陆地生态系统植物多样性对矿质元素输入的响应
2015-01-18毛庆功鲁显楷莫江明
毛庆功, 鲁显楷, 陈 浩, 莫江明
1 中国科学院华南植物园, 中国科学院退化生态系统植被恢复与管理重点实验室, 广州 510650 2 中国科学院大学, 北京 100039
陆地生态系统植物多样性对矿质元素输入的响应
毛庆功1,2, 鲁显楷1,*, 陈 浩1,2, 莫江明1
1 中国科学院华南植物园, 中国科学院退化生态系统植被恢复与管理重点实验室, 广州 510650 2 中国科学院大学, 北京 100039
人类活动的加剧改变了陆地生态系统矿质元素(如氮、磷、钾等)循环的速度和方向,并且对生态系统的结构和功能也产生重要影响。如今,矿质元素输入量的改变及其产生的后续效应对陆地生态系统生物多样性的影响备受学者们的关注。从4个方面综述了全球氮沉降背景下主要矿质元素输入的改变对陆地植物多样性的影响及其机理:1)矿质营养元素限制的概念、确定方法以及与植物多样性的耦合关系;2)概述了氮、磷、钾等主要矿质元素输入对陆地植物多样性的影响:主要表现为负面效应;3)探讨了矿质元素输入影响植物多样性的可能机制,包括生态系统水平上的机制(如竞争排斥、酸化铝毒、物种入侵、同质性假说,间接诱导机制等)和植物个体水平上的机制(如元素失衡和环境敏感性增加等);4)根据目前研究现状,指出了已有研究的局限性,分析了未来可能的研究方向和重点。
矿质元素; 养分限制; 氮沉降; 生物多样性; 陆地生态系统; 全球变化
人类活动所带来的环境问题日益加剧,已经严重威胁到生物多样性[1],其中包括栖息地的丧失,外来物种入侵和环境污染等[2]。如果这种趋势保持不变,预计到21世纪末将有20%—50% 的物种不可逆转地消失[3]。物种多样性减少将导致生态系统结构和功能的衰退[4-5],并直接威胁到人类福祉。因此迫切需要了解多样性变化的原因及保护对策。
近几十年陆地植物多样性正遭受严重威胁。其中一个重要原因就是矿质元素供应的平衡被打破,即氮(N)、磷(P)、钾(K)、钙(Ca)、镁(Mg)甚至微量元素和二氧化碳(CO2)等供应失衡,进而对植物的生长和群落组成产生一系列影响。特别是那些对植物的生长起关键作用并且易受人类活动影响的矿质元素(如N,P等),它们既影响植物的生长又与其他代谢因子(如水和CO2等)共同作用调节植被的组成。矿质元素供应失衡通常由某些元素(如N)输入过量和/或养分元素(如Ca、Mg)大量流失导致。N沉降,土地利用和CO2剧增等全球变化因子都直接或间接地参与了这一过程。由于N素是大多数陆地生态系统植物生长的限制因子,N沉降引起的生物多样性降低备受关注。目前,大气N沉降被认为是威胁本世纪全球生物多样性的第三大因素(继土地利用变化和全球气候变化之后)[6-7]。在工业发展较早的欧美地区,高N沉降对森林生态系统造成的影响(如土壤酸化、养分失衡、生产力降低、森林衰退等)已严重威胁到当地生物多样性,并引起学术界和民众的广泛关注[8-9]。即使是远离人类活动密集的地区,由于受到大气传输及元素循环特征的作用,也同样面临N沉降升高的问题。据估计,34个陆地生物多样性热点地区(包含全球一半的植物种类),在20世纪90年代的N沉降水平是世界陆地平均水平的1.5倍,到2050年将会超过2倍[10],其中近半数地区的N沉降将超过敏感系统响应的临界值(5—10 kg N hm-2a-1)[9]。
