长江口及邻近海域浮游水螅水母、管水母和栉水母的丰度分布与季节变化
2015-02-05陈佳杰徐兆礼朱德弟
高 倩, 陈佳杰,徐兆礼,*,朱德弟
1 中国水产科学研究院东海水产研究所 农业部海洋与河口渔业重点开放实验室, 上海 200090 2 国家海洋局第二海洋研究所, 杭州 310012
长江口及邻近海域浮游水螅水母、管水母和栉水母的丰度分布与季节变化
高 倩1, 陈佳杰1,徐兆礼1,*,朱德弟2
1 中国水产科学研究院东海水产研究所 农业部海洋与河口渔业重点开放实验室, 上海 200090 2 国家海洋局第二海洋研究所, 杭州 310012
依据2002—2003年长江口及邻近海域(29°00′—32° 00′ N、122° 00 —123° 30′ E)4个季节的海洋调查资料,探讨了该水域小型水母(含水螅水母、管水母和栉水母)总丰度的空间分布特征及季节差异。结果显示,该海域共出现小型水母41种,其中水螅水母28种,管水螅11种,栉水母2种。主要优势种包括管水母大西洋五角水母(Muggiaeaatlantica)、双生水母(Diphyeschamissonis)和气囊水母(Physophorahydrostatica)以及水螅水母四叶小舌水母(Liriopetetraphylla)、两手筐水母(Solmundellabitentaculata)和短柄和平水母(Eirenebrevistylus),其中大西洋五角水母为冬、春季第一优势种,双生水母则为夏、秋季第一优势种。该水域小型水母总丰度呈明显季节差异,夏季丰度最高((337.33±55.68) 个/m2),春、秋季次之(分别为(142.09±67.71) 个/m2和(132.84±35.17) 个/m2),冬季最低((113.69±32.72)个/m2)。水温是影响调查区小型水母总丰度季节变化的主要环境因子,小型水母丰度与表层温度显著正相关(P< 0.01)。盐度是影响小型水母平面分布格局的主要环境因子,春季小型水母丰度与底层盐度正相关(P< 0.05),夏季与10 m层盐度正相关(P< 0.01),全年则是与10 m层盐度正相关(P< 0.01)。此外,调查水域小型水母丰度与其饵料——小型桡足类亦存在显著正相关关系(P< 0.01)。
水螅水母;管水母;栉水母;季节分布;长江口
刺胞动物门的水螅水母和管水母以及栉水母动物门的栉水母,统称为小型水母,是海洋浮游动物的重要类群[1- 2]。小型水母作为海洋浮游动物最重要的肉食性类群,具有种类多、分布广、数量大的特点,是海洋生态系统物质循环和能量流动过程中一个重要环节[2- 4]。小型水母能大量摄食鱼卵、仔稚鱼,可对渔业资源早期补充过程产生负面影响[5- 6]。近年来,随着人们对其生态功能认识的逐渐加深[5- 8],小型水母时空格局及其驱动机制的研究逐步成为海洋生态学中的热点[9- 11]。
长江口及邻近水域作为我国重要的高生产力海区之一,是众多经济鱼类的产卵场、索饵场和育幼场。长江口及邻近水域受到长江冲淡水和台湾暖流的直接影响,并受到苏北沿岸水、闽浙沿岸水、黄海冷水团和黑潮水的间接影响,水动力环境复杂[12],从而形成了多样化生境,长期以来一直是我国海洋生态学研究的热点水域。迄今为止,该海区小型水母研究有限,多为物种组成和数量分布规律的研究[13- 16],缺乏对其时空分布规律及其与环境因子关系的定量分析。鉴于此,本研究利用2002—2003年4个航次的海洋大面调查资料,在描述长江口及邻近水域的小型水母空间分布特点和季节演替规律的基础上,利用多元回归分析,探讨环境因子对小型水母时空变动的影响机制。研究结果不仅对长江口及邻近水域小型水母生态学研究提供基础资料,亦将丰富人们对河口区小型水母时空变动规律的理论认知。

图1 长江口及邻近水域调查站位(2002-04—2003-03)[17] Fig.1 Sampling stations in the Changjiang Estuary and the adjacent East China Sea during April 2002—March 2003[17]
1 材料与方法
1.1 调查区域、采样时间和方法
分别于2002年4—5月、8—9月、11月和2003年2—3月在长江口及邻近水域(29°00′— 32°00′N、122°00′—123°30′E)进行了4个航次的水文和生物综合调查,涉及浮游动物的站位布设如图1所示[17]。
