阿拉善荒漠啮齿动物集合群落实证研究
2015-01-18查木哈武晓东付和平乌云嘎张晓东
查木哈,武晓东,付和平,袁 帅,乌云嘎,张晓东
1 内蒙古农业大学生态环境学院, 呼和浩特 010019 2 草业与草地资源教育部重点实验室, 呼和浩特 010019 3 内蒙古自治区林业监测规划院, 呼和浩特 010020
阿拉善荒漠啮齿动物集合群落实证研究
查木哈1, 2,武晓东1,2,,付和平1, 2,袁 帅1, 2,乌云嘎1, 2,张晓东3
1 内蒙古农业大学生态环境学院, 呼和浩特 010019 2 草业与草地资源教育部重点实验室, 呼和浩特 010019 3 内蒙古自治区林业监测规划院, 呼和浩特 010020
当生态学家探求在破碎化的栖息地中,群落物种的共存机制、多样性、局域尺度的性质和过程被放到更广阔的时空框架内时,就出现了“集合群落”这一概念。Leibold提出了集合群落概念,他们将一个集合群落定义为局域群落集,这些群落由各个潜在的相互作用的物种的扩散连接在一起。集合群落理论描述了那些发生在集合群落尺度上的过程,并且提出思考关于物种相互作用的新方法。集合群落概念为群落生态学提供了一个新的革命性的范式,集合群落研究的最基本问题是同一系统中多物种共存的机理、多样性的形成原因与维持机制。该范式强调区域范围内群落中的综合变异,强调环境特证和栖息地之间通过扩散调节的生物相互作用和空间变化。Leibold 等提出了解释集合群落结果理论上的4个生态范式,即(1)中性理论;(2)斑块动态理论;(3)物种分配理论;(4)集团效应理论。之后有大量有关检验这4种生态理论的研究,但是有关陆地脊椎动物系统的集合群落的研究较少。2010—2012年,通过在内蒙古阿拉善荒漠景观中的8个固定样地中,对啮齿动物、栖息地环境因子进行调查。利用冗余分析和偏冗余分析,评估环境特征和空间特征对物种组成的影响。结果表明,环境特征独自解释72.8%的啮齿动物物种组成变化,空间特征独自解释33.8%的物种组成变化,环境特征和空间特征共同解释86.5%的啮齿动物物种组成变化,结果显著(P= 0.032);去除环境特征之后,空间特征解释13.7%的变化(P= 0.246),结果不显著;去除空间特征之后,栖息地变化解释52.7%的变化(P= 0.016);环境特征和空间特征的交互作用解释20.1%的物种组成的变化,该区域啮齿动物群落构成集合群落,物种共存中环境特征起着主导作用,由物种分配理论解释该集合群落结构。
群落生态学;啮齿动物;集合群落;冗余分析
生态学家长期讨论关于共存物种集群(群落)特征的过程和格局。空间和时间格局能为群落经历的机制提供线索,其中一些长期存在和苛刻的争论已经关注能被解释的格局[1]。群落生态学已经赢得了新范式的快速进展,即结合局域和区域过程去更清楚地了解在区域中群落组群的空间结构[2]。群落生态学中局域相对于区域过程决定局域群落结构的相对重要性是中心论题[3]。群落生态学当前的挑战之一是缩小理论和观察的间隙以针对物种的多样性和物种共存[4]。过去的几十年来已经有许多研究专注于封闭种群的交互作用,即种间关系及物种与环境之间的关系怎样形成群落中物种组成的空间变化。集合群落概念为群落生态学提供了一个新的革命性的范式,通过在区域范围内强调群落中的综合变异,强调环境特点和栖息地之间通过扩散调节的生物相互作用和空间变化[5]。
集合群落研究的最基本问题是同一系统中多物种共存的机理、多样性的形成原因与维持机制,强调多物种的长期共存需要在某种尺度上体现出其重要生物学或生态学特征的权衡[5-6],或者说,权衡是群落理论的基石。Leibold 等[5]提出了解释集合群落结构理论上的4个生态范式,即(1)斑块动态理论,该范式假设斑块被确定而且每个斑块能够包含种群。斑块可能被占据或者不能被占据。局域物种多样性被扩散限制,空间动态由局域绝灭和建群主导。(2)物种分配理论,该范式强调资源梯度或者斑块类型引起充分强烈的差异在物种的局域群落统计学和局域物种相互作用上的结果。这种相互作用对斑块质量和加入扩散时影响局域群落组成。