由于各种矿质元素之间相互耦合的关系,一种元素输入格局的改变势必影响其它元素的循环。例如,N沉降的增加可能加剧植物受P及盐基离子(如K,Ca,Mg等元素)供应的限制,对植物生长及多样性维持带来一系列负面影响[11- 14]。在N沉降背景下,研究者们为了深入探讨N沉降增加对植物多样性影响的机理和植物的生存机制,开展了一系列的矿质元素添加实验。这些研究有助于我们更好地认识元素循环的变化对陆地植物多样性的影响及其机理,从而为生态系统管理和可持续发展提供理论依据。
我国是世界上生物多样性最为丰富的国家之一。当前中国在经济高速发展的背景下N沉降呈现快速增长,对生态系统的结构和功能构成的威胁也日益严重[15-16]。在中国南方一些森林,仅降水中N沉降已高达30—73 kg N hm-2a-1[17-18],与欧洲高N沉降速率相当[19]。随着人口的增长和工农业的高速发展,我国的N沉降量将会继续增加[15]。研究矿质元素动态变化对生物多样性的影响及其机制已经成为亟待解决的重要课题。目前,国内有关矿质元素添加对植物多样性变化的相关综述极少,仅有少数关于N沉降对草原和森林生态系统生物多样性影响的综合报道[20-21]。对于N以外的其他元素添加的状况缺乏全面清晰的总结,对于机理的认识还存在很大欠缺。因此,本文综述了主要矿质元素输入对陆地植物多样性的影响,并总结了植物可能的响应与适应机制,以期为制定生态系统可持续发展的策略服务。
1 矿质元素限制的概念、确定方法及与元素与多样性的耦合关系
1.1 元素限制的概念
元素限制的概念最初来自农业,是指由于必要元素供应不足,限制某一株植物或某种植物生产力的增长。因此,矿质元素限制意味着,当植物的生长受到某一必需元素的限制时,它的生物量生产将会低于这种元素供应充足的情况[22]。单株植物或群落越受到养分元素的限制,生物量增量对限制元素添加的响应越敏感[23]。
1.2 元素限制的确定方法
确定植物元素限制的直接方法是通过向系统添加矿质元素,监测其生物量变化状况。但是这种直接方法的应用性较弱,因此多数研究是会通过间接方法反映生态系统元素限制情况。Koerselman[24]的研究通过综述40个施肥实验发现植被N∶P可以很好的判断该地区元素限制状况。作为生态化学计量学方法其原理是在正常情况下,植物体内的元素比在一定范围内相对恒定,外部元素的动态状况会在植物体内元素比上体现。也有学者利用特定群落中敏感物种的相对元素含量来指示特定土壤元素限制状况[25]。此外,一些在元素循环过程中起关键作用的酶的活性也间接表明了植物元素限制的状况,如土壤磷酸酶的活性可以间接反映P的限制状况[26]。在实际应用中,判断元素限制的可靠方法是根据直接和间接证据共同验证,以确定元素限制的类型和程度。
1.3 元素与多样性的耦合关系
