三峡水库长期水淹条件下耐淹植物甜根子草的资源分配特征
2015-02-05潘晓娇苏晓磊
姚 洁,曾 波,杜 珲,潘晓娇,苏晓磊
三峡库区生态环境教育部重点实验室,重庆市三峡库区植物生态与资源重点实验室,西南大学生命科学学院, 重庆 400715
三峡水库长期水淹条件下耐淹植物甜根子草的资源分配特征
姚 洁,曾 波*,杜 珲,潘晓娇,苏晓磊
三峡库区生态环境教育部重点实验室,重庆市三峡库区植物生态与资源重点实验室,西南大学生命科学学院, 重庆 400715
为研究历经三峡水库长期水淹的驯化后,不同海拔高程之间相同种源的甜根子草的生物量分配特征是否发生改变,进而探讨该物种对水淹胁迫表现出的适应性进化特征,试验于2008年初选育相同种源的甜根子草同龄幼苗栽植于三峡水库消落区甜根子草种植试验示范区,并考察了2012年、2013年不同海拔高程甜根子草植株的形态和生物量特征。试验共设置3个海拔高程,即水淹高程168、172m和不受水淹对照高程176m。试验结果表明:(1)较低高程的甜根子草植株较矮小细弱,168m高程的甜根子草植株主茎长和主茎基径显著低于对照176m高程(P<0.05);平均节间长度随高程的降低而缩短;与之相反,主茎长/主茎基径随高程的降低而增大。(2)甜根子草的叶片厚度、叶片长/叶片宽、叶片长/叶鞘长均随海拔高程的降低而减小;与之相反,比叶面积随高程的降低而增大。(3)水淹前,甜根子草近端成熟节间的质量密度随高程的降低而增大;水淹后,其地上存活茎段基部成熟节间的质量密度在各高程之间无显著差异(P>0.05)。以上研究结果表明,甜根子草历经三峡水库长期水淹的驯化后,生物量分配特征在不同海拔高程之间发生了改变,表现出了相应的驯化特征。相较于高高程的甜根子草植株而言,低高程的植株生长缓慢,采取低株高下的高向生物量投资策略;对叶的物质投资大部分分配到叶面积的增加、叶鞘的伸长生长和叶片的直立生长上,以加强植株的光合生产。
三峡水库;甜根子草;水淹强度;生物量分配;形态性状
植物生长发育过程中各种生理功能的实现要求投入一定的资源[1],不同功能之间的资源分配存在竞争和权衡[2-3]。这就要求在自然选择的压力下,植物有理想的生物量分配策略,需要为实现一些优先的功能而以其他功能为代价[4]。植物响应不同生境的物质和能量的任何权衡分配过程都将导致生物量分配的差异[1],这是植物对异质性生境适应的结果。例如弱光环境下,紫茎泽兰(Eupatoriumadenophorum)和兰花菊三七(Gynurasp.)选择增加叶片生物量投入,增大比叶面积和叶面积比[5];苦瓜(Momordicacharantia)将更多的生物量投资到叶面积的增加、茎和叶柄的伸长上[6],以利于其搜寻优越的光环境。在不同土壤养分条件下,植物也会调整自身对土壤养分的获取对策。中等养分条件下,蛇莓(DuchesneaindicaFocke)和绢毛匍匐委陵菜(Potentillareptansvar.sericophylla)对茎的生物量投资最大,而在低养分条件下,其对根系的生物量投资增加[7-8]。部分水淹胁迫下,Lotustenuis茎伸长,使植株重新接触大气。相反,完全水淹胁迫下的植株相对静止,没有发生茎的伸长生长,从而保存能量[9]。由此可见,当处于异质性生境时,植物会通过适当改变其生物量分配来提高自身的适应及竞争能力。
三峡水库建成以后,根据其“蓄清排洪”的调度原则,在6—9月的自然汛期,三峡水库维持在防洪限制水位(145 m)运行,10—12月的蓄水期内,水位回升到175 m,一直到第2年4月都维持在较高的水位[10]。因此,其环境的异质性主要体现在库岸不同海拔高程被水淹没的时间长短、淹没的深度不同。甜根子草是生长于库区内的一种高大禾草,分布于库岸不同海拔高程,在稳定河岸和保持水土方面都有重要的生态作用。当甜根子草植株被水淹没的时候,光合生产减少,同时,淹没引起氧气供应减少,导致无氧呼吸加剧,碳水化合物的利用效率降低,体内营养物质消耗加速[11],植株因光合生产减弱和碳水化合物的大量消耗而更易“饥饿”,这对植株抵御水淹逆境是不利的[12]。可以想象,在每年都会发生的这种周期性水淹下,如果植株在水淹前加强光合生产以提供丰富的碳水化合物,同样在水淹后加强光合生产以快速补充水淹过程中的大量营养消耗,那将有利于植物在水淹胁迫下的存活及淹后的复苏。有研究发现植物在异质性生境中可以通过改变茎的垂直生长、节间长度、叶柄长度、叶片着生角度以及叶面积等将整个植株或觅食位点置放于一个较优越的生境中,以提高植物的觅食效率[13-14],促进光合生产。这种形态上的变化是植物资源获取行为的直观体现。