这个观念强调空间生态位分化超出了空间动态。扩散是重要的,因为它导致组成的变化能追踪局域环境条件的变化。(3)集团效应理论,该范式集中在局域种群动态上的迁入和迁出的效应。在这样的系统中,通过它的是优级物种的竞争者迁入群落的,物种能被拯救免于群落中的局域竞争排除,在这个群落中它们是差的竞争者。这个观念强调空间动态影响局域种群密度的作用。(4)中性理论,该范式指所有物种在竞争能力、运动以及适应性上相似[7],物种之间种群相互作用构成随机漫游。它改变物种相对频率。物种多样性的动态是源于物种丢失的(绝灭、迁出)和获得的(迁入、特化)的概率[5]。
解释集合群落结构理论上的4个生态范式被提出之后有大量有关检验这4种生态理论的研究,特别是在理论上和模拟群落结构的研究上较多[8- 12]。由于从区域角度评价物种共存的机制,但在广阔的空间范围内大多数的动物很难被证明集合群落结构[13]。国外已有较多证明集合群落结构的报道,但大部分的实证工作集中于无脊椎动物和小型水生动物,尤其是生活在相对有界限的湖泊群落[14- 19], 有关陆地脊椎动物系统的集合群落的研究较少[20- 23]。目前国际上有关集合群落的理论研究[8- 11]和实际观察研究[14- 24]由于针对目标动物不同(如浮游动物[14- 16]水体无脊椎动物[17]、陆地无脊椎动物[18- 20]和鸟类[21-22]与哺乳类[23-24]),调查与取样尺度各异,导致研究结果的多样化,仍然不能形成较普适的理论。国内有关集合群落的研究目前极其稀少。
在最近10年间人类通过破坏和破碎化生境对于世界上生态系统的影响极其剧烈地增加[25]。阿拉善荒漠是我国典型的温带荒漠,近10年由于人类生产活动的剧烈加剧使原来一致的景观遭到了严重的干扰,形成了明显的破碎化生境。本研究于2010—2012年,通过对内蒙古阿拉善荒漠区啮齿动物群落的调查,并且分析其栖息地环境特征和空间特征对啮齿动物物种组成变化的作用,检验人为干扰下生境破碎化过程中荒漠区啮齿动物集合群落结构特征,实际验证当今集合群落理论的范式,为集合群落理论的完善和发展提供实证资料和实验性证据。
1 自然条件与研究方法
1.1 研究区概况
研究区(104°10′—105°30′E,37°24′—38°25′N)位于内蒙古阿拉善左旗南部的荒漠景观中,属于亚洲中部荒漠亚区最东部的一个植物地理区域。该地区的草地类型是典型的温性荒漠,植被稀疏,结构单调,覆盖度低,一般仅1%—20%。植物种类贫乏,主要以旱生、超旱生和盐生的灌木、半灌木、小灌木和小半灌木为主。建群植物以藜科(Chenopodiaceae)、菊科(Compositae)和蒺藜科(Zygophyllaceae)为主,其次为蔷薇科(Rosaceae)、柽柳科(Tamaricaceae)。地形起伏不平,丘陵、沙丘与平滩相间。气候为典型的高原大陆性气候,冬季严寒、干燥,夏季酷热,昼夜温差大,极端最低气温-36 ℃,最高气温42 ℃。年平均气温9.0 ℃,无霜期156 d。年降水量45—288 mm,主要集中在7—8月份。年蒸发量3000—4700 mm。土壤为棕漠土,淋溶作用微弱,土质松散、瘠薄,表土有机质含量1%—1.5%,含有较多的可溶性盐。
1.2 研究方法
1.2.1 啮齿动物调查
在阿拉善左旗南部可利用草场中(面积为3393 km2)选择了8个样区(图1),用GPS定位每个样区中心(S1—S8),样地之间的距离为0.22—3.89 km,在每个样区中心建立1个固定的标志重捕样地,样地面积为1 hm2。每个标志重捕样地以方格式布笼,笼距15 m×15 m,每个网格点上布设1个活捕笼(42 cm × 17 cm × 13 cm),以新鲜花生米为饵。2010—2012年4—10月每月初连捕4 d。记录捕获个体种名、性别、繁殖状况、体重及捕获位置。

图1 样地空间分布示意图Fig. 1 The distribution schematic diagram of plots
1.2.2 植物和土壤调查
