典型红壤区自然生态修复的适用性
2015-01-19钟炳林曹世雄
马 华, 钟炳林, 岳 辉, 曹世雄
1 广西壮族自治区水利科学研究院, 南宁 530023 2 福建省长汀县水土保持监测站, 龙岩 366300 3 北京林业大学经济管理学院, 北京 100083
典型红壤区自然生态修复的适用性
马 华1, 钟炳林2, 岳 辉2, 曹世雄3,*
1 广西壮族自治区水利科学研究院, 南宁 530023 2 福建省长汀县水土保持监测站, 龙岩 366300 3 北京林业大学经济管理学院, 北京 100083
自然修复主要通过封山育林、禁止农作、禁牧禁伐措施,减少人类对环境的扰动,利用自然生态环境的自我演替能力,恢复生态环境,实现生态平衡。自然修复作为一种成本低、无污染的生态修复手段很早就受到人们重视,但关于自然修复适用范围的研究较少。为了正确认识自然修复的适用性,选择了我国南方红壤地区长期遭受严重土壤侵蚀危害的福建省长汀县为研究对象,通过对长期自然修复样地的监测资料分析,发现在坡度条件为20%—30%下,当植被覆盖度低于20%的退化阈值时,严重的土壤侵蚀引发的土壤肥力损失将导致生态系统自我退化,自然修复不仅无法改善当地的生态系统,反而会引起生态系统的进一步恶化。由此可见,自然修复并不适合所有的生态系统,当生态系统退化到一定程度时,退化生态系统必须通过人工干预来修复。因此,必须探索适合当地的生态修复模式,在生态系统退化突破阈值时,红壤丘陵区应通过恢复土壤肥力、促进自然植被覆盖度增加、综合提高生态系统健康水平。
生态阈值; 生态修复; 植被覆盖度; 物种多度; 土壤肥力; 生态系统
自然修复或者人工修复在退化生态系统修复中的适用性一直以来都是生态学研究的重要方向。如果一个地区土壤退化十分严重并超过了生态系统内部自然修复的能力时,禁止人为干预或者进行长期封禁,生态系统都无法恢复到原始状态。这是由于植被退化导致表层土壤流失和土壤肥力下降,原有植被生长受限,同时其他原始物种难以入侵[1-2]。尽管生态修复措施被视为是增加植被覆盖度,提高生态系统服务功能的有效手段,但退化生态系统程度不同,人工选择植被的种类和数量、物种之间的相互作用等多种原因使得生态修复的效果有时不尽人意。因此,环境生物学最大的挑战之一是预测人类活动对物种和生态系统相互复杂关系的影响[3]。
当外部扰动引起主要物种、土壤或者生态群落内其他资源发生巨大变化时,生态群落结构极有可能表现出突然变化,即阈值变化[1,4]。很多研究引入并阐述了生态修复中生态阈值的概念[1,5-7],以期达到预测生态系统变化的目的。但是大部分早期的研究是理论性推测和模拟的结果,基于长期野外观测试验的阈值研究相对较少[6,8-9],使得各种生态阈值的理论模型缺少足够的数据验证[10]。由于不同类型的退化生态系统存在显著的地域差异性,何种生态阈值模型适用于何种特定环境缺乏相应的认识[11]。研究学者对半干旱地区过度放牧引起土壤退化不可逆转的研究较多[12-13],但针对湿润地区与生态退化有关的阈值研究相对较少。因此,针对湿润地区生态系统阈值特征的研究就显得更加迫切。