物种多样性的存在依赖于特定的环境因子。在自然生态系统中,环境中的元素水平与多样性存在动态平衡(通常是系统养分受限)。理论上认为,当元素添加刺激生产力的增加超过一定界限时群落多样性将会下降,即打破了元素与多样性之间的平衡[27-28]。最近几年的一些长期研究也证明随着时间的推移,不同物种间的适应性差异将表现出竞争关系和多样性的变化,往往导致高养分下不占竞争优势的物种(如稀有物种,多年生物种和固氮物种)消失[29- 31]。反过来,多样性的变化亦会影响到元素的状态。多样性降低使群落中的物种不能充分利用生态位,从而降低生态系统功能的多样性[32],最终影响元素循环速率等多种生态过程[33]。
2 矿质元素输入对植物多样性的影响
在植物多样性与矿质元素输入的研究中,N和/或P添加是最为常见的。从总体上,元素添加的研究多集中在温带地区(表1)[34-75]。就响应的敏感性而言,不同功能群植物大致为:苔藓和地衣>草本>灌木>乔木。
2.1 N素输入对植物多样性的影响
在各类陆地生态系统中,多数研究表明N添加改变了群落物种组成(表1)。即便是长期低水平的N添加量,也有可能使植被多样性发生显著性改变[7]。为此欧美学者为不断完善N沉降临界负荷的概念和标准做了大量工作[76],并且已经为欧美一些发达国家制定策略以降低对敏感生态系统的N沉降量提供参考依据[7, 76]。
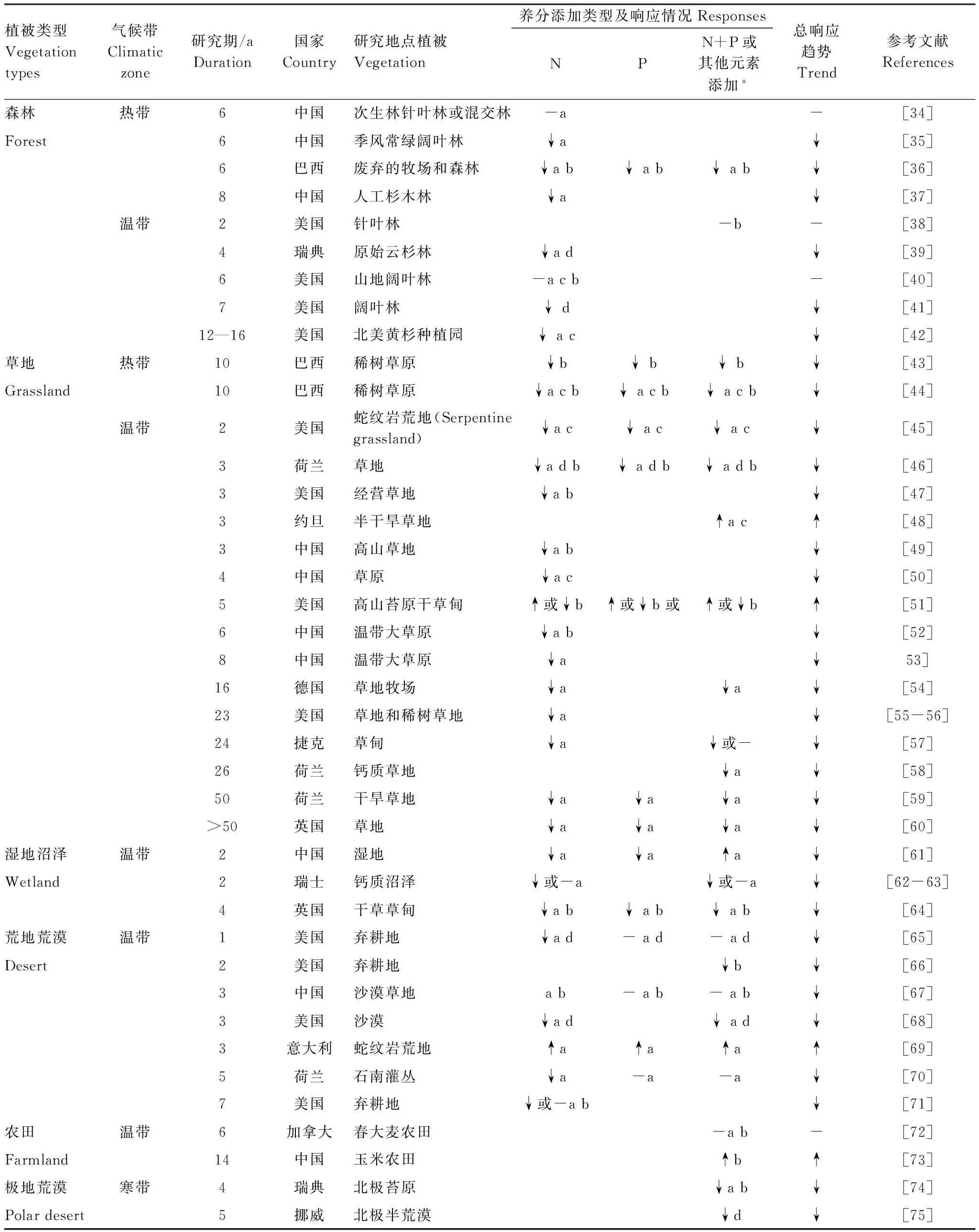
表1 典型陆地生态系统植物多样性对矿质元素添加的响应
在森林生态系统,林下层植物对养分添加的响应比林冠层植物要敏感的多[21]。N添加降低林下层植物多样性,这在温带地区已经被欧美学者多次证实,如在美国Harvard红松(Pinusresinosa)林,Rainey等[77]发现草本层植物的密度和生物量在7a N添加后分别降低了80%和90%。在热带地区,鲁显楷等[35]在鼎湖山季风常绿阔叶林的研究发现,高N添加(150 kg N hm-2a-1)显著地降低了林下层植物的丰富度,该研究同时指出,未来N沉降增加可能威胁热带亚热带“富N”生态系统的植物多样性。此外值得注意的是,国际上对北温带松林的研究普遍发现,长期N沉降通常不利于针叶树种的正常生长。长期N沉降使松针类树种出现生产长减缓,死亡率升高等现象[9, 78-79]。这意味着在陆地上大面积以松树为优势种的森林生态系统在全球N沉降背景下可能面临严峻威胁。
草地生态系统上最具代表性的研究是在美国著名的明尼苏达州开展的草地多样性试验,该研究发现:与对照相比(仅接受大气N沉降),长期低水平N添加(10 kg N hm-2a-1)使植物物种数降低了17%[56];另外,N素添加还改变了物种组成,使该地以C4暖季草(Schizachyriumscoparium)为优势的群落转为多样性较低的混合C3冷季草(Agropyronrepens)群落[55]。而且该实验最近的研究表明,长期高N添加严重降低了植物多样性,即使停止施N 素20a后多样性仍然不能恢复到对照组的水平[80],这说明多样性的降低很可能是不可逆的。在国内,白永飞等[50]通过梯度N素添加(17.5、52.5、105、175、280 kg N hm-2a-1)研究了我国内蒙古成熟和退化草原植物多样性的变化对N沉降的响应,发现N添加显著降低了成熟草原的物种丰富度。我国西部高山草地、高寒草甸、荒漠化草原、沙地草原等野外N添加实验也大多表现出多样性降低的趋势[49, 81- 83]。湿地和荒漠地区的N添加对原有物种生物量和多样性的作用也几乎都是负面的, 但是对禾草和外来物种生产力起正面作用,而且作用的速度相对更快。此外,还有研究发现N增加尤其危胁到一些草地稀有濒危物种的生存[29, 31]。