每个动物取样样区内随机选择3个10 m×10 m样方进行灌木取样,在每个灌木样方中随机选择1个1 m×1 m的草本样方,合计在每个动物取样区内取3个草本样。在这些样方中测量植物的“四度一量”。在灌木样方内对每种灌木分别选取大、中、小3个植株测量其高度,最大和最小冠幅。记录灌木样方内各灌木物种的密度,并取中等植株当年生枝条的十分之一称量鲜重(精度0.01 g)。土壤取样在每个灌木样方内与植物和动物取样同步进行,测量指标包括0—5 cm,5—10 cm,10—15 cm,15—20 cm土壤硬度。土壤硬度使用土壤硬度仪(1 Psi = 6.89 KP)测定。
1.2.3 数据处理
冗余分析(RDA)是约束性排序分析方法,1964年Jongman首次提出该方法,将样点投射到两条排序轴构成的二维平面上,通过样点的散集形态、用象限的分布等来反映研究区的特点。在统计学中,冗余分析是通过原始变量与典型变量之间的相关性,分析引起原始变量变异的原因。以原始变量为因变量,典型变量为自变量,建立线性回归模型,则相应的确定系数等于因变量与典型变量间相关系数的平方。它描述了由于因变量和典型变量的线性关系引起的因变量变异在因变量总变异中的比例[26]。它是多元线性回归的扩展,是一种多变量直接梯度分析方法。通过采用两个变量集的线性关系模型,得到数值矩阵并对特征值进行分解,能将表征环境特征及空间特征与物种的关系反映在坐标轴上。冗余分析通过每个群落环境特征和空间特征矩阵的全变异及P值来判别局域环境和空间过程的重要性,并用R2来检验变异量(计算群落矩阵全变异的百分比)。
偏冗余分析(Partial RDA)是通过完全排除另一组环境(E)或空间(S)特征来解释其变量。对于集合分析,偏冗余分析提供了一个通过解释变异的百分比来描述不同类型研究变量的方法。它根据数据的结构进行分析,具有很强的灵活性[14-15,23,27-28]。利用 Canoco 4. 5 软件进行冗余分析,分析步骤如下:(1)对啮齿动物物种组成、栖息地因子及空间因子均进行log(x+1)转换。(2)对啮齿动物物种变量进行去趋势对应分析(DCA),确定群落属于基于单峰模型的排序分析或线性模型的排序分析,若DCA第一排序轴的长度< 3适合线性排序法。(3)采用向前引入法逐步筛选显著环境变量,每一步均采用蒙特卡罗置换检验。(4)进行偏冗余分析时,先选择需要排除的因子,再对协变量进行删除。(5)利用蒙特卡罗置换检验测度RDA排序轴的显著性,显著性水平为0.05。
通过方差分析来反映环境特征和空间特征对物种组成的影响,分解因子包括:总解释变异(E+S),环境变异(E),空间变异(S),不包括空间的环境变异(E|S),不包括环境的空间变异(S|E)。对空间变异及环境变异进行冗余分析和偏冗余分析得到各成分的变异:(1)空间和环境变异共同解释啮齿动物物种组成的总变异(E+S):为了揭示啮齿动物组成的全变异,以啮齿动物物种组成为因变量,环境和空间变量组成为自变量,进行冗余分析。(2)纯空间变异解释啮齿动物物种组成的部分变异(S|E):为了揭示纯空间成分,以啮齿动物物种组成为因变量,空间变量为自变量,环境因子为协变量,进行了偏冗余分析。(3)纯环境变异解释啮齿动物物种组成的部分变异(E|S):为了揭示纯环境成分,以物种组成为因变量,以环境因子为自变量,空间因子为协变量进行了另一个偏冗余分析。(4)空间和环境变异都解释啮齿动物物种组成的部分变化(E∩S):空间结构和环境因子的交互作用,从被解释的全变异中减去纯空间变异及环境变异。
本研究重点讨论暂时动态,选取了每年7月份的调查数据进行分析。动物及植物数据为3年数据的平均值。捕多次算一次,物种组成用捕获量比例来表示。表征环境特征参照Stevens 和 Tello对莫哈韦荒漠啮齿动物集合群落研究时选取的环境因子,选用了灌木生物量、草本植物盖度和土壤硬度[24];表征空间特征选用了经纬度坐标,空间因子由每个样区中心的经纬度坐标转换为投影坐标而组成[23-24,28]。
1.2.4 集合群落模型的判别
