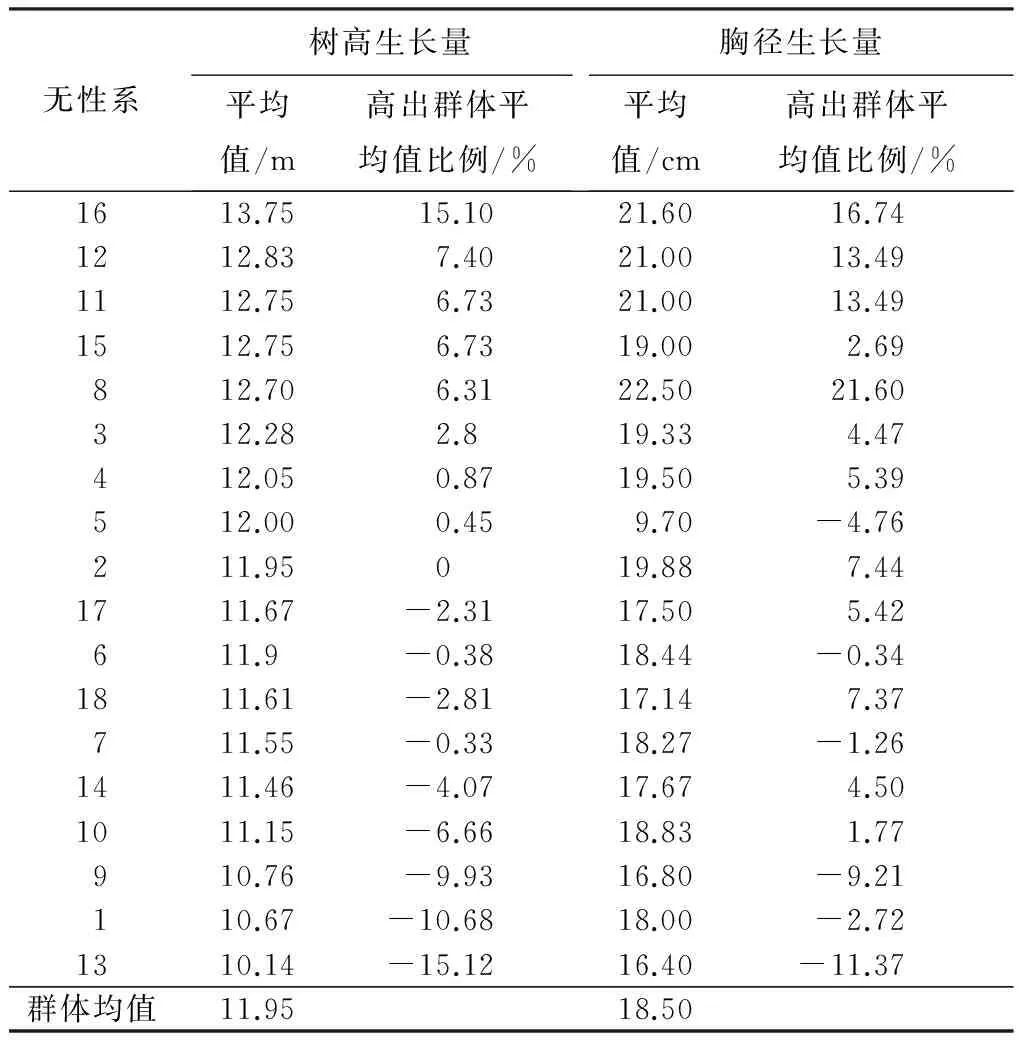
表5 青海云杉无性系树高、胸径生长量统计
3.6 材性性状优良无性系选择
由表6可知,17、18号无性系的基本密度最高,平均基本密度为0.317 g/m3,3、4、7号无性系的基本密度与17、18号无性系间无显著差异;9号无性系的基本密度最低,为0.28 g/m3,较17、18号无性系低13.21%。生长轮宽度、晚材宽度、早材胞壁率、晚材胞壁率、早材管胞双壁厚度、晚材管胞双壁厚度、晚材径向管胞腔径、晚材径向中央直径、管胞比量和木射线比量分别以8、5、10、9、10、3、16、16、12和6号无性系的均值最高,以7、7、16、15、14、5、6、6、6、13号无性系的均值最低。
3.7 生长性状与材性性状间相关分析
如表7所示,树高、胸径、材积3个生长性状是极显著正相关关系。树高、胸径、材积3个生长指标与生长轮宽度和晚材宽度两个主要材性性状指标呈正相关。基本密度和晚材率则与树高、胸径、材积呈负相关关系。
3.8 生长性状和材质性状的联合选择
如表8所示,按15%的入选率和表7中各个无性系指数值的高低,对所有无性系进行综合选择,入选的无性系分别为3、11、17号无性系。

表6 青海云杉无性系材性性状统计
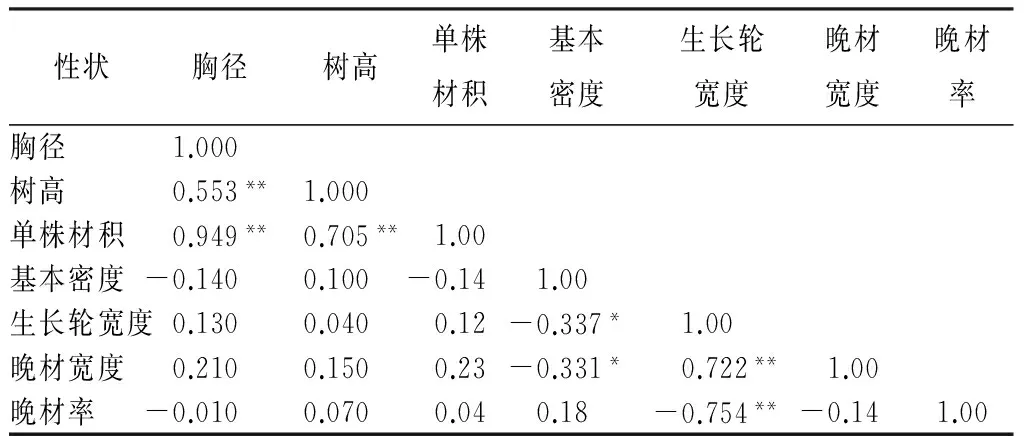
表7 青海云杉无性系生长性状、材性性状间相关分析
4 讨论与结论
本研究发现青海云杉优树无性系间的材性性状具有较大的变异,该结果为青海云杉材性性状的改良提供了可能。基本密度和管胞形态及组织比量在18个无性系间均存在较大的变异,除了早材管胞长度与早材双壁厚度外,其它性状晚材的变异系数均大于早材;树脂道比量、生长轮宽度和晚材径向管胞腔径的变异系数较大(21.0%~34.2%),且基本密度与管胞形态(晚材胞壁率、早材双壁厚度和晚材径向管胞腔径等)及组织比量(除树脂道比量)在无性系之间的差异均达到极显著或显著水平。这说明在无性系水平上开展青海云杉材性因子改良具有很大的潜力。罗建勋等[15]研究发现管胞和木材基本密度在群体间和群体内均存在显著差异。此外,已有研究表明:杨树(Poplusspp.)[17]、泡桐(Pawlowniassp.)[18]、山桂花(Paramicheliabaillonii(Pierre) Hu)[19],日本落叶松(Larixkaempferi(Lamb.) Carr.)和黑杨(PopulusnigraLinn)[20-22]等树种不同无性系间的木材基本密度均存在着极显著差异。本研究表明基本密度和早材管胞长度的重复力均为0.5以上,此结果与前人的研究结果相吻合,如罗建勋等[15]研究发现云杉天然群体的主要材性性状(管胞和基本密度)受较强的遗传控制,王润辉等[23]的研究发现杉木无性系心材率和木材基本密度存在极显著的遗传差异,无性系重复力达中等偏上至较高水平,遗传变异系数各性状间差异较大。马顺兴等[21]研究表明木材基本密度,早、晚材管胞长度等木材性状的重复力均在0.5以上,受中度或中度以上的遗传制约。