浮游动物样品采集和室内处理均按照《海洋调查规范》进行,小型水母用大型浮游生物网(口径80 cm、网目孔径0.505 mm)由底至表层垂直拖曳采集获得;小型桡足类作为小型水母的重要饵料,使用中型浮游生物网(口径50 cm、网目孔径0.160 mm)采集。所获样品经5%福尔马林溶液固定后,在体视显微镜下,按个体计数法对浮游动物样品中的水母鉴定到种[1,18- 21],丰度以个/m2表示;小型桡足类鉴定到大类,丰度以个/m3表示。现场温度、盐度、水深等数据采用SBE37-CTD仪测得。
1.2 数据处理
1.2.1 优势种
本文中小型水母优势种通过以下公式
确定:
(1)
式中,ni为第i种的丰度;fi是该种的出现频率;N为水母类的总丰度。取优势度Y≥0.02的物种为优势种[22]。
1.2.2 逐步回归分析
以水温(℃), 表层(t0) 、10m层(t10m) 、底层(tb) 和盐度, 表层(S0) 、10m层(S10m) 、底层(Sb) 6个因子作自变量,以二次方根转化后的小型水母丰度(A)为因变量,采用逐步回归分析方法,探究环境因子对小型水母丰度变化的影响[23]。
2 结果
2.1 种类组成
调查水域共出现小型水母41种,隶属于2门4(亚)纲7目,其中水螅水母最多,达28种;管水母次之(11种);栉水母最少,仅2种。5月,共出现小型水母15种,其中管水母7种,水螅水母6种;其他月份均以水螅水母物种数最多,均占小型水母总物种数的50%以上。
2.2 小型水母总丰度的空间分布及季节变化
尹爱群手忙脚乱地删掉妻子手机上的图片,又删除了Q Q的聊天记录,拿起提包匆匆下楼,把车在街区里转了几个弯后,确定没有尾巴,才把款存到一家银行里。之后,他把银行卡放到一个小手包里,鬼鬼祟祟地到了扬扬那里,扬扬温柔地叫着老公,刚要撒娇,见其面色难看,怯怯地张了张嘴。
如表1所示,调查水域小型水母总丰度夏季最高((337.33 ±55.68)个/m2),春、秋季次之(分别是(142.09±67.71)个/m2和(132.84±35.17)个/m2),冬季最低((113.69±32.72)个/m2)。管水母构成其数量主体,以春季为例,管水母丰度为(129.37±66.64) 个/m2,占小型水母总丰度的90%以上(图2a和图2i)。
春季,小型水母丰度 > 500 个/m2区域主要位于该水域东南部,其高丰度中心达1379 个/m2,主要由管水母大西洋五角水母(Muggiaeaatlantica)构成(图2a和图2i)。夏季,调查水域有两个高丰度区域(最大丰度 > 750 个/m2), 其一位于中部海域 (127.75—123.5°E, 30.1—30.75°N),管水母大西洋五角水母和双生水母(Diphyeschamissonis)构成其主体; 另一高丰度区东南海域(123.0—123.5°E, 28.8—29.2°N), 最高值为1032.8 个/m2, 主要由双生水母构成(图2b和图2j)。秋季,小型水母出现率为100%,为四季最高。高丰度区,主要由双生水母构成(图2c和图2k)。冬季,小型水母平均丰度和出现率(71.43%)均为全年最低。高丰度区(> 500 个/m2)位于123.5°E断面的29.5—31.0°N海域,主要由大西洋五角水母、水螅水母两手筐水母(Solmundellabitentaculata)和四叶小舌水母(Liriopetetraphylla)等构成(图2d)。

表1 长江口及邻近水域海水温度 和小型水母丰度的季节变化Table 1 Seasonal variation in sea water temperature and abundance of small jellyfish in the Changjiang Estuary and adjacent East China Sea
2.3 小型水母优势种及其季节变化特征
如表2所示,调查水域共出现6种优势种。大西洋五角水母为冬、春季的第一优势种,其丰度分别为44.47 个/m2和81.74 个/m2;双生水母则为夏、秋季第一优势种,平均丰度分别为249.03 个/m2和98.04 个/m2。
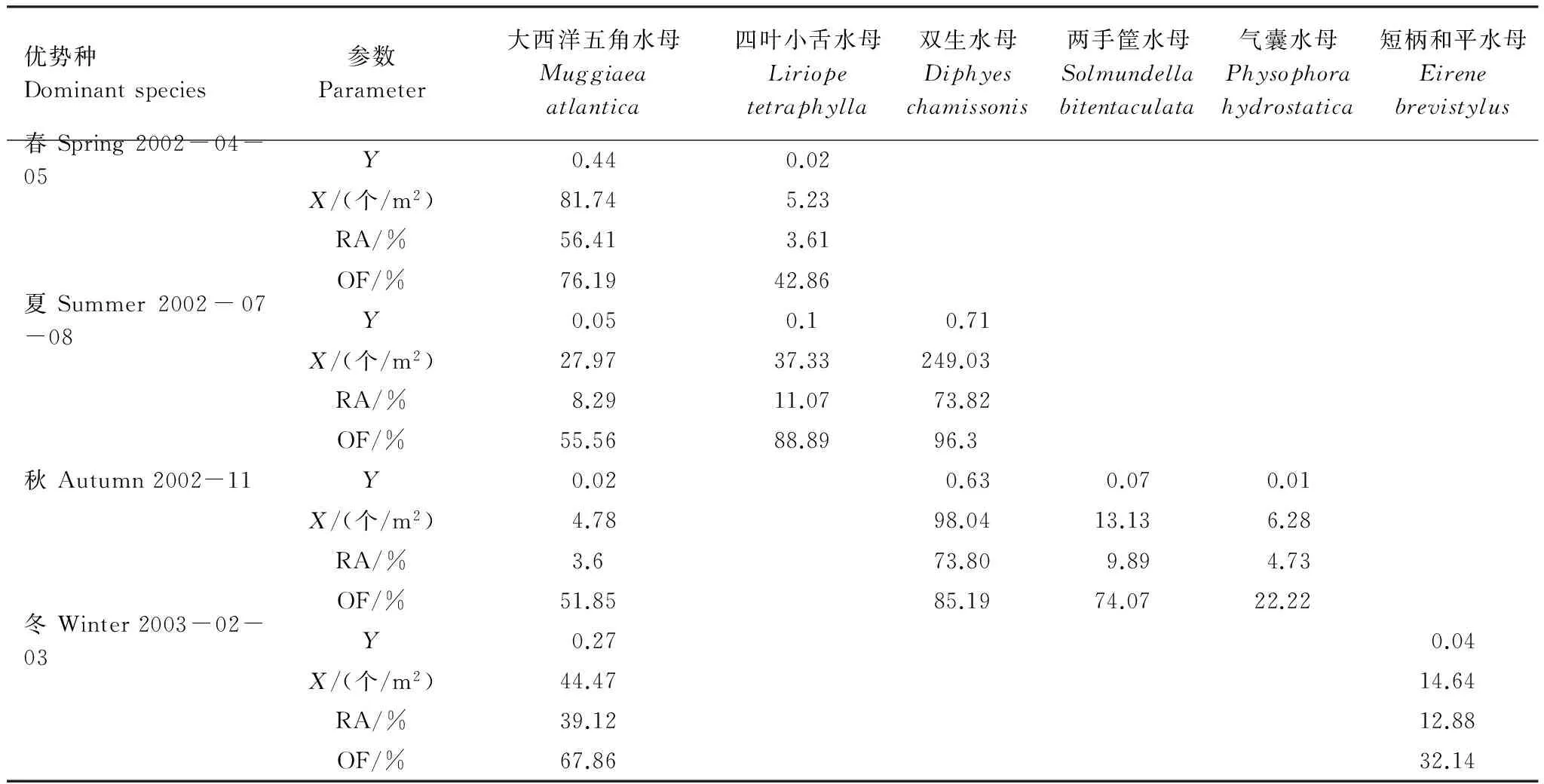
表2 长江口及邻近水域小型水母优势种Table 2 Dominant species of small jellyfish in the Changjiang Estuary and the adjacent East China Sea