物种分配理论:物种分配模型假设在一个集合群落中物种通过强烈响应环境特征使栖息地相联系,并且扩散不足以影响物种在特殊的斑块中续存[29]。空间特征不显著,不能解释物种组成的显著变化。相反,环境特征显著,能够解释物种组成的变化。
集团效应(Cottenie的集团效应+物种分配的观点)[28]:通过物种对环境特征的响应来联接栖息地,但是扩散通过源-汇动态允许物种持续的在较少的斑块中生存[13]。在集团效应中解释物种组成的变化时环境和空间特征均显著。在方差分析中,纯环境与纯空间变异均显著,空间和物种-环境的相互作用的程度反映集合群落结构。
中性理论模型:从生态学观点来看全部物种都是等值的,扩散、竞争能力、出生率和死亡率的差异很小。物种与栖息地特征关系不密切,集合群落样地之间的空间变化结构仅被空间过程决定。因此,空间变异独自解释物种组成的变化,只有纯空间特征显著[7]。
斑块动态理论模型:与中性理论模型相似,空间过程对集合群落结构的确定起重要作用。主要区别于暂时的动态,扩散和竞争的权衡引起样地之间物种组成的不同[30]。表1为集合群落结构类型的判断树[28]。
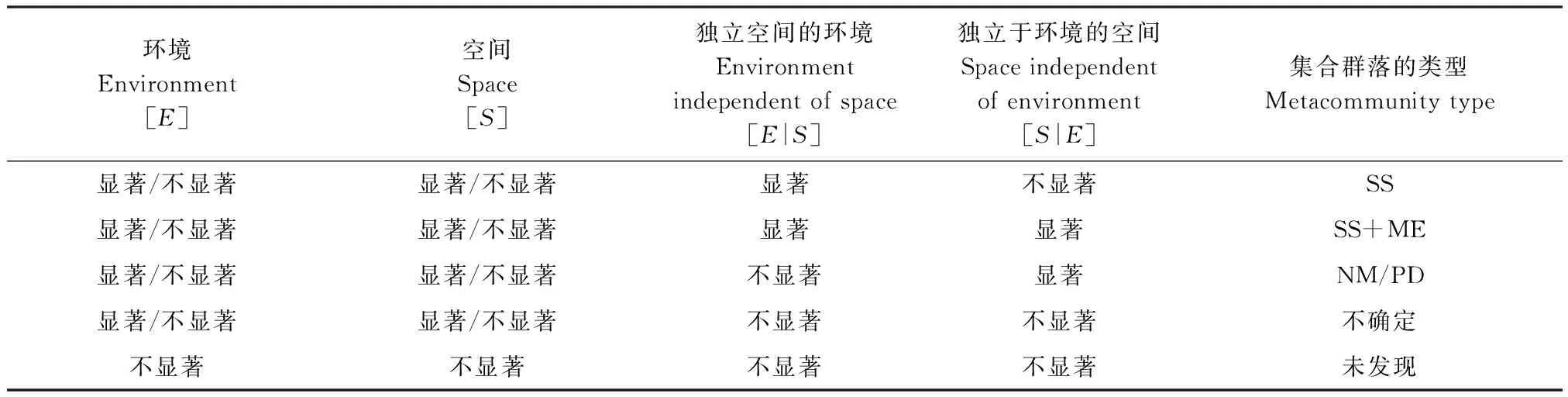
表1 显著性结构和集合群落类型关系的判断树
2 结果与分析
2.1 啮齿动物物种组成
2010—2012年7月,8个局域群落中共布放活捕笼5376笼日,共捕获啮齿动物3科6属6种(图2),其中仓鼠科(Cricetidae)3种,包括:子午沙鼠(Merionesmeridianus),长爪沙鼠(Merionesunguiculatus),小毛足鼠(Phodopusroborovskii);跳鼠科(Dipodidae)2种,包括:三趾跳鼠(Dipussagitta),五趾跳鼠(Allactagasibirica);松鼠科(Sciuridae)1种为阿拉善黄鼠(Spermophilusalaschanicus)。图2为该地区各鼠种捕获量比例,子午沙鼠的捕获量比例最高,为31.70%;长爪沙鼠最低,为6.24%。

图2 啮齿动物物种组成Fig. 2 Rank of rodent species composition
表2中不同样地之间对应的数字为扩散个体的数量,例如S1与S2之间的扩散个体数为7,S1与S3之间的扩散个体数为2。Hubbel定义集合群落为通过多物种的扩散连接局域群落[7]。本研究中在不同的生境斑块中监测到子午沙鼠、三趾跳鼠、五趾跳鼠和小毛足鼠等4个鼠种的38个个体的实际扩散(表2),表明该区域啮齿动物群落是由多个物种扩散而连接的局域群落而组成。为了进一步了解该区域啮齿动物是否构成集合群落,并且判断由集合群落4个范式中的哪一个所决定,进行了冗余分析和偏冗余分析。
2.2 冗余分析与偏冗余分析
由表3 可知,DCA第一排序轴长为2.072,排序轴的长度小于3,因此基于线性模型的冗余分析 (RDA) 适用于本研究。由表4 可知RDA前4个排序轴的信息,解释变量[E]、[S]、[E|S]、[S|E]与物种的前两个排序轴的方差累计贡献率均达100%,[E+S] 与物种的前两个排序轴的方差累计贡献率为95.8%,表明各解释变量的前两个排序轴完全可以进行解释。[E]、[S]、[E|S]、[S|E]、[E+S]物种的前4个排序轴的方差累计贡献率分别为97.7、97.3、86.5、98.9、97.5, 表明各解释变量的前4个排序轴可以进行解释。
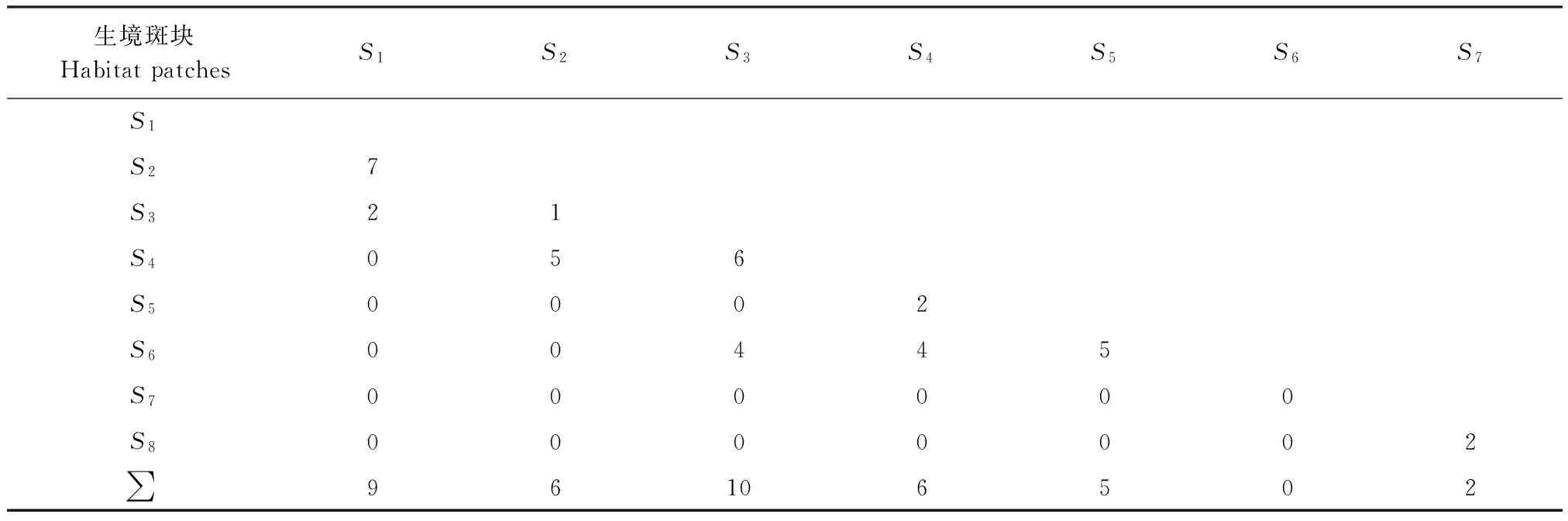
表2 不同生境斑块中个体扩散数量

表3 DCA排序轴的特征值和轴长