为此,本试验选育相同种源的甜根子草同龄幼苗于2008年初栽植于三峡水库消落区甜根子草种植试验示范区,并调查了2012年、2013年不同海拔高程甜根子草植株的形态和生物量特征,以期研究历经三峡水库长期水淹的驯化后,不同海拔高程之间的相同种源的甜根子草的生物量分配特征是否发生改变,进而探讨该物种在不同强度水淹胁迫下的资源获取行为和最适生长反应对策。
1 材料和方法
1.1 研究地概况
研究地点为西南大学三峡水库消落区野外研究实验基地(107°30′—108°14E,30°03′—30°35′N),地处三峡库区中部,属于典型的丘陵地貌,该地雨量充沛,日照充足,年均温18.2℃,日照时数1327.5h,年降雨量1200mm,空气相对湿度80%[15]。三峡水库修建之前,为农用坡耕地,土壤类型为黄壤。库岸不同高程均无明显水土流失状况。
1.2 试验材料
甜根子草(SaccharumspontaneumLinn.),禾本科,多年生草本植物,具有发达的根状茎,固土力强,是巩固河堤的保土植物。本研究中的材料取自于西南大学三峡水库消落区野外研究实验基地(基地面积约300m2)内的甜根子草植被,该植被采用由西南大学三峡库区生态环境教育部重点实验室选育的相同种源的甜根子草同龄幼苗于2008年初构建形成。株行距50cm×50cm,栽植后每隔1d浇1次透水,直至分蘖苗成活。其栽植下限延至海拔168m,上限延至海拔176m。经过历年水淹后,168m以上甜根子草均保存完好。
1.3 试验方法
1.3.1 试验设计
本研究以海拔高程表征水淹强度,共选择3个海拔高程水平,分别为:受三峡水库蓄水淹没的高程168、172m;不受水淹的高程176m,对生长于消落区不同海拔高程的甜根子草的植株状态进行了研究。本试验中以一个冬蓄夏排蓄水周期为分界点,2012年水库蓄水前定义为水淹前,2013年水库排水库岸出露定义为水淹后(图1)。

图1 2012年7月1日至2013年8月1日三峡水库消落区野外研究实验基地区内水位变化 Fig.1 Water level at water-level-fluctuation-zone in the field research base from July 1st, 2012 to August 1st, 2013水位信息来自重庆水文水资源信息网(http://www.cqhwr.com/)
2012年9—10月,在每个高程水淹之前,从生长于每个高程的甜根子草植株中随机选择30株当年长出的不受遮蔽的正常健壮植株,测量每个植株的主茎长、主茎基径、平均节间长度。然后从主茎的基部剪断主茎,选取主茎远端长出的成熟全展叶1—2片(本实验中统称为远端叶,即2011年水淹结束后,新生植株恢复生长2—3个月后产生的成熟叶),测定其形态和生物量指标(具体测定指标见1.3.2)。选择主茎基部的第1个正常成熟节间(本实验中统称为近端成熟节间),剪取该节间,测定其形态和生物量指标(具体测定见1.3.2)。
2013年初在每个高程水淹结束时(即植物刚露出水面后),在各高程带中随机选择30株经历三峡水库蓄水而存活下来的甜根子草植株,截取其存活茎段基部约5cm长的节间段测定其形态和生物量指标(具体测定见1.3.2)。水淹结束后,当某个高程的新生植株主茎上长出的第1、2片真叶成熟后,马上在该高程随机选择30株退水后新生甜根子草植株,选取主茎上长出的第1、2片成熟全展真叶(本实验中统称为近端叶,即水淹结束后,新生植株产生的第一批新叶),测定其形态和生物量指标(具体测定指标见1.3.2)。