图2 长江口及邻近水域小型水母各类群丰度空间分布Fig.2 Spatial distribution of different groups of small jellyfish in the Changjiang Estuary and adjacent East China Sea 小型水母(a. 春、b. 夏、c. 秋、d. 冬),水螅水母(e. 春、f. 夏、g. 秋、h. 冬)、管水母(i. 春、j. 夏、k. 秋、l. 冬)和栉水母(m. 春、n. 夏、o. 秋)
2.4 小型水母主要优势种的温、盐适应特征
小型水母主要分布在表、中层水体[24],因此,本研究选择10 m层温度和盐度研究优势种的环境适应特征。大西洋五角水母和双生水母是调查水域最重要优势种,其温、盐度适应特征如图3所示。双生水母为亚热带广盐种,当温度大于18 ℃,其丰度和出现频率明显增加;当温度为25—27 ℃时,该种广泛分布于各水域,且丰度较高。与双生水母相比,大西洋五角水母则适盐较高,其高丰度主要分布于盐度>30的水域,出现率为70%;而当盐度 < 30时,其丰度较低,出现率降至45.24%。

图3 大西洋五角水母和双生水母的温度、盐度适应特征Fig.3 Temperature and salinity adaption of Muggiaea altantia, and Diphyes chamissonis
2.5 温、盐度和小型桡足类对小型水母丰度变动的影响
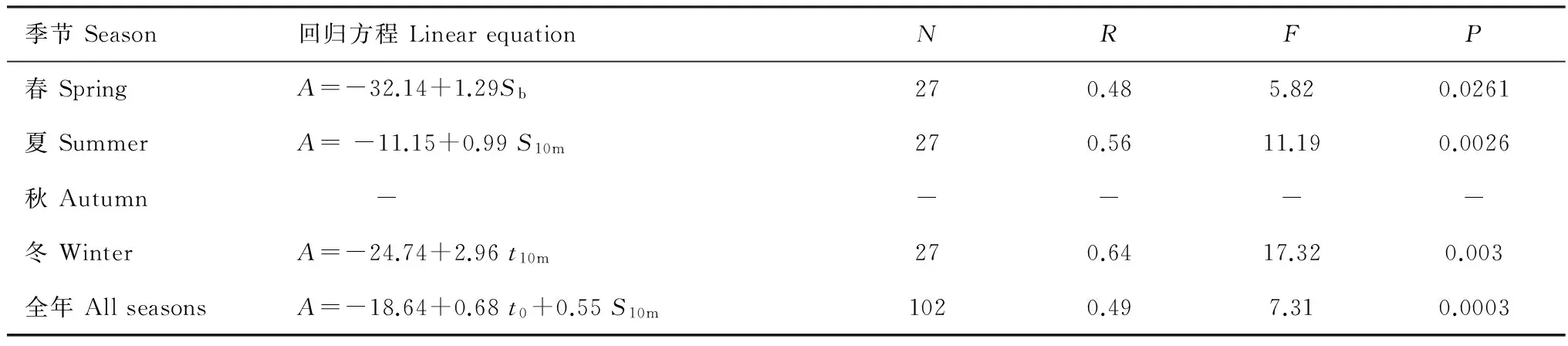
表3 小型水母总丰度与温、盐度的回归分析Table 3 Linear regression between sea water temperature, sea water salinity and abundance of small jellyfish
3 讨 论
3.1 环境因子对小型水母分布的影响
小型水母营浮游生活,对环境较为敏感,其生长、繁殖和分布与其所处水团的性质(例如温、盐度)密切相关[25]。研究表明,长江口及邻近海域小型水母丰度呈现明显季节性差异,具体表现为夏高冬低。该分布特点与其他中高纬度海区小型水母季节分布特点相吻合[26- 28]。逐步分析结果表明,该分布特点与该水域季节性温度差异密切相关(表3)。夏季水温达全年最高值(10 m层平均水温为26.1 ℃),小型水母丰度亦达全年最高337.33 个/m2(表1和表2)。夏季为高温适应的小型水母提供了适宜的生长和繁殖条件。以双生水母为例,该种广泛分布于东海、南海和泰国湾等水域,是南海北部、东海外海和闽江口等亚热带水域的优势种[29]。当水温 > 18 ℃时,暖水种双生水母丰度和出现频率则大幅增加(图3)。夏季高温适应性的双生水母丰度急剧增加,平均丰度高达249.03 个/m2(占小型水母总丰度70%)(图3)。秋季,调查海域浮游动物群落虽仍呈现亚热带特征,然而伴随着水温的降低((18.86±0.06)℃),暖水种双生水母的数量大幅降低(表2,图3)。冬、春季,适温较低的暖温种大西洋五角水母为最主要的优势种。春季(温度为17.5 ℃),大西洋五角水母丰度达全年最大(81.74 个/m2)。冬季水温较低(表温:(10.36±0.08)℃),不利于大西洋五角水母的生长和繁殖(图3)。大西洋五角水母丰度为44.47 个/m2,仅大约为春季数量的1/2。
食物也是夏季小型水母丰度较高的重要原因。在挪威的Korsfjorden湾,适宜温度以及食物导致锥体浅室水母(Lensiaconoidea)和北极单板水母(Dimophyesarctica)丰度在5—6月(春、夏季)达全年最高[30]。本研究中,夏季调查海域叶绿素a和小型桡足类均达到全年最大值(表1)。小型桡足类如小拟哲水蚤等是小型水母的重要饵料[31-32]。小型桡足类旺发,为小型水母提供了丰富的饵料。小型水母丰度与小型桡足类显著正相关,进一步证实了饵料的重要性。小型水母,特别是营无性生殖且终生浮游的管水母,对食物响应较快,生长和繁殖率变化快[33- 34]。夏季,双生水母数量激增至249.03 个/m2, 成为夏季第一优势种(表2)。四叶小舌水母丰度增加至37.33 个/m2,约为春季的6倍(表2)。栉水母如球型侧腕水母也在夏季达到全年最大值。总之,夏季大量饵料进一步促进了小型水母数量的丰富。