非生物与土壤或水分条件的变化决定了相应的物种可以在特定的地区生存,这些因素的生物阈值被广泛认为是修复退化生态系统的重要因素[5]。在退化土地管理中,阈值反映了植被和土壤的复杂变化,生态修复中如何掌握人为干预(如植树造林)和自然修复(如封山禁牧)的量度还很困难[14-16]。不同的生态阈值类型在生态系统管理中的作用不尽相同[17],正确应用阈值理论进行生态修复,可以起到事半功倍的效果;相反,盲目使也可能导致管理工作方面的缺失,给生态修复工作造成难以逆转的危害。如果环境管理人员通过研究生态阈值,了解不同的生态阈值适用于与之相对应的生态系统,确定合适的生态修复政策,因地制宜的生态修复措施将更加有效。为了证明生态退化阈值的广泛存在,并验证自然修复的适用范围,1986年起,选择位于温带湿润区的福建省长汀县开展长期观测试验,从而更好地了解生态退化阈值并指导红壤丘陵地区退化生态系统的修复工作。
1 研究方法
1.1 研究区概况
我国红壤丘陵区广泛分布于热带、亚热带,包括长江流域的大部分及其南部,跨南方10个省区,面积113万km2,占国土总面积的11.8%,人口约占全国的30%。近一个世纪以来的森林采伐、以及单一农作方式,导致植被覆盖度减少、植物多样性丧失、水土流失严重、洪涝灾害频繁、森林景观破坏等一系列环境问题[18-19]。与我国北方生态脆弱区相比,这里的环境问题有过之而无不及。新中国成立之后,这种形势并未得到有效改变,毁林毁草、开垦荒地的面积不断扩大。因此,这一地区的生态修复研究更加棘手[20]。2000年开始,福建省每年从省财政拨出1000万人民币,在福建省长汀县策武乡、河田镇、三洲乡、和濯田镇开展以可持续发展为目标、自然与社会相和谐的生态修复项目,并取得积极进展,引起国际社会的广泛关注[21]。为此,选择全国最早开展水土保持研究的四大监测区之一的福建省长汀县开展研究,从而更好地了解阈值变化对全国红壤丘陵地区退化生态系统管理的作用。
研究区域位于福建省长汀县,属温带湿润区(25°18′40″ N—26°02′05″ N, 116°00′45″ E—116°39′20″ E),具有温和湿润的亚热带季风气候特征。该县面积为309 720 hm2,年平均降水量为1737.1 mm,最大年为2552 mm,最小年为1074 mm,年平均蒸发量为1443.8 mm。全县年均气温18.3℃,年最低气温-7.9 ℃,最高温度39.4℃。长汀土壤以红壤为县内主要土壤资源,分布广、面积大,占土地总面积的79.8%。红壤孔隙度小、抗蚀性差,降雨易在裸露的土壤表层形成径流,产生严重的土壤侵蚀、形成水土流失。长期以来,由于战争、薪材、以及以粮为纲粮政策,导致全县生态景观逐步退化。退化的土地植被覆盖度下降、在强降雨作用下极易产生严重的水土流失,形成洪涝灾害,导致全县生态环境进一步恶化[22]。
1.2 样地布设
为了研究福建省长汀县生态系统自然修复的潜力,本研究选择了长期封禁监测的水土保持样地作为研究的对象。样地选具有类似的地形地貌、坡度范围在20%—30% 的中坡位。选取时要做到避免跨越河流、道路、山脊,也不应靠近林缘。区域内的地貌以丘陵为主,矿物晶粒粗大的黑云母花岗岩在长期湿热气候条件下发育成红色风化壳。样地红壤土壤孔隙度小(>0.005 mm孔隙度为1.9%—18.7%)、水分渗透缓慢,<1 mm微团聚体含量高达600 g/kg以上、抗蚀性差,降雨易在土壤表层形成径流,产生严重的土壤侵蚀。