但是也有研究表明,N的输入并没有明显影响群落的组成。如Gilliam等[84]对美国弗吉尼亚西部的Fernow实验森林研究表明,3a加N试验后(35 kg N hm-2a-1)草本层的群落并没有明显的变化。随后通过季节和年际尺度加以检验,依旧得出了相同结论[40]。因此,N添加是否改变多样性,还要看N添加的种类,数量,持续时间和生态系统类型。
2.2 P素输入对植物多样性的影响
尽管人类活动已经在自然背景值上将P的通量提高了3倍,但是由于P的循环特征,陆地生态系统,特别是低纬度的热带地区,表现为P限制[12, 22]。
相比于N素,P在土壤中本身较为稳定,植物可能更容易对P可用性增加的状况作出响应,因此在P和N作用于多样性的大小关系上,P的作用可能更为关键。一方面,有长期研究表明,相比N添加,P添加对植物群落的影响具有遗留效应(Legacy-effects)[58]。另一方面,相对于N添加研究的数量,P添加的研究相对较少,对多样性变化的机理解释也较少。单独的P素添加研究较为少见,P添加多出现在与N,K,Ca等组合的实验设计中,以研究元素限制、元素平衡以及施肥效应等对群落组成的影响。多数研究认为P素输入增加对多样性的维持具有负面效应或无显著影响。如在森林系统[36],湿地生态系统[61],草地生态系统[64]等的研究中都有证据表明P添加降低物种丰富度或多样性。但是也有例外,P添加的作用可能还取决于群落结构和处理时间。如地中海矮灌丛群落(Mediterranean dwarf-shrub community),连续5a施加P(45, 90 kg P hm-2a-1)显著地增加了一年生豆科植物的生产力和物种丰富度,但对群落总丰富度并没有显著影响[85]。在荒漠荒地生态系统 P添加甚至提高了蛇纹岩荒地植物丰富度和地被盖度[69]。
近些年,不少研究发现P限制的条件对物种组成及高多样性的维持具有重要作用,虽然机理并不非常清楚,但是维持低P的环境无疑非常必要。Janssens等[86]集合欧洲5个国家281个站点的研究数据发现,最高的植物丰富度出现在P限制和K浓度适中的土壤上,在调查范围内高P浓度(>5 mg P/100 g干土)的区域均不存在高物种丰富度(>20种/100 m2)。同样在欧洲,Wassen等[87]进行的大尺度研究发现,稀有物种往往在P限制的地区具有更高丰富度。许多实验和调查研究以及模型分析的结果也支持P素增加比N素增加将对植物多样性产生更严重的威胁[88]。因此P可用性的变化相对于其他养分元素对植物丰富度可能更为重要。鉴于P的限制状况对维持生态系统物种多样性的重要作用,P可用性的增加可能会影响一些物种的生存,至少是威胁到那些稀有濒危物种的存在现状。