除了食物和温度,盐度也是调查海域小型水母丰度水平分布的主要因子。盐度变化可影响小型水母的浮力、繁殖和摄食率[35-36]。小型水母种群对盐度变化较为敏感,高丰度值一般是在其最适盐度附近[37-39]。本研究中长江口及邻近水域小型水母全年分布与10 m层盐度显著正相关 (P=0.003),小型水母高丰度区主要位于高盐水域(表3)。调查海域小型水母组成以适盐较高的外海种为主,仅广盐种双生水母为近海种。以春季为例,优势种管水母大西洋五角水母和水螅水母四叶小舌水母均适盐较高 (图2a,表2),为外海种[39]。它们主要分布在盐度大于30的水域,当盐度低于30时候,出现频率和丰度均显著降低(图3)。因此,春季小型水母丰度与底层盐度显著正相关,其丰度在东南部水域达到极大值,西北方向上呈现舌状分布(表3,图2a)。总之,盐度是决定小型水母优势种分布及其丰度的重要因素。
3.2 小型水母空间分布与水团的关系
在大中尺度研究中,小型水母分布与物理海洋过程例如洋流、锋面、水团混合、上升流等密切相关[28, 40- 41]。长江口及邻近水域受长江冲淡水、台湾暖流和沿岸流等影响,水文动力环境复杂多变[42- 43],对小型水母空间分布可产生重要影响。
冬季,受偏北风影响,长江冲淡水沿岸南下,其范围仅限于贴岸的一狭带内[42]。长江径流南下向杭州湾口、舟山群岛海域方向扩展,该区域几乎无小型水母个体出现(图2d)。此时,长江口外海存在一个总体上呈南北方向分布的温度锋和盐度锋,低温、低盐的长江冲淡水和东海沿岸流沿着东海沿岸向南流,而高温、高盐台湾暖流从底层向北入侵,垂直分布均匀[42]。此时,小型水母主要分布在冷暖流交汇偏暖水侧。冬季小型水母丰度与10 m层温度显著正相关(表3),进一步证实了上述推测。
春季,小型水母主要分布在长江口东南部水域—台湾暖流与长江冲淡水交汇偏暖流一侧。台湾暖流为靠近浙江、福建近海终年向北流动的一支高温、高盐的海流, 其内侧分支在舟山群岛以东、长江口以南转向东[12]。由于台湾暖流从东南部接近长江口水域,小型水母则随着暖流进展,遇到长江口冲淡水后在长江口南侧30°00′N,123°30′E 处聚集(图2a和图2i)。
夏季,长江口及邻近水域夏季存在稳定的浙江沿岸上升流和长江口上升流[44]。研究表明,上升流与小型水母繁盛密切相关。例如,在太平洋东部热带水域的巴拿马湾,上升流及其由此引发的生产力增加与水螅水母爆发密切相关[45]。南海北部海域上升流增强驱动产生的丰富食物,是双生水母等管水母夏季出现高峰的重要原因[46]。本研究中,上升流为真光层补充了大量无机磷,有利于浮游植物的生长[47],并促进了小型桡足类和小型水母的旺发。以往研究也证实了夏季沿岸上升流对长江口及邻近水域浮游植物繁盛的作用[48]。由此推测,夏季,长江口及邻近海域小型水母分布广泛且丰度最大,亦与上升流及其引发的高生产力密切相关。而秋季,水团与小型水母丰度分布关系不甚紧密(图3,表3)。
从以上分析来看,冬、春季小型水母高丰度主要分布于长江冲淡水和台湾暖流交汇偏暖水侧;夏季小型水母分布广泛且丰度最高,与上升流及其引发的高生产力密切相关。
3.3 与外海小型水母的比较
纵观整个东海小型水母分布,长江口及邻近水域位于丰度较高的水域,其小型水母丰度明显高于东海北部近海和外海[49]。这是由于在长江口及邻近水域存在生物生产力锋区,随着冲淡水向外海方向扩散,水体层化,垂直稳定度大,有利于悬浮泥沙迅速沉降,致使可利用率大大增加,在河口输入的营养盐,可满足浮游植物群落快速增长[50]。而生物生产力锋区浮游植物旺发,促进了小型浮游桡足类生长,最终为小型水母提供了丰富的饵料。此外,上升流亦与小型水母繁盛密切相关。无论东海外海[49],还是长江口水域,最重要的优势种都是大西洋五角水母和双生水母,显示上述两个物种在东海小型水母丰度变动特征中重要性。
致谢:感谢林茂研究员对样品鉴定的指导,沈晓民老师对写作的帮助。
[1] 高尚武, 洪惠馨, 张士美. 中国动物志 无脊椎动物 第二十七卷 刺胞动物门 水螅虫纲管水母亚纲 钵水母纲. 北京: 科学出版社, 2002.
[2] Richardson A J, Bakun A, Hays G C, Gibbons M J. The jellyfish joyride: causes, consequences and management responses to a more gelatinous future. Trends in Ecology and Evolution, 2009, 24(6): 312- 322.