1.3.2 指标测定
用直尺(精确到0.1cm)测量目标叶的叶片和叶鞘的长、宽(最宽处测量)。完成上述测量后,对每叶剪取叶片中部约10cm长的叶片段,叶鞘上部约10cm长的叶鞘段,分别测量叶片段和叶鞘段的长、宽,使用精度为0.01 mm的日本孔雀PEACOCA厚度计-G-MT测厚仪避开中脉后测量叶片段厚度。将叶片段和叶鞘段在80℃下烘干至恒重后称量干重。
用直尺(精确到0.1cm)测量节间的长度,用精度为0.02mm的游标卡尺测量节间的中位长轴直径、中位短轴直径。将节间在80℃下烘干至恒重后称量干重。
1.3.3 数据分析
根据测定的数据计算以下指标进行分析:
(1)主茎长/主茎基径、叶片长/叶片宽、叶鞘长/叶鞘宽、叶片长/叶鞘长
(2)比叶面积(SLA) 、叶片质量密度:
SLA=叶片段面积(cm2) /叶片段干重(g)
叶片质量密度(g.cm-3) =叶片段干重(g) /叶片段体积(cm3)
(3)节间质量密度:将试验所测量的节间看成规则圆柱体的近似体,故目标节间的体积计算如下:
节间体积(cm3) =节间长(cm)×中位长轴直径(cm)×中位短轴直径(cm)×π/4
节间质量密度(g/cm3)=节间干重(g)/节间体积(cm3)
用统计分析软件SPSS16.0进行试验数据的处理和分析。利用one-way ANOVA分别对三峡水库消落区水淹前和水淹后生长于不同海拔高程的甜根子草的节间、叶片的形态和质量密度特征之间的差异进行检验,若差异显著,采用Duncan 进行多重比较,在方差分析时若方差不齐则进行数据变换保证方差齐性。其差异性(P<0.05)用字母标记法表示。文中的图表均用Origin8.6制作完成。
2 结果
2.1 不同高程甜根子草植株主茎和节间形态性状的变化
不同海拔高程甜根子草植株主茎和节间形态性状各异(图2)。海拔168m处的甜根子草植株主茎长和主茎基径显著低于对照176m高程(P<0.05),而海拔172m与海拔176m处的甜根子草植株的主茎长和主茎基径之间没有显著差异(P>0.05)。同时,甜根子草植株的主茎长与主茎基径之比随海拔高程的降低逐渐升高,海拔168m处的甜根子草植株的主茎长与主茎基径之比显著高于对照176m高程(P<0.05) 。此外,随着海拔高程的降低,甜根子草植株的平均节间长度逐渐缩短,且各海拔高程之间差异显著(P<0.05)(图2)。
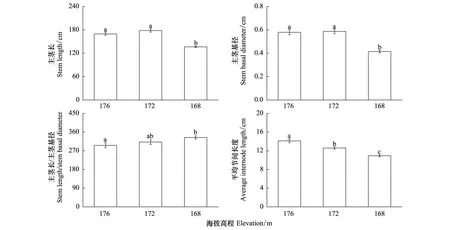
图2 不同海拔高程甜根子草的主茎长、主茎基径、主茎长/主茎基径和平均节间长度(平均值±标准误)Fig.2 Stem length, stem basal diameter, stem length/ basal diameter ratio and average internode length of Saccharum spontaneum Linn. growing at different elevations (mean±SE)
2.2 不同高程甜根子草植株叶形态性状的变化
甜根子草植株近远端叶的叶片厚度随海拔高程的降低而逐渐减小(图3),且168、172m高程的叶片厚度均显著低于对照176m高程(P<0.05),而比叶面积随海拔高程的降低而逐渐增加,168m高程显著高于对照176m高程(P<0.05)。
甜根子草对叶的物质投资大部分分配到叶鞘的伸长生长上。甜根子草植株近远端叶的叶片长与叶鞘长之比随海拔高程的降低逐渐降低(图2),且水淹高程168m显著低于对照176m高程(P<0.05)。近端叶的叶鞘长宽比随海拔高程的降低而逐渐升高,且168、172m高程显著高于对照176m高程(P<0.05)。试验中发现,随着海拔高程的降低,甜根子草植株近远端叶的叶片长宽比逐渐减小,且各高程之间差异显著(P<0.05)。
在本试验中,甜根子草远端叶的叶片质量密度在各海拔高程之间没有显著差异(P>0.05) ,但其新生近端叶的叶片质量密度随海拔高程的降低而逐渐增大,168、172m高程均显著高于对照176m高程 (P<0.05) (图3)。