表8 青海云杉无性系单株材积、基本密度和管胞长度的平均值和综合选择指数值

无性系单株材积/m3基本密度/g·m-3管胞长度/μm指数10.190.294735.00152.4420.210.294142.33148.2330.190.314961.00161.7240.220.304892.67159.5350.220.294425.33151.2660.200.304668.67155.9270.210.304410.33153.7080.240.294364.33151.8890.190.284600.33147.97100.230.304711.00157.88110.230.304949.33160.33120.170.304704.00154.70130.200.294396.00150.00140.220.304844.33158.58150.200.304643.00155.54160.210.304476.00154.83170.180.324490.33159.57180.170.324511.67158.91
材性性状的相关分析表明,木材基本密度与早材弦向管胞直径、管胞数量间有极显著的负相关,与生长轮宽度、晚材宽度、早材弦向管胞腔径、早材径向管胞直径间有显著的负相关,此结论与王秋玉等[24]对红皮云杉(Piceakoraiensisnakai))的研究结果相一致。王大鹏[25]对日本花柏(Chamaecyparispisifera)的研究也发现木材密度与早、晚材管胞长度、管胞直径、胞壁厚度显著负相关。早、晚材管胞长度与晚材率间呈极显著正相关,而管胞数量与三者极显著或显著负相关,故可以筛选出管胞长度大、管胞数量少、木材强度较高的无性系。而基本密度与早晚材的管胞长度均没有显著相关关系的结果与茹广欣等[3]的研究相吻合。此外,生长轮宽度与基本密度、晚材宽度、早材管胞长度、早材径向管胞腔径、早材弦向管胞腔径、早材径向管胞直径、早材弦向管胞直径及管胞数量有极显著或显著的相关关系。同时,红皮云杉的生长轮宽度与管胞直径及管胞长度也呈显著的正相关[13],故生长轮越宽其管胞直径和管胞长度也越大。
木材密度既影响木材力学强度,又决定木材及纤维制品的产量和品质[26];生长轮是树木生长整个生命过程的反应,生长轮的宽窄可以反应树木的生长速度,表明木材随树龄积累的材积和生长量[27]。此外,笔者只对青海云杉无性系的基本密度、管胞特征及晚材率进行了研究,今后还将进一步开展无性系间纸浆得率等化学性状的相关研究。