[3] Lynam C P, Hay S J, Brierley A S. Jellyfish abundance and climatic variation: contrasting responses in oceanographically distinct regions of the North Sea, and possible implications for fisheries. Journal of the Marine Biological Association of the United Kingdom, 2005, 85: 435- 450.
[4] Purcell J E, Uye S I, Lo W T. Anthropogenic causes of jellyfish blooms and their direct consequences for humans: a review. Marine Ecology Progress Series, 2005, 350: 153- 174.
[5] Purcell J E, Sturdevant M V. Prey selection and dietary overlap among zooplanktivorous jellyfish and juvenile fishes in Prince William Sound, Alaska. Marine Ecology Progress Series, 2001, 210: 67- 83.
[6] Purcell J E, Arai M N. Interactions of pelagic cnidarians and ctenophores with fish: a review. Hydrobiologia, 2001, 451(1- 3): 27- 44.
[7] Kideys A E. Fall and rise of the Black Sea ecosystem. Science, 2002, 297(5586): 1482- 1484.
[8] Sullivan B K, Keuren D V, Clancy M. Timing and size of blooms of the ctenophoreMnemiopsisleidyiin relation to temperature in Narragansett Bay, RI. Hydrobiologia, 2001, 451(1- 3): 113- 120.
[9] Lynam C P, Heath M R, Hay S J, Brierley A S. Evidence for impacts by jellyfish on North Sea herring recruitment. Marine Ecology Progress Series, 2005, 298: 157- 167.
[10] Costello J H, Sullivan B K, Gifford D J. A physical-biological interaction underlying variable phenological responses to climate change by coastal zooplankton. Journal of Plankton Research, 2006, 28(11): 1099- 1105.
[11] Attrill M J, Wright J, Edwards M. Climate-related increases in jellyfish frequency suggest a more gelatinous future for the North Sea. Limnology and Oceanography, 2007, 52(1): 480- 485.
[12] Su J L. Circulation dynamics of the China Seas: north of 18°N // Robinson A R, Brink K. The Sea. Vol. 11. The Global Coastal Ocean: Regional Studies and Syntheses. New York: Wiley, 1998: 483- 506.