图3 不同海拔高程甜根子草近远端成熟叶的叶片厚度,比叶面积,叶片长/叶鞘长,叶鞘长/叶鞘宽,叶片长/叶片宽和叶片质量密度(平均值±标准误)Fig.3 Leaf blade thickness, specific leaf area, leaf blade length/ sheath length ratio, leaf sheath length/ width ratio, leaf blade length/ width ratio and leaf blade mass density of the proximal and distal mature leaves of Saccharum spontaneum Linn. growing at different elevations (mean±SE)
2.3 水淹前和水淹后甜根子草植株茎成熟节间质量密度的分析
水淹前甜根子草植株近端成熟节间的质量密度随着海拔高程的降低而增大,且168、172m高程的节间质量密度均显著高于对照176m高程(P<0.05)(图4)。水淹结束后,甜根子草地上存活茎段基部节间的质量密度在各海拔高程之间无显著差异(P>0.05)(图4)。此外,植株水淹前和水淹后的节间质量密度之间的差异程度表现为168m >172m >176m。

图4 不同海拔高程甜根子草水淹前后的节间质量密度(平均值±标准误) Fig.4 The internode mass density of Saccharum spontaneum Linn. growing at different elevations before and after submergence (mean±SE)
3 讨论
植物在异质性生境中可能会改变其形态结构性和生物量配置而将植株置放在资源优越的生境中[13-14],这些变化提高了植物的觅食效率,而这些改变在一定程度上是由组织和器官的分配模式和对环境的反应决定的[16]。对众多的植物特别是陆生植物而言,三峡水库消落区的水淹是一种极端的环境胁迫。在水淹情况下,由于部分或全部叶组织没入水中,植株光合生产会降低;同时,水淹使植株处于缺氧或无氧状态,导致植物对营养物质的利用效率降低,营养物质消耗增大[11]。为了缓解水淹对植株造成的伤害,不同的耐淹植物能够采取不同的策略来适应水淹环境,有的植物可以加速枝条和叶片的伸长生长,以快速“逃离”水淹逆境[17-19],而有的植物则减缓生长,降低能量消耗,依赖大量的营养储备耐受长时间的水淹[18, 20-21]。本研究发现,低高程的甜根子草植株较矮小细弱(图2)。这可能是由于甜根子草在生产光合产物能力一定的情况下,168m高程的植株倾向于减缓生长,降低能量消耗,分配更多的资源用于贮藏,来抵御即将来临的水淹威胁,而172m高程和对照176m高程则倾向于将更多的资源投资到地上部分的再生产上。因此可以认为在光合生产能力一定的情况下,有限的生长及丰富的资源储备对于高强度水淹胁迫环境中的植株生存是有利的。随着海拔高程的降低,甜根子草植株的主茎长与主茎基径之比逐渐升高(图2),这表明与172、176m高程相比,168m高程的甜根子草植株采取基于低株高水平的纵向生物量投资策略,有利于将植株上的叶片放到相对较高较亮的位置,以利于获取较多的光资源,并延缓被水淹没的时间。
叶是植物利用光能、合成有机化合物的重要场所,在植物的生长发育过程中起重要作用,其形状特征直接影响到植物的基本行为和功能[22]。水淹来临前,近端叶逐渐枯萎,远端叶成为主要的功能叶,承担全部光合作用过程,为了在淹前提供更多的物质和能量,低高程甜根子草植株对远端叶的物质投资大部分分配到叶面积的增加、叶鞘的伸长生长和叶片的直立生长上(图3)。这可能是由于对长时间水淹压力的适应左右了甜根子草叶生物量分配的方向,将叶生物量从叶片厚度的累积转移到扩大叶面积上,从而改变光学路径来调节植物的吸光效率[23];将生物量转移到增加叶鞘伸长上,以便其快速获得阳光资源,避免其他叶片和植株的遮蔽作用。