早有研究表明中幼云杉人工林、幼龄欧洲云杉(Piceaabies(L.) Karst)木材基本密度与其相关生长性状呈负相关趋势[14,28],这与本研究的基本密度和晚材率与树高、胸径、材积呈负相关关系的结论相一致。可见青海云杉优良无性系选择时,若过分强调生长量的选择,将会导致材质的下降,所以应采取生长与木材材性性状的综合选择较好。此外,由生长性状和材性性状单独选择优良无性系可知,两者选出的优良无性系系号不一致,生长性状指标选出来的3个优良无性系是16、12、11;而这3个无性系的各个材性性状并不是最好。这表明青海云杉无性系生长快的材质不一定最好,材质最好的无性系不一定生长最快。所以要对生长性状、材性性状多性状联合分析来选择生长快、材质优良的无性系[21-23]。
林木育种常用的多性状选择方法有:连续选择法、独立标准法、简单加权法和指数选择法。本研究参考文献[29],用简化的选择指数法,因为指数选择除具有一般选择方法的优点外,还考虑了各个性状的相对经济权重。因此,指数选择法是多性状选择的最佳方法。最终入选的无性系为3、11、17号无性系,这为今后青海云杉的无性系造林推广提供参考。
[1] 刘兴聪.青海云杉[M].兰州:兰州大学出版社,1992:1-16.
[2] 马常耕.世界云杉无性系林业发展现状[J].世界林业研究,1993(6):25-31.
[3] 茹广欣,李林,朱秀红.Pilodyn在青海云杉活立木基本密度预测中的应用[J].河南农业大学学报,2009,43(5):506-510.
[4] Miranda I, Almeida M H, Pereira H. Influence of Provenance, subspecies, and site on wood density inEucalyptusglobuluslabill[J]. Wood and Fiber Science,2001,33(1):9-15.
[5] Miranda I, Almeida M H, Pereira H. Provenance and site variation of wood density inEucalyptusglobulusLabill, at harvest age and its relation to an non-destructive early assessment[J]. Forest Ecology and Management,2001,149(1/3):235-240.
[6] 成俊卿.木材学[M].北京:中国林业出版社,1995.
[7] Pashin A J, de Carl Z. Textbook of Wood Technology[M]. New York: McGraw-HillBook Company,1980.
[8] 白默飞,刘盛全,周亮,等.兴安落叶松管胞形态特征和微纤丝角及其径向变异的研究[J].安徽农业大学学报,2009,36(2):189-193.
[9] 郭明辉,陈广胜,王金满,等.不同土质对人工林落叶松纤维形态及造纸性能的影响[J].东北林业大学学报,2002,30(2):43-45.