2.3 NP或NPK多元素同时输入对植物多样性的影响
多种矿质元素共同添加的实验可以研究养分动态对植物群落组成的影响并探索退化生态系统的恢复管理措施。但是植物多样性对NP或NPK添加的响应在不同研究中的结论并不一致。在少数元素添加量小,系统需肥量大及种间竞争尚不激烈的研究中,多元素添加有利于多样性的恢复和维持,但在大多数研究中多元素添加表现出多样性降低的趋势。NP共同添加可能起到两方面直接作用:一方面可以相互抵消由于NP单独添加带来的养分失衡。如Siddique等[36]在巴西亚马逊热带次生林的研究发现,N和P共同添加(100 kg N hm-2a-1, 50 kg P hm-2a-1)降低树木均匀度和物种自然恢复的速度,但小于单独添加N或P的作用。另一方面多元素添加可能共同加剧种间竞争。在英国的Park Grass 试验中,N、P共同添加导致了丰富度更为显著地降低[89]。Ren等[90]通过N、P、K(分别100、200、100 kg hm-2a-1)的添加研究了不同方式元素组合对高寒草甸植物群落物种丰富度和生产力的影响,发现随着元素组合数量的增加,植物多样性逐渐降低,当3种矿质元素共同添加时,植物丰富度降低的最为显著。同时,多元素添加所产生的多样性变化还有可能是元素添加带来的直接和间接效应共同作用的结果,其中间接效应多表现出元素添加对土壤化学性质和微生物的作用上。如Jacobson等[43]对巴西疏林草地乔木层的研究发现,添加NP(各100 kg hm-2a-1)8a后,伴随着土壤P和Al3+的增加以及pH值的下降,多样性显著降低。
钾(K)和钙(Ca)也是植物所必需的大量元素。然而K和Ca限制的情况很少发生,一般只有在低盐基饱和度或高水平酸沉降等异常干扰下才可能出现[24, 91]。因此K或Ca添加对物种组成一般不产生显著影响。在许多研究中,Ca添加并非以增加土壤养分元素为目的,而是通过含钙化合物添加(如CaCO3)控制土壤酸度和降低锰(Mn)、镍(Ni)等元素的可用性[69-70]。微量元素缺乏的现象并没有广泛性,因此微量元素添加的研究更多的是为了确保这些元素在整个研究过程不作为受限制的因子[50, 56]。除此之外,在大气CO2浓度升高背景下探究N、P添加效应的科学问题,国际上已开展了一些初步研究,但是对多样性的报道却非常有限,已有研究主要集中在草地生态系统。过去有研究发现CO2浓度升高可以降低植物体内的N浓度[92]。最近,Reich等[93]在温带草原的研究发现,同时作为植物养分的N和CO2单独增加均对多样性表现出负面影响,有趣的是当二者共同添加时,具有相抵消的交互作用,即N沉降增加的背景下增加CO2可以缓解由N沉降带来的负面效应。但这种交互作用是否在其他生态系统中也是一致的还有待进一步的研究。
3 矿质元素输入影响陆地植物多样性的机理
根据矿质元素添加对多样性动态的影响尺度,可以将植物多样性变化的机理分为两类:生态系统水平上的机理和植物个体水平上的机理。
3.1 生态系统水平上的机理
3.1.1 竞争排斥机制