同时甜根子草植株权衡叶片长宽的资源分配额度,保持低高程甜根子草植株叶片的直立生长状态。与平展叶相比,直立叶是一种更加高效的叶部性状,叶片直立能使光能均匀地分布在全部叶片上,Duncan的研究结果表明,直立叶型玉米的光合生产率显著高于平展叶型玉米[24]。本实验中还发现低高程植株水淹前后节间质量密度差异最大(图4),即营养消耗最多,可以想象,为了填补水淹过程中由营养消耗造成的体内营养亏空,以进行良好的恢复生长,水淹刚结束后,甜根子草必须调整近端叶的生物量分配模式达到最佳程度,以保证退水后低高程植株能够快速获得空间和阳光资源。因此甜根子草植株的新生近端叶也表现出了和远端叶相同的生物量分配模式,即低高程的植株对近端叶的生物量投资大部分分配到叶面积的增加、叶鞘的伸长生长和叶片的直立生长上(图3)。明显的,较低高程的甜根子草植株对叶的生物量分配模式都朝着促进光合作用,为抵御年复一年的水库水淹提供强大的物质保障的方向发展。
水淹胁迫可以分为两个阶段:水淹阶段和出水阶段[25]。有研究认为,植物在水淹阶段的存活率与水淹之前的物质储备量有关,这主要是因为水淹之前的物质储备是供给植物在水淹下厌氧呼吸的能量来源[26-27]。植物出水后,其在水下产生的叶片多数无法适应有氧环境而迅速凋落,无法进行正常的光合生产[28],此时植物的初期生长和新的枝叶的产生需要依靠植株体内的物质剩余,也就是说水淹后物质的剩余状况在一定程度上影响了植株的初期恢复生长。主茎节间作为提供物质能量的来源之一,其生物量对于植物存活和恢复生长阶段叶的重新生长也是至关重要的。本研究中用节间质量密度刻画单位体积节间的重量(质量),节间质量密度反映了主茎节间内的干物质含量(生物量)。水淹前168、172m高程植株近端成熟节间的质量密度均显著高于对照176m高程(P<0.05)(图4),而水淹后甜根子草地上存活茎段基部成熟节间的质量密度在各海拔高程之间无显著差异(P>0.05)(图4),即在一定程度上反映了水淹前处在高强度水淹环境中的植株主茎节间的生物量更高,而水淹后主茎节间的生物量在各海拔高程之间无差异,这可能是对甜根子草在水淹中的存活和水淹后的快速恢复生长的一种适应机制。
在胁迫环境条件下,植物通过形态性状、生长发育和生理过程的调节,能在一定程度上降低其承受的环境压力,维持较高的适合度。这种能力对于加强物种抵御骤然变化的环境有特殊意义,它赋予了特定基因型个体一定的适应性弹性,有助于提高种群的生存能力[29]。以往研究认为,由环境诱导的表型变异主要源自发育的不稳定,是一种“环境噪音”,只在当代表现,不能遗传;但也有研究表明由环境诱导的变异并非仅仅是对局部异质生境适应状态的暂时调整,其结果有可能会影响物种或种群后续的选择进化[30]。由此可见,目前对此问题的看法仍未统一。通过本研究可以看出,相同种源的甜根子草历经三峡水库长期水淹的驯化后,生物量分配特征在不同海拔高程之间产生了差异,即相较于高高程的甜根子草植株而言,低高程的植株生长缓慢,采取低株高下的高向生物量投资策略;对叶的物质投资大部分分配到叶面积的增加、叶鞘的伸长生长和叶片的直立生长上,表现出了最适的资源获取行为和最适生长反应对策。至于其表现出来的适应性状能否遗传,这种调节能力是否存在随着水库蓄水周期的增加而减弱等问题,有待深入研究。
[1] 田迅. 松嫩平原湿地优势根茎植物种群生态学研究 [D]. 长春: 东北师范大学, 2004.
[2] Obeso J R. The costs of reproduction in plants. New Phytologist, 2002, 155(3): 321- 348.
[3] Bazzaz F A, Ackerly D D. Reproductive allocation and reproductive effort in plants // Fenner M. Seeds: The Ecology of Regeneration in Plant Communities. Wallingford, UK: CABI Publishing, 1992: 26- 26.