样地以马尾松(PinusmassonianaLamb)为主的次生针叶林组成乔木层,其生长量极低,林下以稀疏灌丛为主,先锋草本为鹧鸪草(EriachnepallescensR. Br.)和芒萁(Dicranopterispedata)。先对多个样地进行植被覆盖度的调查,然后在实验处理林地内选出不同植被盖度的400 m2(20 m×20 m) 样地30块。有研究表明,植被盖度在30% 以上时,退化林地具有自我修复能力[23],但研究区域降水充沛,利于植被生长,因此估计该地区植被阈值可能低于30%。根据1986年原始的植被覆盖度,把这些地块分为6组(每组5个样地):(1)小于20%,(2)20.1至25%,(3)25.1至30%,(4)30.1至35%,(5)35.1至40%,(6)大于40%。从1986年起,为了恢复当地生态环境,当地已禁止在封禁区进行砍伐、放牧等活动。自然修复的方式为封山育林边缘树立封山育林区标志碑,向居民宣传封禁的目的和意义,并有工作人员24 h看护,严禁人为干扰,通过林内自我恢复能力进行植被修复。
1.3 样地监测
样地监测的时间从1986年开始,是由当地水土保持站工作人员完成每年植被覆盖度、物种丰富度、土壤侵蚀、土壤养分的调查和监测。
(1)植被覆盖度 为了估算研究样地的植被覆盖度,需要测量树木的冠幅、地面灌木、草本植物的覆盖度;为了使各样地数据具有可比性,相关监测数据应在较短时间测得。因此,在每年生长旺季(6月下旬至8月下旬),避免植被的发芽或落叶时期。选取样方内具有代表性的乔木进行覆盖度测量,用钢卷尺测量每棵树最大和最小树冠半径,测算单个树木冠幅,乘以样方内相同树种林木的数量可得乔木覆盖面积。如有树冠重叠的地方,仔细确定重叠的范围,减去重叠部分,避免重复计算。乔木覆盖度等于乔木覆盖面积与样地总面积的比值。在没有被树木覆盖地方,用两条10m相交的直角断面进行对角线调查采样,调查灌草植物地面覆盖度。如灌草有重叠的地方,确定重叠的范围,减去重叠部分。灌草覆盖度等于灌草总覆盖度与样地总面积的比值。
(2)植物物种丰富度(也称植物多度) 是指实验样地出现的植物种数。每年8月收集各样地所有植物物种样本进行鉴定,无法鉴定的植物的种类制成标本,由福建师范大学专门研究人员进一步确认。
(3)土壤侵蚀 在类似试验小区的斜坡设置长为20 m,宽为5 m的径流小区,在径流小区底部修建容量为15 m3的浆砌石沉沙池。在降雨后24h内把所有的泥沙清理出池底,抽取3个泥沙样品在105 ℃下烘干12 h,然后称重,计算侵蚀模数(单位面积土壤流失量)和径流量(地表径流占降水的比例)。
(4)土壤养分 每隔5a(即1986,1991,1996,2001,2006,2011年),在每个实验样地的对角线等距离选取3个样点。用环刀取30 cm表层土壤,测量土壤养分成分。土壤有机质含量通过油浴加热的重铬酸钾氧化法测定、全氮测定采用碱蒸馏法、全磷测定采用原子吸收分光光度法(Varian spectrophotometer; Varian Inc., Palo Alto,California)、全钾用氢氟酸与高氯酸消解法测定。
(5)表层土壤沙粒:在土壤养分监测的同时,用相同方式取5 cm表层土壤,用不同型号的筛子筛选土样,确定样品大于1 mm沙粒含量。