冗余分析排序结果表明(图3,表5),不同的生境中啮齿动物分布不同(图3)。由表5可知,环境特征独自解释72.8%的啮齿动物物种组成变化,环境因子中梭梭生物量与草本植物盖度箭头连线最长,对啮齿动物物种组成有显著影响 (图3)。空间特征独自的解释33.8%的物种组成变化,空间轴与啮齿动物物种的关系较弱于环境轴与啮齿动物物种的关系,空间轴箭头连线短于环境轴(图3)。环境特征和空间特征共同解释86.5%的啮齿动物物种组成变化,结果显著(P= 0.032)。去除环境特征之后,空间特征解释13.7%的变化(P= 0.246),结果不显著。去除空间特征之后,环境特征解释52.7%的变化(P= 0.016)。环境特征和空间特征的交互作用解释20.1%的物种组成的变化。以样地中扩散个体数量为因变量,以样地之间的距离为自变量做冗余分析,距离能够解释52.9%的扩散个体数量的变化(P= 0.002),说明距离显著影响扩散。纯环境变量对物种组成的变化作用显著,纯空间作用不显著,根据表1能够判断该区域啮齿动物构成集合群落,并由物种分配理论解释集合群落的结构。
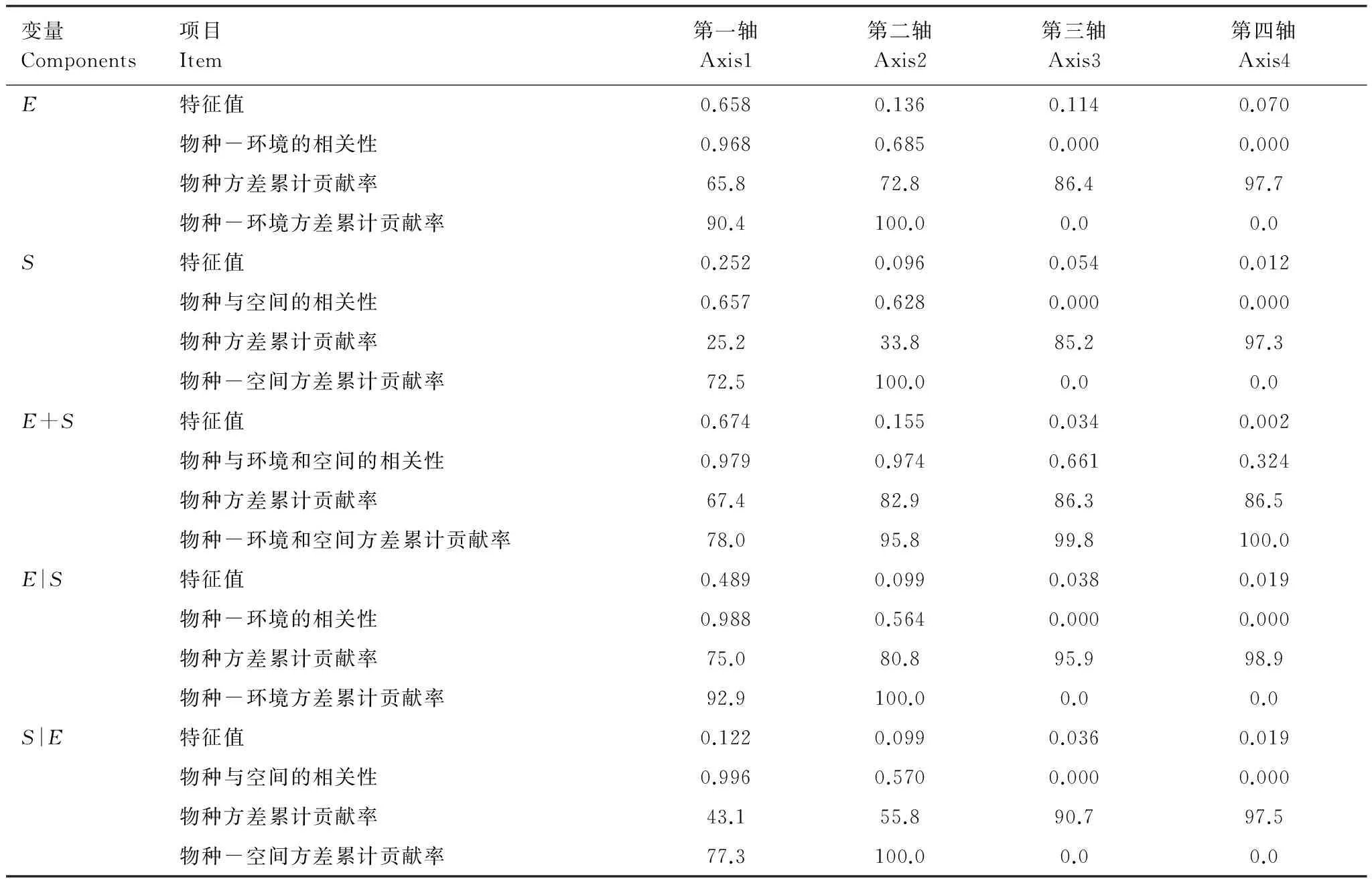
表4 解释变量冗余分析及偏冗余分析结果
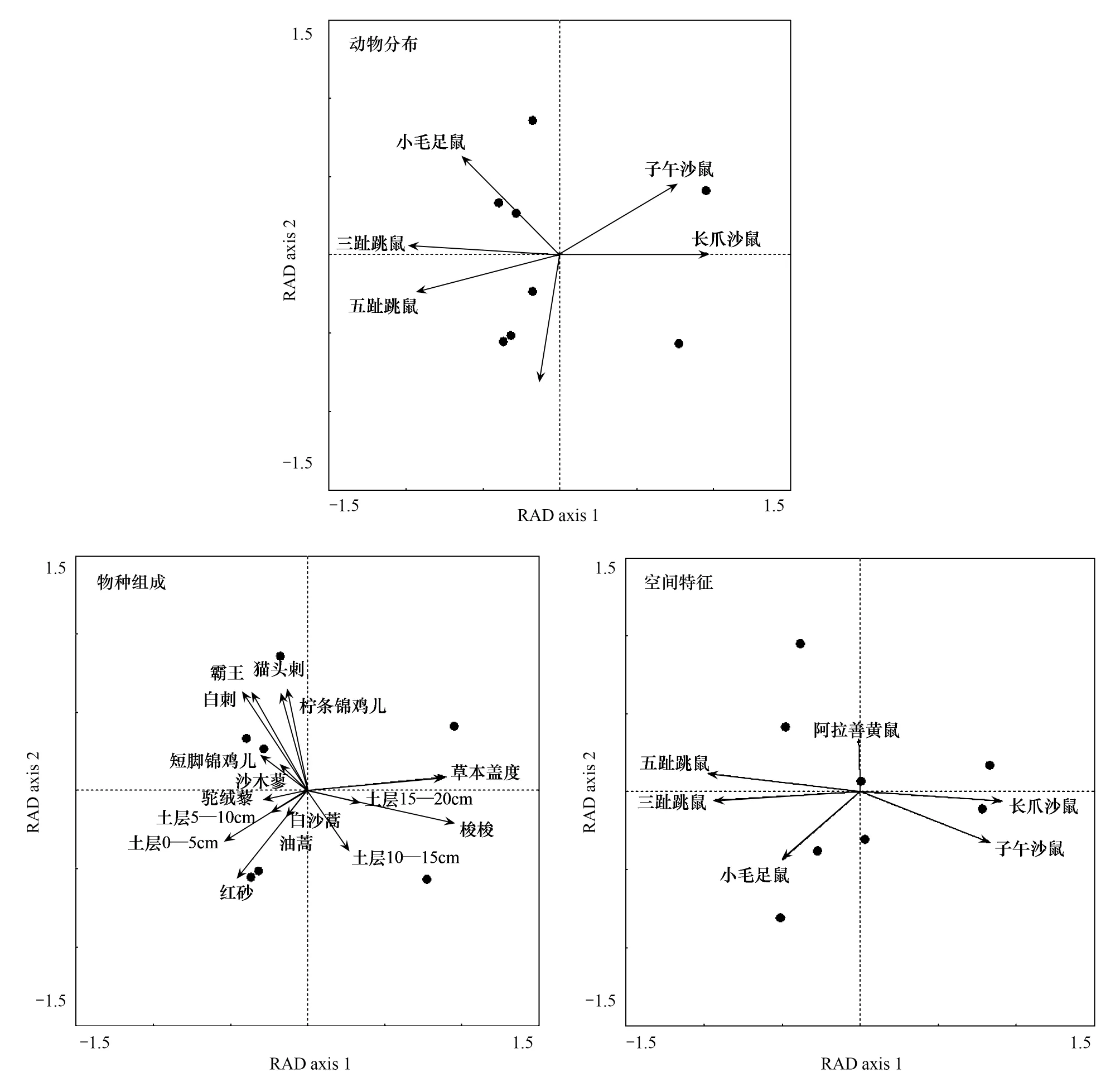
图3 环境和空间变量解释啮齿动物物种组成的冗余分析Fig.3 Results from redundancy analyses using environmental and spatial predictors to account for rodent species composition across the metacommunity

表5 冗余分析与偏冗余分析各变异的解释结果
3 讨论
集合群落现已成为斑块化生境中生物群落中普遍存在的群落结构。集合群落的思想是在集合种群的理论框架基础上发展起来的,也是群落格局和动态的重要理论基础之一,并已成为近些年来日益受到关注的生态学研究热点[31]。自Leibold等提出集合群落结构(物种共存)的4个范式(物种分配、集团效应、斑块动态和中性理论)以来得到了世界上群落生态学家广泛的重视和认同,由此兴起了日益高涨的有关集合群落研究。学者们以此理论框架为基础,针对不同的研究对象,在不同的生态局域或区域尺度上探求理论上的模拟模型和实践上的真实性。当今国际上有关水体动物集合群落集中了较多的研究[14- 16],特别是以Cottenie为代表的研究者们对分布于岛屿上界限分割明显,且有溪流相连接的池塘中的浮游生物集合群落进行了较为深入的研究,目的是为了寻找池塘复杂水体中浮游动物集合群落结构直接观察到的证据,并确定浮游动物群落结构变差中局域和区域(空间)过程的贡献。研究结果表明,在这样高度相连接的池塘中局域环境限制能够足以构成局域群落。理论模拟模型研究从群落组成物种的竞争、扩散、绝灭、物种周转、生物多样性等多方向开展了研究,研究结果可谓五花八门[8- 11]。脊椎动物集合群落的研究相对较少,往往是采用大尺度研究[21, 23](如国家尺度),以揭示区域过程(空间变化)在脊椎动物集合群落结构和动态的作用。如Driscoll 和Lindenmayer 研究鸟类和爬行类集合群落理论的实验检验,发现不同的集合群落机制作用短暂,在不同的区域中对于不同的分类组有时同时起作用,这种反应的复杂性意味着集合群落的概念仍然不能用于管理上的预测[22]。就当今有关集合群落的理论和野外实验研究结果看,Leibold等提出的集合群落物种共存的四个范式仍然不是一种普适的定律,由于研究对象和尺度的不同,产生不同的研究结果。
荒漠啮齿动物群落对于了解局域生物的相互作用和环境条件在局域多样性上的效应代表了一个独特的系统。在区域水平上几乎没有研究检验过大量的群落,而且对于了解群落的区域段的结构没有假设的集合群落范式(观念)[24]。有关啮齿动物集合群落研究目前缺乏,可能是3种原因导致了这种现象。一是方法上的,简单来说,从集合群落方法而来的许多思想是理论上的,而且这样的创造很容易应用到小尺度中型实验生态系统的实验中;第二是已经反复证明了了解局域控制的这些简单系统仍是粗糙系统,而且了解的局域过程合成的假设,对于了解这些群落的结构是充分的;第三是哺乳动物群落在空间上分布广阔,并且扩散常常充分隐含而且难以监测[24]。