[4] 张洪轩. 植物生长可塑性研究 [D]. 长春: 东北师范大学, 2008.
[5] 王俊峰, 冯玉龙. 光强对两种入侵植物生物量分配、叶片形态和相对生长速率的影响. 植物生态学报, 2004, 28(6): 781- 786.
[6] 陶建平, 钟章成. 光照对苦瓜形态可塑性及生物量配置的影响. 应用生态学报, 2003, 14(3): 336- 340.
[7] 董鸣, 张淑敏, 陈玉福. 匍匐茎草本蛇莓对基质养分条件的克隆可塑性. 植物学报, 2000, 42(5): 518- 522.
[8] 张淑敏, 于飞海, 董鸣. 土壤养分水平影响绢毛匍匐委陵菜匍匐茎生物量投资. 植物生态学报, 2007, 31(4): 652- 657.
[9] Manzur M E, Grimoldi A A, Insausti P, Striker G G. Escape from water or remain quiescent? Lotus tenuis changes its strategy depending on depth of submergence. Annals of Botany, 2009, 104(6): 1163- 1169.
[10] 罗芳丽, 王玲, 曾波, 叶小齐, 陈婷, 刘巅, 张艳红, Kuhn A. 三峡库区岸生植物野古草(ArundinellaanomalaSteud.)光合作用对水淹的响应. 生态学报, 2006, 26(11): 3602- 3609.
[11] Gibbs J, Greenway H. Review: mechanisms of anoxia tolerance in plants. I. Growth, survival and anaerobic catabolism. Functional Plant Biology, 2003, 30(3): 353- 353.
[12] 张小萍, 曾波, 陈婷, 叶小齐, 罗芳丽, 刘巅. 三峡库区河岸植物野古草(Arundinellaanomalavar.depauperataKeng)茎通气组织发生对水淹的响应. 生态学报, 2008, 28(4): 1864- 1871.
[13] Dong M. Morphological plasticity of the clonal herbLamiastrumgaleobdolon(L.) Ehrend. & Polatschek in response to partial shading. New Phytologist, 1993, 124(2): 291- 300.
[14] Slade A J, Hutchings M J. The effects of light intensity on foraging in the clonal herbGlechomahederacea. Journal of Ecology, 1987, 75(3): 639- 650.
[15] 王祥荣, 蒋勇. 长江三峡库区(重庆段)沿江景观生态研究. 北京: 中国建筑工业出版社, 2006.
[16] Müller I, Schmid B, Weiner J. The effect of nutrient availability on biomass allocation patterns in 27 species of herbaceous plants. Perspectives in Plant Ecology, Evolution and Systematics, 2000, 3(2): 115- 127.
[17] Voesenek L A C J, Clomer T D, Pierik R, Millenaar F F, Peeters A J M. How plants cope with complete submergence. New Phytologist, 2006, 170(2): 213- 226.
[18] Voesenek L A C J, Rijnders J H G M, Peeters A J M, van de Steeg H M, de Kroon H. Plant hormones regulate fast shoot elongation under water: from genes to communities. Ecology, 2003, 85(1): 16- 27.
[19] Groeneveld H W, Voesenek L A C J. Submergence- induced petiole elongation inRumexpalustrisis controlled by developmental stage and storage compounds. Plant and Soil, 2003, 253(1): 115-123.
[20] He J B, Rijinders J G H M, van de Steeg H M, Rijnders J G H M, Voesenek L A C J, Blom C W P M. Survival tactics ofRanunculusspecies in river floodplains. Oecologia, 1999, 118(1): 1- 8.
[21] 罗芳丽, 曾波, 陈婷, 刘巅. 三峡库区岸生植物秋华柳对水淹的光合和生长响应. 植物生态学报, 2007, 31(5): 910- 918.
[22] 张林, 罗天祥. 植物叶寿命及其相关叶性状的生态学研究进展. 植物生态学报, 2004, 28(6): 844- 852.
[23] Enríquez S, Pantoja-Reyes N I. Form-function analysis of the effect of canopy morphology on leaf self-shading in the seagrassThalassiatestudinum. Oecologia, 2005, 145(2): 234- 242.
[24] Duncan W G. Leaf angles, leaf area, and canopy photosynthesis. Crop Science, 1971, 11(4): 482- 485.