1.4 统计分析
该研究用SPSS软件(12.0)进行显著性(LSD)分析。
2 研究结果与分析
2.1 植被覆盖度和植物丰富度的变化
研究结果表明(图1),封育前样地植被覆盖度若在20%以下,经过25a的自然修复后,植被覆盖度比封育前略有减少,植被覆盖度不仅没有上升,反而出现不同程度的下降,表明自然封禁在一定程度上加剧了这些样地植被的退化。与之相比,经过自然修复后,植被覆盖度逐年呈上升趋势,植被能得到较好恢复,封禁前植被覆盖度在20%—25%的样地能恢复到80%左右;封禁前植被覆盖度大于25%的样地能恢复到100%。植物丰富度也存在类似现象,植被覆盖度低于20%的样地,其植物种数均低于7种。从图1可以看出,当监测样地植物种类少于7种时,经过25a自然修复后,纵向图形没有明显的升高,说明物种数量没有增加。当监测样地植物种类大于7种时,纵向当年的物种数均比前五年有所增加。各组统计分析均达到极显著水平(P<0.01)。因此,植被恢复的同时,物种数也会得到丰富,有利于构建完整的自然生态系统结构。
2.2 土壤养分变化
研究结果表明(图2),经过25a自然修复后,封育前植被覆盖度低于20%样地的土壤有机质、全氮、全磷、全钾含量与封育前植被覆盖度高于20%样地相比有明显的差异。封育前植被覆盖度低于20%样地的土壤有机质、全氮、全磷、全钾含量分别小于3.0、0.14、0.16、5.0 g/kg,自然修复后,土壤养分不仅没有改善,反而出现不同程度的下降,自然封禁在一定程度上加剧了这些样地土壤的退化。从图2可以看出,纵向看,封育前植被覆盖度高于20%样地当年的土壤养分均比前5a有所增加,2011年时达到最高水平。横向看,样地的植被覆盖度越高,其土壤养分状况越好。因此,植被覆盖度过低对土壤化学成分的改良能力相对较弱,植被的恢复有助于对土壤的改良,促进土壤与植物之间营养物质的循环。各组统计分析均达到显著水平(P<0.05)。
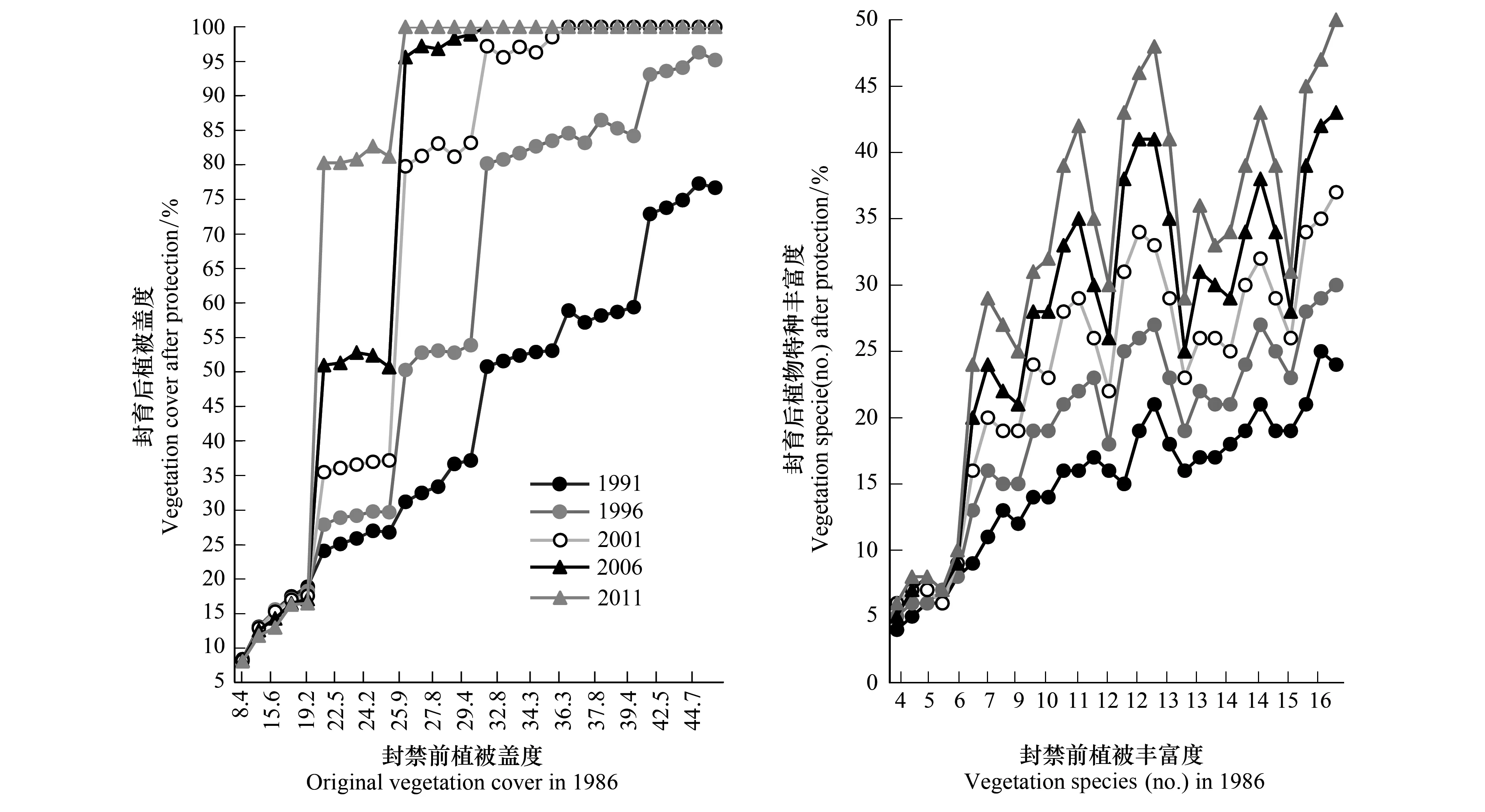
图1 实验样地25a自然修复植被覆盖度和丰富度变化Fig.1 Vegetation cover an richness change during 25 years natural restoration
2.3 水土保持效果
研究结果表明(图3),经过25a自然修复后,封育前植被覆盖度低于20%样地的土壤侵蚀模数和径流系数与封育前植被覆盖度高于20%样地相比差距极为明显。经过自然修复后,封育前植被覆盖度低于20%样地水土流失严重,当年土壤侵蚀模数和径流系数均较前5a略有增加。经过自然修复后,水土保持效果不仅没有改善,反而出现不同程度的恶化,自然封育在一定程度上加剧了这些样地的退化。然而,封育前植被覆盖度高于20%样地经过25a自然封禁的改造后,水土流失状况明显得到好转。纵向看,封育前植被覆盖度高于20%样地当年的水土保持能力均比前五年有所增加,2011年时达到最高水平。横向看,样地的植被覆盖度越高,其水土保持能力越好。因此,植被覆盖度的增加有利于防止水土流失的发生。各组统计分析均达到极显著水平(P<0.01)。
2.4 植被覆盖度与各指标之间的关系