[10] 谢新良,石淑兰,魏德津,等.日本落叶松化学组成与纤维特性的研究[J].国际造纸,2004,23(1):24-28.
[11] 陈广胜,郭明辉,黄冶,等.不同初值密度兴安落叶松人工林木材解剖特征的径向变异[J].东北林业大学学报,2001,29(2):12-16.
[12] 徐魁梧,徐永吉,龚士淦.不同立地指数对人工林红皮云杉管胞形态的影响[J].南京林业大学学报,1999,23(1):57-59.
[13] 徐魁梧,潘彪,张耀丽,等.人工林红皮云杉木材管胞形态及其径向变异的研究[J].南京林业大学学报,1997,21(3):39-42.
[14] 四川省云杉纸浆材协作组.云杉人工林材性变异的初步研究[J].西北农林科技大学学报:自然科学版,2001,29(3):29-34.
[15] 罗建勋,李晓清,孙鹏,等.云杉天然群体管胞和木材基本密度性状变异的研究[J].北京林业大学学报,2004,246(6):80-85.
[16] 徐金梅,吕建雄,鲍甫成,等.祁连山青海云杉木材密度对气候变化的响应[J].北京林业大学学报,2011,33(5):115-121.
[17] 王克胜,卞学瑜,佟永昌,等.杨树无性系生长和材性的遗传变异及多性状选择[J].林业科学,1996,32(2):111-117.
[18] 茹广欣,冯胜,袁金玲,等.泡桐无性系木材基本密度遗传变异研究[J].河南农业大学学报,2001,35(4):335-338.
[19] 冯弦,陈宏伟,刘永刚,等.山桂花人工林木材基本密度和纤维长度变异规律的研究[J].广西林业科学,2003,32(1):20-23.
[20] 马顺兴,王军辉,张守攻,等.日本落叶松无性系木材组织比量的遗传变异[J].南京林业大学学报,2006,30(5):55-59.
[21] 马顺兴,王军辉,张守攻,等.日本落叶松无性系木材性质的遗传变异[J].林业科学研究,2008,21(1):69-73.
[22] 沈亚洲,王军辉,张守攻,等.欧洲黑杨生长与材性联合选择[J].西南林业大学学报,2012,32(2):11-15.
[23] 王润辉,胡德活,郑会全,等.杉木无性系生长和材性变异及多性状指数选择[J].林业科学,2012,48(3):45-50.
[24] 王秋玉,李凤娟,任旭琴.红皮云杉纸浆材的初步选择[J].东北林业大学学报,2001,29(5):22-25.
[25] 王大鹏.日本花柏木材管胞解剖特征与基本密度的变异[D].武汉:华中农业大学园艺林学学院,2007:1-65.
[26] 鲍甫成,江泽慧,姜笑梅,等.中国主要人工林树种幼龄材与成熟材及人工林与天然林木材性质比较研究[J].林业科学,1998,34(2):30-36.
[27] 徐有明,徐峰.木材学[M].北京:中国林业出版社,2006.
[28] 梁凤山,赵悦,孟凡忠,等.云衫遗传多样性研究进展[J].林业科技,2000,25(3):8-10.
[29] 韦力权.杉木无性系生长与材性综合遗传改良研究[D].南宁:广西大学林学院,2004:1-62.
Wood Traits ofPiceacrassifoliaClones//
Li Qingfen, Wang Junhui
(Key Laboratory of Tree Breeding and Cultivation of State Forestry Administration Institute of Forestry, Chinese Academy of Forestry, Beijing 100091, P. R. China); Li Dengping, Hu Changshou, Qi Shengxiu(Forestry Center of Dongxia, Datong County, Qinghai Province)//Journal of Northeast Forestry University,2015,43(12):12-16,35.
Piceacrassifolia; Clones; Wood traits; Principal component analysis; Cluster analysis
李青粉,女,1987年2月生,国家林业局林木培育重点实验室(中国林业科学研究院林业研究所),博士研究生;现工作于广西壮族自治区林业科学研究院经济林研究所,工程师。E-mail:li63757416@163.com。
王军辉,国家林业局林木培育重点实验室(中国林业科学研究院林业研究所),研究员。E-mail:wangjh808@sina.cn。
2013年10月17日。
S791.18;S722.3+3
1)“十二五”国家科技支撑计划课题(2012BAD01B01)。
责任编辑:戴芳天。