[25] Ram P C, Singh B B, Singh A K, Ram P, Singh P N, Singh H P, Boamfa I, Harren F, Santos E, Jackson M B, Setter T L, Reuss J, Wade L J, Pal Singh V, Singh R K. Submergence tolerance in rainfed lowland rice: physiological basis and prospects for cultivar improvement through marker-aided breeding. Field Crops Research, 2002, 76(2- 3): 131- 152.
[26] Sarkar R K. Saccharide content and growth parameters in relation with flooding tolerance in rice. Biologia Plantarum, 1997, 40(4): 597- 603.
[27] Vriezen W H, Zhou Z Y, Van Der Straeten D. Regulation of Submergence - induced Enhanced Shoot Elongation inOryzasativaL. Annals of Botany, 2003, 91(2): 263- 270.
[28] Sarkar R K, Reddy J N, Sharma S G, Ismail A M. Physiological basis of submergence tolerance in rice and implications for crop improvement. Current Science, 2006, 91(7): 899- 906.
[29] 郭泉水, 洪明, 裴顺祥, 王祥福, 聂必红, 杨永明, 梁洪海. 香根草形态性状和光合特性对三峡库区消落带水陆生境变化的响应. 西北植物学报, 2012, 32(11): 2328- 2335.
[30] Gao L X, Chen J K, Yang J. Phenotypic plasticity: Eco-Devo and evolution. Journal of Systematics and Evolution, 2008, 46(4): 441- 451.
Effects of long-term submergence on resource allocation ofSaccharumspontaneumLinn. in Three Gorges reservoir
YAO Jie, ZENG Bo*, DU Hui, PAN Xiaojiao, SU Xiaolei
KeyLaboratoryofEco-environmentsinThreeGorgesReservoirRegion(MinistryofEducation),ChongqingKeyLaboratoryofPlantEcologyandResourcesResearchinThreeGorgesReservoirRegion,CollegeofLifeScience,SouthwestUniversity,Chongqing400715,China
To reveal the biomass allocation response ofSaccharumspontaneumLinn. at different elevations to long-term submergence in Three Gorges Reservoir, the adaptability ofS.spontaneumto submergence was studied. Similar-sized seedlings ofS.spontaneumwith the same provenance were planted at different elevations in our study area located in the hydro-fluctuation zone of the Three Gorges reservoir region in 2008. The morphological traits and biomass of the plants were examined in 2012 and 2013. Three elevations (in the submerged area: 168 m and 172 m; non-submerged area: 176 m) were measured. The results showed that: (1) The plants at the lower elevation were shorter, and the length and basal diameter of stems at 168 m elevation were significantly lower than those at 176 m elevation (P< 0.05). The ratio of stem length to basal diameter significantly increased with decreasing elevation, while the average internode length decreased with decreasing elevation. (2) The leaf blade thickness, the ratio of leaf length to leaf width, and the ratio of leaf length to sheath length decreased with decreasing elevation, but specific leaf area increased with decreasing elevation. (3) The mass density in basal internodes showed no significant difference at different elevations after submergence (P> 0.05), although there was a significant increment with decreasing elevation before submergence. These results demonstrated thatS.spontaneumcould decrease the growth of seedlings, increase leaf area and sheath elongation, and increase the hyponastic growth of leaves to adapt to long-term submergence at different elevations. This suggests that the alteration of biomass allocation might be a survival strategy forS.spontaneumwhen exposed to long-term submergence.
Three Gorges reservoir;SaccharumspontaneumLinn.; submergence intensity; biomass allocation; morphological character
国家重点基础研究发展计划项目(2012CB723205);国家自然科学基金资助项目(31370443,31070474,30440035);教育部高校博士点基金课题(20100182110022);中央高校基本科研业务费专项资金项目(XDJK2013A003)
2014- 03- 03;
日期:2015- 04- 20
10.5846/stxb201403030363
*通讯作者Corresponding author.E-mail: bzeng@swu.edu.cn
姚洁,曾波,杜 珲,潘晓娇,苏晓磊.三峡水库长期水淹条件下耐淹植物甜根子草的资源分配特征.生态学报,2015,35(22):7347- 7354.
Yao J, Z B, Du H, Pan X J, Su X L.Effects of long-term submergence on resource allocation ofSaccharumspontaneumLinn. in Three Gorges reservoir.Acta Ecologica Sinica,2015,35(22):7347- 7354.