[13] 高哲生, 张志南. 舟山的水螅水母类. 山东海洋学院学报, 1962, (1): 65- 91.
[14] 洪惠馨. 东海水母类的研究Ⅰ. 浙江沿海的管水母类 (Siphonophora). 上海水产学院论文集, 1964, (1): 111- 130.
[15] 许振祖, 黄加祺, 刘光兴. 长江口及其邻近海域水螅水母纲新种和新记录记述. 海洋学报, 2006, 28(6): 112- 118.
[16] 陈洪举, 刘光兴. 夏季长江口及邻近海域水母类生态特征研究. 海洋科学, 2010, 34(4): 17- 24.
[17] 周伟华, 袁翔城, 霍文毅, 殷克东. 长江口邻域叶绿素a和初级生产力的分布. 海洋学报, 2004, 26(3): 143-150.
[18] 周太玄, 黄明显. 烟台水螅水母类的研究. 动物学报, 1958, 10(2): 173- 191, 图版I-V.
[19] 许振祖, 金德祥. 福建沿海水母类的调查研究(一). 厦门大学学报, 1962, 9(3): 206- 224.
[20] Kramp P L. The Hydromedusae of the Pacific and Indian Oceans. Sections II and III. Dana-Report, 1968, 72: 1- 200.
[21] 张金标. 中国海洋浮游管水母类. 北京: 海洋出版社, 2005.
[22] 徐兆礼, 陈亚瞿. 东黄海秋季浮游动物优势种聚集强度与鲐鲹渔场的关系. 生态学杂志, 1989, 8(4): 13- 15.
[23] 李春喜, 姜丽娜, 邵云. 生物统计学. 北京: 科学出版社, 2008.
[24] Gao S W. The vertical distribution of the Medusae, Pteropoda, Heteropoda and Thaliacea in the East China Sea. Studia Marina Sinica, 1990, 31: 83- 91.
[25] Molinero J C, Casini M, Buecher E. The influence of the Atlantic and regional climate variability on the long-term changes in gelatinous carnivore populations in the northwestern Mediterranean. Limnology and Oceanography, 2008, 53(4): 1456- 1467.
[26] Gibbons M J, Richardson A J. Patterns of jellyfish abundance in the North Atlantic. Hydrobiologia, 2009, 616: 51- 65.
[27] Fraser J H. The ecology of the ctenophorePleurobrachiapileusin Scottish waters. Journal du Conseil International pour l′Exploration de la Mer, 1970, 33(2): 149- 168.
[28] Brinckmann-Voss A. Seasonality of hydroids (Hydrozoa, Cnidaria) from an intertidal pool and adjacent subtidal habitats at Race Rocks, off Vancouver Island, Canada. Scientia Marina, 1996, 66: 89- 97.
[29] Alvarino A. Siphonophores of the Pacific with A Review of the World Distribution. Bulletin of the Scripps Institution of Oceanography, 1971, 16: 1- 432.
[30] Hosia A, Båmstedt U. Seasonal abundance and vertical distribution of siphonophores in western Norwegian fjords. Journal of Plankton Research, 2008, 30(8): 951- 962.
[31] Purcell J E. Dietary composition and diel feeding patterns of epipelagic Siphonophores. Marine Biology, 1981, 65(1): 83- 90.
[32] Purcell J E. Feeding and growth of the siphonophoreMuggiaeaatlantica(Cunningham 1893). Journal of Experimental Marine Biology and Ecology, 1982, 62(1): 39- 54.
[33] Mackie G O, Pugh P R, Purcell J E. Siphonophore biology. Advances in Marine Biology, 1987, 24: 98- 262.
[34] Clarke A, Peck L S. The physiology of polar marine zooplankton. Polar Research, 1991, 10(2): 355- 369.
[35] Mills C E. Density is altered in hydromedusae and ctenophores in response to changes in salinity. Biological Bulletin, 1984, 166(1): 206- 215.
[36] Ma X P, Purcell J E. Temperature, salinity, and prey effects on polyp versus medusa bud production by the invasive hydrozoanMoerisialyonsi. Marine Biology, 2005, 147(1): 225- 234.
[37] Licandro P, Souissi S, Ibanez F, Carré C. Long-term variability and environmental preferences of calycophoran siphonophores in the Bay of Villefranche (north-western Mediterranean). Progress in Oceanography, 2012, 97- 100: 152- 163.
[38] Lo W T, Yu S F, Hsieh H Y. Effects of summer mesoscale hydrographic features on epipelagic siphonophore assemblages in the surrounding waters of Taiwan, western North Pacific Ocean. Journal of Oceanography, 2013, 69(5): 495- 509.
[39] 徐兆礼. 东海水螅水母环境适应与生态类群. 应用生态学报, 2009, 20(1): 177- 184.
[40] Thibault-Botha D, Lutjeharms J R E, Gibbons M. Siphonophore assemblages along the east coast of South Africa; mesoscale distribution and temporal variations. Journal of Plankton Research, 2004, 26(9): 1115- 1128.
[41] 林茂, 张金标. 东海中部管水母类和海樽类的密集及其对水团边界的指示作用. 国家海洋局科技司, 黑潮调查研究论文选(一). 北京: 海洋出版社, 1993, 5: 452- 459.