国际上最早的长期矿质元素添加实验是开始于1856年英国洛桑实验站的 Park Grass Experiment[60]。而竞争机制一直成为温带(特别是草地)研究中解释养分效应引起多样性变化的主流理论。在具体的研究和争论过程中,不同学者提出了4种主要假说:1)总体竞争假设。2)地上竞争假设。3)地下竞争假设。4)密度自疏假设。
Grime[27]认为中等的养分条件下,将存在相对高的多样性;随养分元素可用性增加,竞争加剧,竞争力强的物种获得更高竞争优势,排除掉竞争力弱的物种,降低多样性。这被认为是不区分地上地下竞争的总体竞争假设或中度生产力假说。随后Newman[94]的评论认为多样性高的地方地下竞争同样强烈,并非地下竞争导致了多样性降低,从而提出植物地上竞争可能决定了多样性衰退。由于强调植物对光竞争所起到的重要作用,这种假设被认为是地上竞争假设或光竞争假设,并得到了许多实验证据[65, 95]和非对称生长理论的支持[96]。另一类假设则仅考虑个体竞争的结果,即群落水平上的密度自疏假设。最初由Oksanen[97]提出:养分的可用性与群落物种数存在钟形曲线的关系,但解释这种关系的变量可能是各个物种的个体数量而非群落的生物量。养分可用性超过临界值后群落中各植物种死亡率增加的程度趋于相同,但是这种密度自疏仅在养分相对肥沃的生境中导致物种丰富度降低。该假设提出后引来的质疑远多于支持。Rajaniemi等[98]认为密度的降低对解释种的损失十分有限,随机自疏,地上和地下竞争可能都降低了多样性。随后Rajaniemi等[66]通过人为控制地上和地下竞争混合种植下的施肥实验,发现在非光限制或人为干扰土地上的多样性取决于植物根之间的竞争,即地下竞争假设或根竞争假设。虽然以上4种假设具有不同的适用范围,但共同的一点是限制性养分添加后加剧了个体间的竞争。
此外,除了使某些植物获得个体数量和大小上的直接优势,养分添加还将通过冠层盖度,凋落物量,后代数量甚至化感作用的增加继续放大竞争排斥的间接作用。例如有研究表明N调控的凋落物量和后代植物的建成对群落丰富度有重要影响[53, 99-100]。
3.1.2 土壤酸化作用机制
植物生长对于土壤酸化非常敏感。大量研究表明在一定范围内植物丰富度与土壤酸度有一定的相关性[60-70, 89, 101-102],而过量的N输入可以导致土壤的酸化。有研究指出北方草地生态系统由酸化作用所引起的物种丰富度降低甚至大于养分增加导致的竞争排斥作用[31, 89]。在土壤酸化过程中,可交换阳离子Ca2+、Mg2+等加速流失,可溶性Al3+、Fe3+、Mn2+离子的浓度增加[13]。这些过量的Al3+可以通过伤害根系以及阻碍根系对Ca2+和Mg2+的吸收影响植物的养分平衡和正常生长[35, 70, 103-104],在酸性土壤上,植被的状况很大程度上取决于物种对于过量铝铁锰的耐受性[105]。N添加产生的酸化作用对多样性的不利影响在不同土壤类型下是有区别的,在酸性土壤上的作用可能更显著。Azevedo等[104]分析了全球140个不同研究中的2409个物种后发现,物种丰富度最高的地区虽然位于土壤酸度相对较低的热带森林(pH值 = 4.1),但是热带森林植物丰富度对土壤 小于4.1后的降低高度敏感。
3.1.3 物种入侵学机制