相关分析结果表明,植被覆盖度与植物多度、土壤养分、水土保持效果之间存在极显著的线性相关关系(图4,P<0.001)。植被覆盖度与植物多度、土壤养分呈正相关,与土壤侵蚀模数、径流系数呈负相关。这是由于植被盖度的增加能减少雨滴的击溅,植被根系能固持土壤,枯枝落叶层不仅能增大地表粗糙度、吸收降水、还能有效延长下渗水分的时间。增加森林系统水土保持能力可以防止土壤营养成分的流失,为植被的生长提供养分供给。植被通过吸收土壤养分,促进了植被覆盖度的增加,同时也为土壤表层提供了充足的枯落物,其不仅能在微生物的分解下为土壤补充各类营养成分,以完成养分循环,还能有效阻止雨水的直接打击,减少了土壤养分随水流失。因此,植被覆盖度的增加是生态环境改善最直接的表现指标。
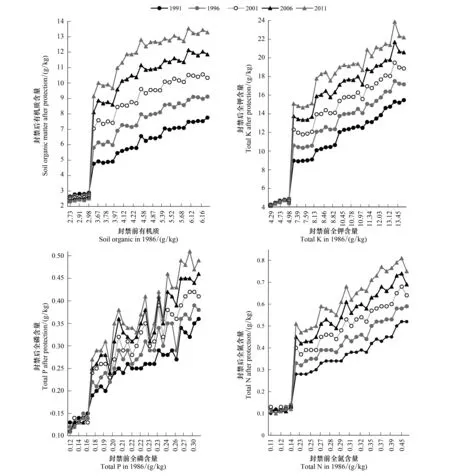
图2 自然修复样地中土壤养分含量变化Fig.2 Soil fertility change during 25 years natural restoration
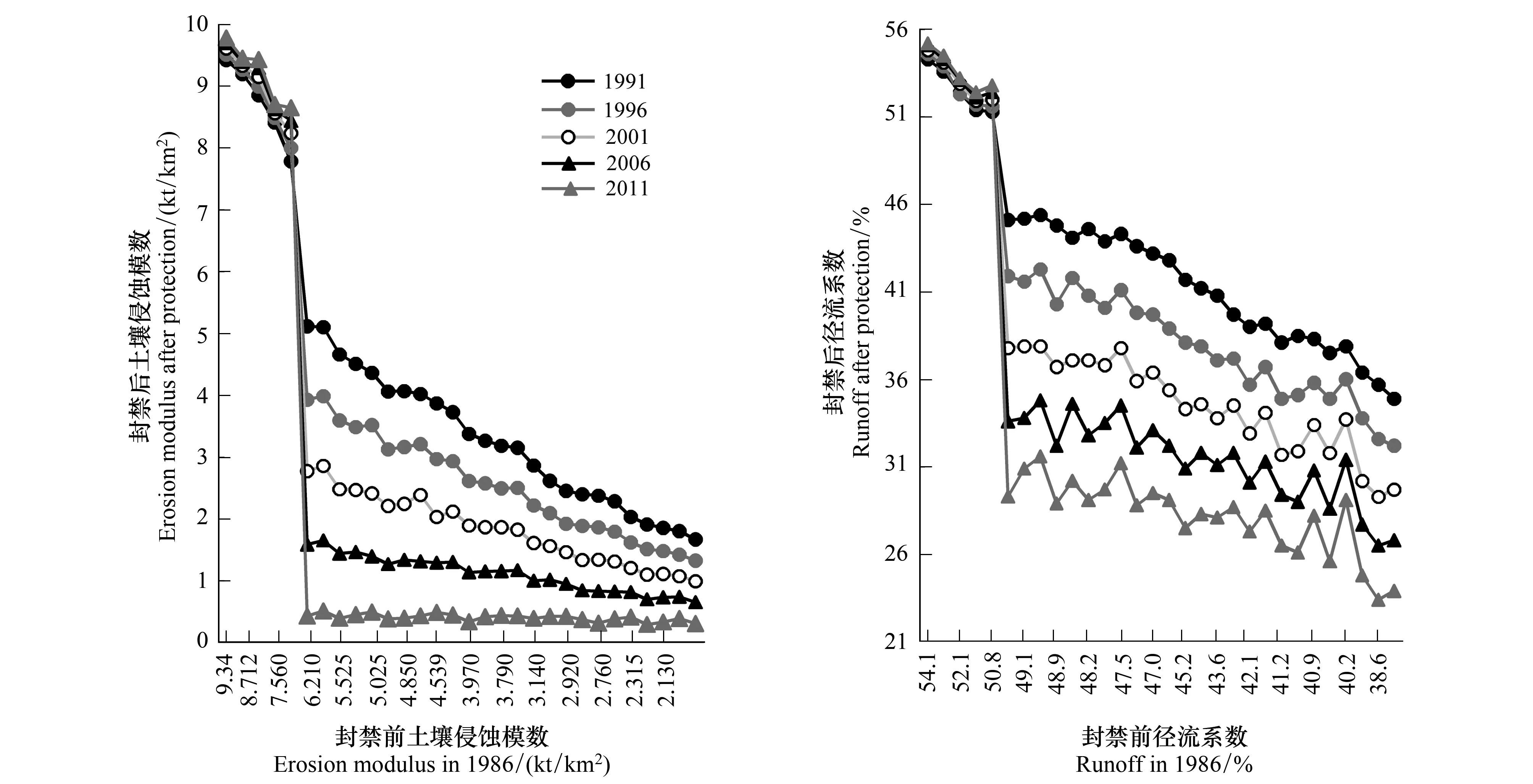
图3 自然修复后水土保持变化Fig.3 Soil and water conservation change during 25 years natural restoration
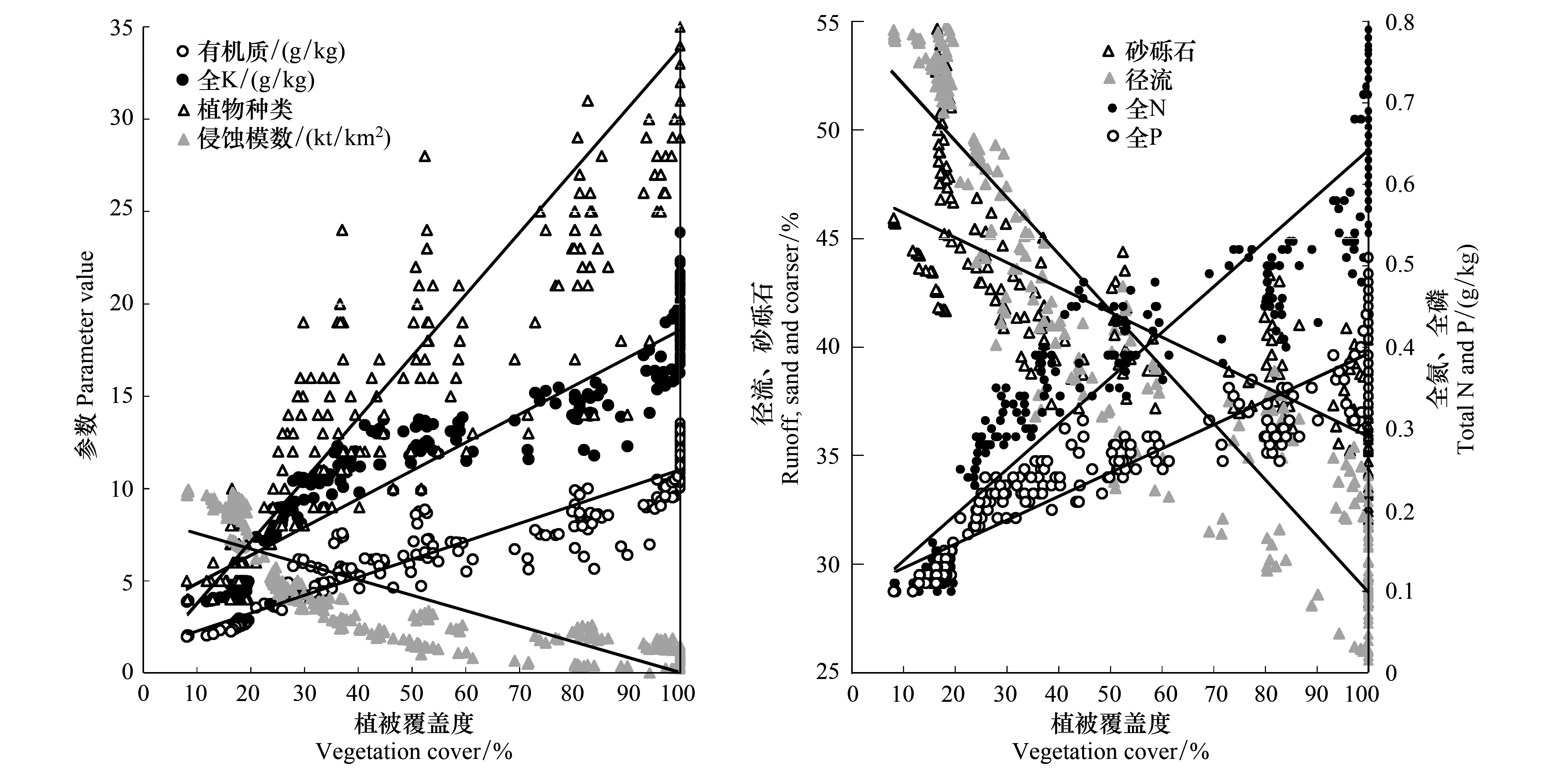
图4 监测样地植被盖度与其他指标的相关关系Fig.4 Relationships between vegetation cover and various soil and other parameters at the study plots
3 讨论
传统生态系统恢复的工作重点是重新建立干扰机制或者非生物条件,并依靠演替过程,促进生物群落的恢复。不过,生物因素和自然环境之间强烈的相互影响可以改变以这些演替为基础的效果[24]。因此,退化林地的修复结果往往出乎意料。很多研究学者认为,退化的生态系统处于极不稳定的状态,有时生态修复的方式会导致退化林地朝逆向方向发展[25-26]。确定退化生态系统处于何种状态,对于选择正确的生态修复方式有重要意义。结果表明,在20%—30%的坡度下,当植被覆盖度低于20%时,林地在没有人为干扰的情况下,植被覆盖度持续下降;在初始植被覆盖度高于20%的样地,即使没有人为干预,生态系统尚未丧失其自我恢复能力,即使不加以人为干预,也可为自然修复提供条件,通过缓慢恢复最终达到100%的植被盖度率(图1),土壤的肥力和其他土壤参数也逐渐改善(图2,图3),为天然植被恢复提供保障。实验结果显示:在20%—30%的坡度下,该湿润地区退化林地自然修复与人工修复之间的阈值为20%的植被盖度。由此可见,退化生态系统植被覆盖度大于20%时,可保持原始生态系统原本的结构、功能和反馈机制[14,16]。景观连续性和结构的变化、物种的丧失、优势物种的变化、不同营养层次的相互作用,都会引起土壤和其他生物的变化[6];如果自我恢复能力不足时,生态系统服务也会随之下降[27]。正确地应用阈值概念可以使不可逆的退化系统向自我修复的方向发展。