[42] 苏育嵩. 黄、东海地理环境与环流系统分析—长江口及济州岛邻近海域综合调查研究报告(第二章). 青岛海洋大学学报, 1989, 19(1): 145- 158.
[43] Li G X, Han X B, Yue S H, Wen G Y, Yang R M, Kusky T M. Monthly variations of water masses in the East China Seas. Continental Shelf Research, 2006, 26(16): 1954- 1970.
[44] 赵保仁, 任广法, 曹德明, 杨玉玲. 长江口上升流海区的生态环境特征. 海洋与湖沼, 2001, 32(3): 327- 333.
[45] Miglietta M P, Rossi M, Collin R. Hydromedusa blooms and upwelling events in the Bay of Panama, Tropical East Pacific. Journal of Plankton Research, 2008, 30(7): 783- 793.
[46] Li K Z, Yin J Q, Huang L M, Song X Y. Comparison of siphonophore distributions during the southwest and northeast monsoons on the northwest continental shelf of the South China Sea. Journal of Plankton Research, 2012, 34(7): 636- 641.
[47] 王保栋. 长江口及邻近海域富营养化状况及其生态效应 [D]. 青岛: 中国海洋大学, 2006.
[48] Chen Y L L, Chen H Y, Gong G C, Lin Y H, Jan S, Takahashi M. Phytoplankton production during a summer coastal upwelling in the East China Sea. Continental Shelf Research, 2004, 24(12): 1321- 1338.
[49] 徐兆礼. 东海水母类丰度的动力学特征. 动物学报, 2006, 52(5): 854- 861.
[50] 宁修仁, 史君贤, 蔡昱明, 刘诚刚. 长江口和杭州湾海域生物生产力锋面及其生态学效应. 海洋学报, 2004, 26(6): 96- 106.
Abundance distribution and seasonal variation of medusae, siphonophores, and ctenophores in the Changjiang (Yangtze River) Estuary and the adjacent East China Sea
GAO Qian1, CHEN Jiajie1, XU Zhaoli1,*, ZHU Dedi2
1KeyandOpenLaboratoryofMarineandEstuarineFisheries,MinistryofAgriculture,EastChinaSeaFisheriesResearchInstitute,ChineseAcademyofFisherySciences,Shanghai200090,China2SecondInstituteofOceanography,SOA,Hangzhou310012,China
Small jellyfish (here including medusae, siphonophores, and ctenophores) are widespread in the marine pelagic realm. They usually constitute one of the most important assemblages of predators in pelagic food webs by serving as a direct link between zooplankton and higher trophic levels. They can at times be the most abundant non-crustacean invertebrate predators due to their great voracity and their rapid population growth, resulting in extensive seasonal blooms. Thus, studies on small jellyfish have become popular in marine ecology. Studies concerning zooplankton distribution at large spatial scales, particularly for weak swimmers like small jellyfish, emphasize the importance of the physical characteristics of water masses, including salinity and temperature. In our study, the seasonal abundance and horizontal distribution of small jellyfish were assessed in the Changjiang Estuary (Yangtze River) and the adjacent East China Sea (29°00′—32°00′N, 122°00′—123°30′E) in 2002—2003. The Changjiang Estuary and the adjacent East China Sea are one of the most productive aquatic ecosystems. High primary production supports high fishery production in adjacent coastal waters, including the Changjiang and Zhoushan fishery grounds. A total of 41 species of small jellyfish (11 siphonophores, 28 medusae, 2 ctenophores) were collected. Numerically, siphonophores were most abundant (58.56%—91.05% of the total collection by number). The dominant species were the siphonophoresDiphyeschamissonis,Muggiaeaatlantica, andPhysophorahydrostaticaand medusaeLiriopetetraphylla,Solmundellabitentaculata, andEirenebrevistylus.Muggiaeaatlanticawas dominant all year round, particularly in spring (76.19% of the total collection by number) and winter (39.12%).D.chamissoniswas dominant in summer and autumn, with abundances of 249.03 ind./m2(73.82% of the total collection by number), and 98.04 ind./m2(73.80%), respectively. The numerical abundance of small jellyfish was highest in summer ((337.33±55.68) ind./m2), lowest in winter ((113.69±32.72) ind./m2), and intermediate in spring and autumn ((142.09±67.71) ind./m2and (132.84±35.17) ind./m2). Seasonal variations in the abundance of small jellyfish were closely associated with the surface water temperature. The average surface water temperature peaked in summer ((27.26±0.02) ℃). Then, both the temperature and salinity in the studied area were suitable for the development and reproduction ofD.chamissonis. In addition, the summer upwelling and its associated high productivity might also benefit the jellyfish because large numbers of carnivores ultimately require an elevated level of primary production. There were strong correlations between temperature, 10-m water salinity, and small jellyfish abundances (P< 0.01), suggesting that they are important in determining the spatial distribution of small jellyfish. The bottom salinity in spring was positively related with the abundance (P< 0.05), and the 10-m water salinity in summer was also positively correlated with abundance (P< 0.01). In winter, the abundance was obviously increased from inshore to offshore. In addition, small jellyfish abundance was also strongly correlated with their food, small copepods (P< 0.01), which proved the importance of food availability for carnivores. This study provides basic knowledge on the distribution patterns of small jellyfish, which is essential to further understand the ecological roles played by small jellyfish and their responses to the hydrological conditions in the Changjiang Estuary and the adjacent East China Sea.