选择阿拉善荒漠人为干扰下破碎化生境中的啮齿动物群落,在区域水平上研究荒漠啮齿动物集合群落的特征。在长期定位研究和取样的基础上,分析多个局域群落(样地)由扩散联系的集合群落在区域过程(空间变化)中的特征,检验Leibold 等提出的集合群落结构的4个范式在真实群落中的真伪及其哪个作用是主要的,目的是为集合群落理论的发展提供野外真实群落的实证,并完善该理论。研究结果显示物种共存中环境特征起着主导作用,实验证实物种分配模式解释当地集合群落的结构。Cottenie为了阐述(检验)集合群落特征与局域环境和区域空间过程之间的相关性,收集了有关集合群落结构的158个公开发表的数据集,采用环境和空间变量数据,对其标准化处理之后进行了整体的分析,结果表明大约50%的群落组成的变化被环境和空间变量解释。大多数数据集由物种分配模式(SS)构成,其次是物种分配模式和集团效应的结合(SS+ME)[28]。Meynard和Quinn在智利温带森林,研究了环境梯度和空间结构对鸟类集群的影响,他们认为群落组成和多样性的空间结构较弱,多样性受环境的强烈影响,SS在驱动鸟类的集群结构中起着主导作用。在其研究的尺度上,集合群落理论对总结该区域中驱动鸟类的集群机制提供了新的思路[21]。研究与温带森林鸟类的研究都验证了SS。研究结果与Steven 和Tello 对莫哈韦荒漠啮齿动物物种组成的模式相比较分析,Steven 和 Tello研究取样的尺度更大,空间结构显著,由此可知研究野外真实集合群落的共存机制,即Leibold 等提出的4种范式,取样的尺度仍然是影响哪种范式决定集合群落结构的主要因素。因此,对于荒漠啮齿动物集合群落的检验,确定Leibold等提出的四种范式中的哪一种范式主要决定集合群落结构,采用不同取样尺度进行比较分析是非常必要的。
在传统的群落生态学中,生态位理论解释群落结构特征模式[32-33],然而物种的空间分布和群落的结构是种群统计学、行为特征以及物种在空间上异质环境条件下相互作用的结果,因而物种集群是一种综合的反应变化。生态位理论是物种分配集合群落概念的基础[5],也是在自然界最常见的过程[28]。以生态位理论解释群落结构物种的共存机制只是从群落组成种内在的关系上探求原因。付和平等对该地区啮齿动物的空间生态位及其季节特征进行了研究,结果表明在荒漠生境中,啮齿动物对资源利用存在较大程度的趋同性,生态位重叠和生态位分化随着不同生态季节的变化发生明显的改变[34]。有些研究表明在一个群落中某物种的存在和物种的丰富度是由环境或栖息地特征决定的[35- 37]。通过物种集群空间上的变化分析有助于推测形成生态群落外形的过程。群落生态学家面临着描绘格局和过程清晰连接的挑战[38]。最近Mclntire和 Fajardo提出一种概念框架,通过分析空间格局或者空间残留以强化生态学中利用空间作为揭示没有测过的和不能测试的生态过程的替代物[39]。因而对于集合群落的空间分析是完善和修正Leibold等提出的四个范式的必然要求。
野外研究时间段虽然从每年4月到10月,但只选取了7月份的啮齿动物物种组成作为分析变量。一是在该环境中连续多年的取样,分析发现7月份啮齿动物组成的物种数和植物物种的多样性最丰富,代表了该区域荒漠生态系统中啮齿动物和植物物种变化的最丰富阶段[40-41]。二是当地的农事活动和放牧强度最大,是人为干扰最为强烈的时间段,能够较好的揭示人为干扰下荒漠啮齿动物集合群落共存的特征。当今集合群落范式对于群落的空间的动态有了较多的实证[23-24, 28],但几乎没有研究绝对调查时间尺度在范式上的重要性。因此,将时间尺度动态整合到集合群落范式中是未来集合群落生态发展中的重要方向,也是未来研究的重点。
[1] Brown J H, Fox B J, Kelt D A. Assembly rules: desert rodent communities are structured at scales from local to continental. The American Naturalist, 2000, 156(3): 314- 321.