退化林区的生态修复与土壤质量密切相关,特别是与土壤养分的利用率有关[28]。结果表明,植被覆盖度与土壤养分状况呈显著相关性(图4),这是由于在长汀严重水土流失区,土壤养分的高低是限制植被恢复的重要因子。在自然修复样地中,丧失自我修复的植被群落,土壤肥力退化;具有自我修复能力的植被群落,土壤养分随植被覆盖度的增加而增加(图2)。在热带、亚热带降雨量充足的地区,加上砍伐、烧山等传统人工造林方式,使得长期累积的土壤有机物和养分容易在雨季被大雨淋失。长汀县也地处亚热带地区,降雨丰富,因此应当注重对表层土壤的保护,防治土壤养分随地表径流流失。植被不仅能改良土壤肥力状况,还能防治水土流失。结果表明,植被覆盖度和土壤养分与土壤侵蚀模数、径流系数、>1 mm砾石比例之间呈显著负相关。封禁前初始总植被覆盖度低于20%的自然修复样地由于植被退化,水土保持效果不仅没有改善,反而出现不同程度的恶化;随着植被覆盖度的增加,土壤侵蚀模数、径流系数下降,植被水土保持作用得到提高(图4)。植被盖度无法恢复,大面积的裸露表土在雨季被雨水溅蚀,径流冲刷,土壤养分不断流失。最终形成了土壤养分不断缺失,植被继续退化,土壤侵蚀量越大的恶性循环。
生态系统中的不同指标存在各自的阈值特征(图1,图3)。由于气候环境变化会影响物种生命周期的适应性变化;同时,大多数退化土地分布在不同的土地利用类型[29],这就使识别各种生态系统阈值特性非常困难[6];因此,生态学家一直希望开发出复杂的预测工具和更广泛的概念框架,帮助环境管理人员正确地应用阈值概念,防止生态系统出现不可逆的退化[29-30]。但是,这些研究框架对于多数管理人员和当地居民来说过于复杂,很少能够应用到生产实践来预测生态系统的临界变化[31],快速的环境变化使这项任务变得更艰巨。因此,我们必须找到简单易行的方法,快速评估变化生态系统的具体状态。植被覆盖度与其他生态指标之间的显著关系(图4),表明研究区用植被覆盖度变化来判断生态系统退化的阈值特征是可靠的。
自然生态系统具有复杂性特征[32],在受到人为干扰或者环境变化(如气候变化)时,生态系统自我恢复会更加困难,不适当的人工干预会引起生态系统进一步退化[9]。不同物种相互作用的复杂生态系统可能有不同的阈值特征[29,33]。以往研究结果表明,当生态系统退化超出阈值,就会引起本地物种的灭绝[27]。不同地区的研究结果表明,当植被覆盖率下降到10%—30%时,会发生不同程度的物种消失[8,34],这些退化的生态系统若不进行人为干预,将无法恢复到原始状态。在半干旱地区,低于30%植被覆盖度时,也有类似的结论[12-13]。在开展退化或者受损生态系统的修复时,必须了解生态系统的初始状态,并确定改变这种状态的一种或者多种关键因素[7]。不同地区的环境不同,生态修复的路径也不尽相同[32]。必须强调,不同地区之间存在自然因素差异,在20%—30%坡度条件下,20%植被覆盖度的阈值判断,只适用于中国南方红壤丘陵研究区域的特定环境。我们的研究结果对干旱地区并不适用,各地学者应针对不同地区的特征开展研究,并利用相关研究结果指导各地退化区域的环境管理。
[1] Lamb D, Erskine P D, Parrotta J A. Restoration of degraded tropical forest landscapes. Science, 2005, 310(5754): 1628-1632.