medusae; siphonophores; ctenophores; seasonal distribution; Changjiang Estuary
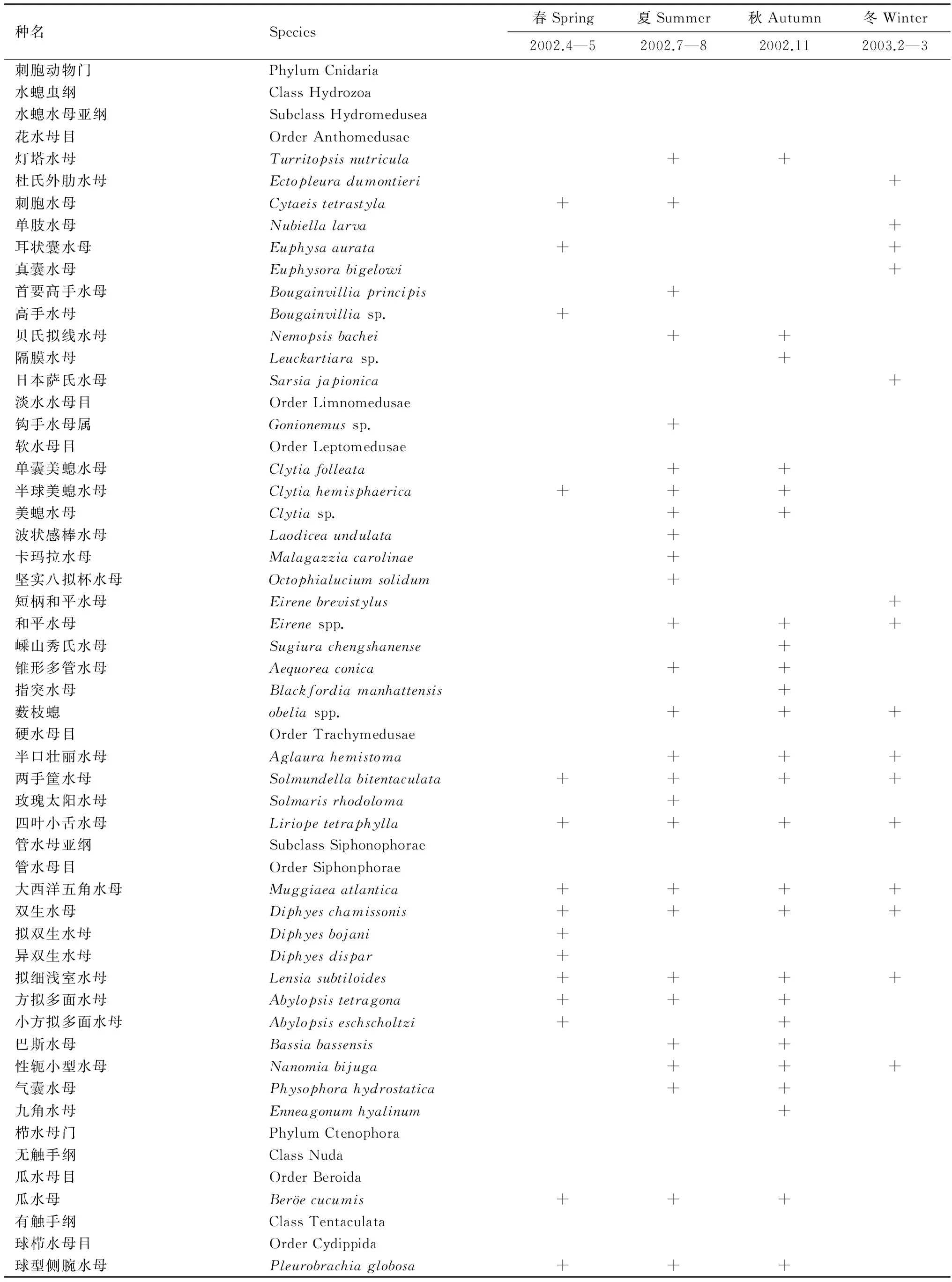
附录Ⅰ 长江口及邻近水域水螅水母、管水母和栉水母种名录Appendix Ⅰ Species list of medusae, siphonophores and ctenophores in the Changjiang Estuary and the adjacent East China Sea
国家重点基础研究发展计划(973计划)(2010CB428705);海洋公益性行业科研专项经费项目(201305027-8)
2014- 03- 21;
日期:2015- 04- 20
10.5846/stxb201403210505
*通讯作者Corresponding author.E-mail: xiaomin@sh163.com
高倩, 陈佳杰,徐兆礼,朱德弟.长江口及邻近海域浮游水螅水母、管水母和栉水母的丰度分布与季节变化.生态学报,2015,35(22):7328- 7337.
Gao Q, Chen J J, Xu Z L, Zhu D D.Abundance distribution and seasonal variation of medusae, siphonophores, and ctenophores in the Changjiang (Yangtze River) Estuary and the adjacent East China Sea.Acta Ecologica Sinica,2015,35(22):7328- 7337.