[2] López- González C, Presley S J, Lozano A, Stevens R D, Higgins C L. Metacommunity analysis of Mexican bats: environmentally mediated structure in an area of high geographic and environmental complexity. Journal of Biogeography, 2012, 39(1): 177- 192.
[3] Ricklefs R E. Community diversity: relative roles of local and regional processes. Science, 1987, 235(4785): 167- 171.
[4] Ng I S Y, Carr C M, Cottenie K. Hierarchical zooplankton metacommunities: distinguishing between high and limiting dispersal mechanisms. Hydrobiologia, 2009, 619(1): 133- 143.
[5] Leibold M A, Miller T E. From Metapopulations to Metacommunities. Ecology, Genetics, and Evolution of Metapopulations. Elsevier Academic Press, 2004:102-220.
[6] Chesson P. Mechanisms of maintenance of species diversity. Annual Review of Ecology and Systematics, 2000, 31: 343- 366.
[7] Hubbell S P. The Unified Neutral Theory of Biodiversity and Biogeography. Princeton, NJ: Princeton University Press, 2001:78-152.
[8] Chase J M. Towards a really unified theory for metacommunities. Functional Ecology, 2005, 19(1): 182- 186.
[9] Calcagno V, Mouquet N, Jarne P, David P. Coexistence in a metacommunity: the competition-colonization trade- off is not dead. Ecology Letters, 2006, 9(8): 897- 907.
[10] Mouquet N, Miller T E, Daufresne T, Kneitel J M. Consequences of varying regional heterogeneity in source-sink metacommunities. Oikos, 2006, 113(3): 481- 488.
[11] Koelle K, Vandermeer J. Dispersal- induced desynchronization: from metapopulations to metacommunities. Ecology Letters, 2005, 8(2): 167- 175.
[12] Venail P A, MacLean R C, Bouvier T, Brockurst M A. Diversity and productivity peak at intermediate dispersal rate in evolving metacommunities. Nature, 2008, 452(7184): 210- 214.
[13] Mouquet N, Loreau M. Coexistence in metacommunities: the regional similarity hypothesis. The American Naturalist, 2002, 159(4): 420- 426.
[14] Cottenie K, Michels E, Nuytten N, De Meester L. Zooplankton metacommunity structure: regional vs. local processes in highly interconnected ponds. Ecology, 2003, 84(4): 991- 1000.
[15] Cottenie K, De Meester L. Connectivity and cladoceran species richness in a metacommunity of shallow lakes. Freshwater Biology, 2003, 48(5): 823- 832.
[16] Cottenie K, De Meester L. Metacommunity structure: synergy of biotic interactions as selective agents and dispersal as fuel. Ecology, 2004, 85(1): 114- 119.
[17] Noda T. Metacommunity- level coexistence mechanisms in rocky intertidal sessile assemblages based on a new empirical synthesis. Population Ecology, 2009, 51(1): 41- 55.
[18] Heino J. Metacommunity patterns of highly diverse stream midges: gradients, chequerboards, and nestedness, or is there only randomness? Ecological Entomology, 2005, 30(5): 590- 599.
[19] Hylander K, Nilsson C, Jonsson B G, Göthner T. Differences in habitat quality explain nestedness in a land snail meta- community. Oikos, 2005, 108(2): 351- 361.
[20] Brooks D R, Perry J N, Clark S J, Heard M S, Fribank L G, Holdgate R, Mason N S, Shortall C R, Skellern M P, Woiwod L P. National- scale metacommunity dynamics of carabid beetles in UK farmland. Journal of Animal Ecology, 2008, 77(2): 265- 274.
[21] Meynard C N, Quinn J F. Bird metacommunities in temperate South American forest: vegetation structure, area, and climate effects. Ecology, 2008, 89(4): 981- 990.
[22] Driscoll D A, Lindenmayer D B. Empirical tests of metacommunity theory using an isolation gradient. Ecological Monographs, 2009, 79(3): 485- 501.
[23] Presley S J, Higgins C L, López- González C, Stevens R D. Elements of metacommunity structure of Paraguayan bats: multiple gradients require analysis of multiple ordination axes. Oecologia, 2009, 160(4): 781- 793.
[24] Stevens R D, Tello J S. Do desert rodents form metacommunities? Journal of Mammalogy, 2012, 93(4): 1029- 1041.
[25] Ebenman B, Law R, Borrvall C. Community viability analysis: the response of ecological communities to species loss. Ecology, 2004, 85(9): 2591- 2600.
[26] Jongman R H, Ter Braak C J F, Van Tongeren O F R. Data Analysis in Community and Landscape Ecology. Cambridge: Cambridge University Press, 1995: 144- 147.
[27] Borcard D, Legendre P, Drapeau P. Partialling out the spatial component of ecological variation. Ecology, 1992, 73(3): 1045- 1055.