[2] Ludwig J A, Wilcox B P, Breshears D D, Tongway D J, Imeson A C. Vegetation patches and runoff-erosion as interacting ecohydrological processes in semiarid landscapes. Ecology, 2005, 86(2): 288-297.
[3] Berlow E L, Dunne J A, Martinez N D, Stark P B, Williams R J. Simple prediction of interaction strengths in complex food webs. Proceedings of the National Academy of the Sciences of the United States of America, 2009, 106(1): 187-191.
[4] Srinivasan U T, Carey S P, Hallstein E, Higgins P A T, Kerr A C, Koteen L E, Smth A B, Watson R, Harte J, Norgaard R B. The debt of nations and the distribution of ecological impacts from human activities. Proceedings of the National Academy of the Sciences of the United States of America, 2008, 105(5): 1768-1773.
[5] Norton D A. Species invasions and the limits to restoration: learning from the New Zealand experience. Science, 2009, 325(5940): 569-571.
[6] Sasaki T, Okayasu T, Jamsran U, Takeuchi K. Threshold changes in vegetation along a grazing gradient in Mongolian rangelands. Journal of Ecology, 2008, 96(1): 145-154.
[7] Jackson S T, Hobbs R J. Ecological restoration in the light of ecological history. Science, 2009, 325(5940): 567-569.
[8] Radford J Q, Bennett A F, Cheers G J. Landscape-level thresholds of habitat cover for woodland-dependent birds. Biological Conservation, 2005, 124(3): 317-337.
[9] Liu J G, Dietz T, Carpenter S R, Alberti M, Folke C, Moran E, Pell A N, Deadman P, Kratz T, Lubchenco J, Ostrom E, Ouyang Z Y, Provencher W, Redman C L, Schneider S H, Taylor W W. Complexity of coupled human and natural systems. Science, 2007, 317(5844): 1513-1516.
[10] Andersen T, Carstensen J, Hernandez-Garcia E, Duarte C M. Ecological thresholds and regime shifts: approaches to identification. Trends in Ecology and Evolution, 2009, 24(1): 49-57.
[11] Martin K L, Kirkman L K. Management of ecological thresholds to re-establish disturbance-maintained herbaceous wetlands of the south-eastern USA. Journal of Applied Ecology, 2009, 46(4): 906-914.
[12] Rietkerk M, van de Koppel J. Alternate stable states and threshold effects in semi-arid grazing systems. Oikos, 1997, 79(1): 69-76.
[13] van de Koppel J, Rietkerk M, Weissing F J. Catastrophic vegetation shifts and soil degradation in terrestrial grazing systems. Trends in Ecology and Evolution, 1997, 12(9): 352-356.
[14] Suding K N, Hobbs R J. Threshold models in restoration and conservation: a developing framework. Trends in Ecology and Evolution, 2008, 24(5): 271-279.
[15] McVicar T R, Li L T, Van Niel T G, Zhang L, Li R, Yang Q K, Zhang X P, Mu X M, Wen Z M, Liu W Z, Zhao Y A, Liu Z H, Gao P. Developing a decision support tool for China′s re-vegetation program: Simulating regional impacts of afforestation on average annual streamflow in the Loess Plateau. Forest Ecology and Management, 2007, 251(1/2): 65-81.
[16] McVicar T R, Van Niel T G, Li L T, Wen Z M, Yang Q K, Li R, Jiao F. Parsimoniously modeling perennial vegetation suitability and identifying priority areas to support China′s re-vegetation program in the Loess Plateau: Matching model complexity to data availability. Forest Ecology and Management, 2010, 259(7): 1277-1290.
[17] Bestelmeyer B T. Threshold concepts and their use in rangeland management and restoration: the good, the bad, and the insidious. Restoration Ecology, 2006, 14(3): 325-329.
[18] 陈志清, 福建省长汀县河田镇的水蚀荒漠化及其治理. 地理科学进展, 1998, 17(2): 67-72.
[19] 曾金华, 钟炳林. 长汀县水土流失演变及今后治理对策. 福建水土保持, 2002, 14(4): 37-39.
[20] 岳辉, 钟炳林. 长汀县水土保持监测实践与探讨. 亚热带水土保持, 2006, 18(3): 63-64.
[21] 杨学震, 钟炳林, 谢小东. 丘陵红壤的土壤侵蚀与治理. 北京: 中国农业出版社, 2005.
[22] Cao S X, Zhong B L, Yue H, Zeng H S, Zeng J H. Development and testing of a sustainable environmental restoration policy on eradicating the poverty trap in China′s Changting County. Proceedings of the National Academy of the Sciences of the United States of America, 2009, 106(26): 10712-10716.
[23] Lindenmayer D B, Fischer J, Cunningham R B. Native vegetation cover thresholds associated with species responses. Biological Conservation, 2005, 124(3): 311-316.
[24] Myers N. Environmental services of biodiversity. Proceedings of the National Academy of the Sciences of the United States of America, 1996, 93(7): 2764-2769.
[25] Suding K N, Gross K L, Houseman G R. Alternative states and positive feedbacks in restoration ecology. Trends in Ecology and Evolution, 2004, 19(1): 46-53.
[26] Klotzli F, Grootjans A P. Restoration of natural and semi-natural wetland systems in Central Europe: progress and predictability of developments. Restoration Ecology, 2001, 9(2): 209-219.
[27] Schlaepfer M A, Runge M C, Sherman P W. Ecological and evolutionary traps. Trends in Ecology and Evolution, 2002, 17(10): 474-480.