[28] Cottenie K. Integrating environmental and spatial processes in ecological community dynamics. Ecology Letters, 2005, 8(11): 1175- 1182.
[29] Chase J M, Leibold M A. Ecological Niches. Chicago: University of Chicago Press, 2003.
[30] Yu D W, Wilson H B, Pierce N E. An empirical model of species coexistence in a spatially structured environment. Ecology, 2001, 82(6): 1761- 1771.
[31] Wilson D S. Complex interactions in metacommunities, with implications for biodiversity and higher levels of selection. Ecology, 1992, 73(6): 1984- 2000.
[32] Hutchinson G E. Concluding remarks. Cold Spring Harbor Symposium on Quantitative Biology, 1957, 22: 415- 427.
[33] Leibold M. A. The niche concept revisited: mechanistic models and community context. Ecology, 1995, 76(5): 1371- 1382.
[34] 付和平, 武晓东, 杨泽龙. 不同干扰条件下荒漠啮齿动物生态位特征, 生态学报, 2005, 10(25): 2637-2643.
[35] Wellborn G A, Skelly D K, Werner E E. Mechanisms creating community structure across a freshwater habitat gradient. Annual Review of Ecology and Systematics, 1996(27): 337- 363.
[36] Gilbert B, Lechowicz M J. Neutrality, niches, and dispersal in a temperate forest understory. Proceedings of the National Academy of Sciences of the United States of America, 2004, 101(20): 7651- 7656.
[37] Parris K M, Lindenmayer D B. Evidence that creation of a Pinus radiata plantation in south- eastern Australia has reduced habitat for frogs. Acta Oecologica, 2004, 25(1- 2): 93- 101.
[38] Vellend M. Conceptual synthesis in community ecology. The Quarterly Review of Biology, 2010, 85(2): 183- 206.
[39] McIntire E J B, Fajardo A. Beyond description: the active and effective way to infer processes from spatial patterns. Ecology, 2009, 90(1): 46- 56.
[40] 张福顺. 不同干扰条件下荒漠啮齿动物种群和群落动态研究 [D]. 呼和浩特: 内蒙古农业大学, 2011.
[41] 阿娟, 张福顺, 张晓东, 武晓东. 荒漠植物群落特征及其与气候因子的对应分析. 干旱区资源与环境, 2012, 26(1): 174-178.
An empirical research of rodent metacommunities in Alashan Desert
CHA Muha1,2, WU Xiaodong1,2,*, FU Heping1,2, YUAN Shuai1,2, WU Yunga1,2, ZHANG Xiaodong3
1CollegeofEcologyandEnvironmentalscience,InnerMongoliaAgriculturalUniversity,Hohhot010019,China
2KeyLaboratoryofPratacultureandGrasslandResources,MinistryofEducation,Hohhot010019,China
3InnerMongoliaForestryMonitoringandPlanningInstitute,Hohhot010020,China
Introduction of the metacommunity concept has made a substantial contribution to better understanding of community composition and dynamics of a fragmentized habitat at a regional level. Leibold and Miller defined metacommunity as a set of local communities that are linked by dispersal of multiple potentially interacting species. The metacommunity concept has provided a new and revolutionary paradigm for the community ecology by emphasizing integrated variation among communities at the regional level. This variation is accentuated by biotic interactions and spatial variation in environmental characteristics and modulated by dispersal among sites. The basic problems of metacommunity research are the multiple species coexistence mechanism, the reasons behind the diversity, and mechanism maintaining this diversity within the same system. Leibold and Miller proposed four paradigms for metacommunities: the patch-dynamic view, the species-sorting view, the mass effects view, and the neutral view, each emphasizing different processes of potential importance in metacommunities. A variety of studies verified the four ecological paradigms, but only few addressed the terrestrial vertebrates system. In recent years, as a result of human activities and natural disturbances, Alashan Desert habitat in Inner Mongolia has been largely fragmentized. This habitat fragmentation is a major threat to animal communities. From 2010 to 2012, we investigate rodent communities and habitat environmental factors at eight fixed experimental sites in Inner Mongolia′s Alashan Desert. We used a set of full and partial redundancy analyses to estimate four additive partitions of variance in rodent species composition: (a) unexplained variation, (b) variation explained by spatial characteristics, (c) variation explained by local environmental conditions, and (d) variation explained by both spatial and environmental characteristics. The results of full and partial redundancy analyses showed that habitat and spatial characteristics alone accounted for 72.8% and 33.8%, respectively, of the variation among sites regarding rodent species composition. All explained variables accounted for approximately 86.5% of the variation (P= 0.032) in rodent species composition among sites. Spatial characteristics accounted for 13.7% of the variation (P= 0.246) in species composition after removing variation shared with habitat characteristics. Habitat characteristics accounted for 52.7% (P= 0.016) of the variation in species composition after removing variation shared with spatial characteristics effect. Finally, spatial structured habitat characteristics (interaction term) accounted for approximately 20.1% of the total variation in rodent species composition among sites. The environmental component to rodent species composition was greater than the spatial component. Pure environmental characteristics accounted for significant amounts of variation in rodent species composition, and pure spatial characteristics accounted for non-significant amounts of variation in rodent species composition. The results from variance decomposition suggested that the species sorting model of metacommunity structure fits better the rodent community than the mass effect or neutral models.
community ecology; rodent; metacommunity; redundancy analysis
国家自然科学基金资助项目(30760044, 31160096);公益性行业科研专项经费项目(201203041)
2013- 12- 09;
日期:2015- 04- 24
10.5846/stxb201312092913
*通讯作者Corresponding author.E-mail: wuxiaodong_hgb@163.com
查木哈,武晓东,付和平,袁帅,乌云嘎,张晓东.阿拉善荒漠啮齿动物集合群落实证研究.生态学报,2015,35(17):5612- 5622.
Cha M H, Wu X D, Fu H P, Yuan S, Wu Y G, Zhang X D.An empirical research of rodent metacommunities in Alashan Desert.Acta Ecologica Sinica,2015,35(17):5612- 5622.