[28] Radford J Q, Bennett A F, Cheers G J. Landscape-level thresholds of habitat cover for woodland-dependent birds. Biological Conservation, 2005, 124(3): 317-337.
[29] Garten C T Jr, Ashwood T L. Modeling soil quality thresholds to ecosystem recovery at Fort Benning, GA, USA. Ecological Engineering, 2004, 23(4/5): 351-369.
[30] Suding K N, Gross K L, Houseman G R. Alternative states and positive feedbacks in restoration ecology. Trends in Ecology and Evolution, 2004, 19(1): 46-53.
[31] Chapin III F S, Robards M D, Huntington H P, Johnstone J F, Trainor S F, Kofinas G P, Ruess R W, Fresco N, Natcher D C, Naylor R L. Directional changes in ecological communities and social-ecological systems: a framework for prediction based on Alaskan examples. The American Naturalist, 2006, 168(S6): S36-S49.
[32] Byers J E, Cuddington K., Jones C G, Talley T S, Hastings A, Lambrinos J G, Crooks J A, Wilson W G. Using ecosystem engineers to restore ecological systems. Trends in Ecology and Evolution, 2006, 21(9): 493-500.
[33] Hunt D E, David L A, Gevers D, Preheim S P, Alm E J, Polz M F. Resource partitioning and sympatric differentiation among closely related bacterioplankton. Science, 2008, 320(5879): 1081-1085.
[34] Lindenmayer D B, Fischer J, Cunningham R B. Native vegetation cover thresholds associated with species responses. Biological Conservation, 2005, 124(3): 311-316.
Research on the application of natural ecological restoration in a typical region of China with red soils
MA Hua1, ZHONG Binglin2, YUE Hui2, CAO Shixiong3,*
1GuangxiInstituteofWaterResourcesResearch,Nanning530023,China2SoilandWaterConservationBureauofChangtingCounty,Longyan366300,China3CollegeofEconomicsandManagement,BeijingForestryUniversity,Beijing100083,China
Natural restoration is mainly accomplished by allowing natural reforestation to occur, combined with prohibiting agriculture and grazing to reduce human disturbance of the environment. Taking advantage of the succession that occurs in natural ecosystems is a common ecological restoration measure that can restore ecosystems and keep them in balance. However, conservation and restoration biologists have increasingly recognized that ecological communities are likely to exhibit threshold changes in structure that may prevent succession from occurring. Because long-term monitoring data are generally lacking, little is known about the consequences of such ecological thresholds for the processes of ecosystem degradation and recovery. To identify whether a degradation threshold exists that defines the boundary between the possibility of natural recovery and the need for artificial restoration of an ecosystem and to use this knowledge to support the development of a suitable strategy for environmental restoration, we have performed long-term monitoring of vegetation recovery in China′s Changting County since 1986. We found a severe problem in this area, which we refer to as the “irreversible loss of soil services”: when vegetation cover decreases below an ecological degradation threshold, leading to sustained degeneration of the vegetation community, erosion of the surface soil and declining soil fertility occur. These changes represent a severe and long-term disturbance of the vegetation, the soil, and the landscape. We identified a degradation threshold at about 20% vegetation cover, which suggests that for some sites with a vegetation cover of between 20 and 30%, vegetation cover can serve as a simple proxy for more sophisticated approaches to identifying thresholds; when vegetation cover declines to this level, restoration must start with the restoration of soil fertility and continue by facilitating vegetation development. Our results support the concept of ecological thresholds (specifically, for soil services in a warm and wet region of China) and provide a model to inform restoration strategies for other degraded ecosystems. Ecosystem restoration sometimes fails because ecological interactions are more complex or human intervention is more difficult than anticipated; factors other than human disturbance, such as climate variability (e.g., a drought shortly after planting), can result in failure of a strategy that would succeed under better conditions. Some degraded ecosystems can be sustained only through ongoing management, but many conservation efforts preclude such interventions. Although ecologists can recognize many of the species changes that are likely to precipitate threshold changes in community composition, biotic interactions can be unexpected, and because responses often depend strongly on local conditions, they cannot be broadly generalized. For example, complex ecosystems with multiple interacting species may have multiple thresholds based on different components of the ecosystem. Desertification is another example and has been shown to result from strong biogeomorphic feedbacks that operate across several spatial scales. When overgrazing of arid grasslands reduces vegetation cover, water infiltration decreases, further limiting plant growth and leading to persistent desertification. Such spatial discontinuities, calledecotones, can be detected using multivariate data ordered in one dimension through comparisons of measures of dissimilarity computed between the systems on either side of the discontinuity. The “irreversible loss of soil services” described in our study, whether at national, regional, or local scales will have a variety of thresholds, and it will be necessary to calibrate this index for different regions before it becomes a useful management tool. However, as our results show, it is possible to identify useful proxies for such thresholds and use them to guide subsequent management of degrading sites.
ecological thresholds; ecological restoration; natural recovery; species richness; soil fertility; ecosystems
国家自然基金重点项目(91025002)
2014-01-21;
日期:2014-11-19
10.5846/stxb201401210161
*通讯作者Corresponding author.E-mail: shixiongcao@126.com
马华, 钟炳林, 岳辉, 曹世雄.典型红壤区自然生态修复的适用性.生态学报,2015,35(18):6148-6156.
Ma H, Zhong B L, Yue H, Cao S X.Research on the application of natural ecological restoration in a typical region of China with red soils.Acta Ecologica Sinica,2015,35(18):6148-6156.