增加养分元素会加强外来物种侵入的风险[68, 106],从而威胁本土物种的多度和多样性。主要原因有两个方面:其一,本土物种对自身生存环境的长期适应,限制性元素的增加对本土物种的有利作用程度有限,甚至会产生不利作用;其二,外来物种具有更强的可塑性,元素可用性增加更有利于外来物种的侵入和种群壮大。对于一些本土物种,Bubier等[107]认为添加限制性元素虽然使该元素在植物体内积累,但却并没有更多地向光合机构分配。而对于入侵植物种,大量研究证实其成功入侵是由于对限制资源的利用效率高于本地物种。有研究发现我国的入侵植物紫茎泽兰(Ageratinaadenophora)和飞机草(Chromolaenaodorata) 就偏好N素较高的环境,在形态、生物量分配、生长和光合特性上对N具有良好的可塑性,土壤N含量升高利于紫茎泽兰和飞机草的入侵,且不受过高N水平的抑制[108]。入侵物种总是具备“理想杂草”的一些特性,使其可以在特定条件下获得竞争优势,例如在从苗期到繁殖期的高速生长,在适宜条件下产生更多繁殖体,由此威胁当地物种组成及生物多样性[109]。矿质元素输入改变了限制资源在物种间的分配格局,因此从长期效应来看,可能更有利于外来种入侵。许多研究发现,在资源贫乏忍耐型物种占优势的群落,N素输入增加将会有利于竞争性强的物种,特别是禾草类物种的侵入[44, 51, 110]。
3.1.4 同质性假说
在自然状态下土壤矿质元素的异质化为多样性的维持提供了条件,特定养分元素的大量输入将降低土壤养分元素的异质化程度,随着土壤可利用养分空间同质性的增加,物种多样性将会降低[111-112]。元素输入对物种多样性和群落结构的影响可能取决于系统的初始状态,主要包括元素输入的数量、土壤异质性以及地上植被类型。Theodose和Bowman[51]发现同一气候环境下两个高山苔原群落,多样性对养分元素输入的响应因土壤资源可利用性不同而异:贫瘠系统多样性趋于升高,肥沃系统多样性趋于降低。通常,长期贫瘠或肥沃条件下,由于植被对环境的长期适应,系统对养分添加反应不敏感[71],但是在相对N丰富的地区(如热带亚热带原始森林),大气N沉降所导致的N异质性降低可能使这些系统多样性的维持更加脆弱[7]。不同群落的养分含量和异质性的程度是不同的,因此植物多样性对养分动态变化的响应也有差异。Bartels和Chen[113]对不同林下层植物的响应进行对数线性分析发现:资源量和资源异质性是否成为某一研究中决定植物多样性的因素,取决于研究对象所处的演替阶段,干扰程度以及生物群落的存在现状。资源的数量控制着年轻森林和成熟森林的物种多样性,而资源的异质性则控制着原始森林的多样性[113]。
3.1.5 间接诱导机制
养分元素可利用程度将会调控与植物密切相关的土壤微生物群落,间接影响植物的生长和群落组成。菌根真菌多样性对于维持植物多样性以及生态系统功能具有重要作用。土壤养分可用性的增加使菌根真菌的多样性降低,间接影响植物多样性的维持已经被不少研究证明[37, 114]。养分添加改变了土壤中的真菌群落的组成[115](如丛枝菌根真菌),并使其孢子数显著降低[116]。微生物的新陈代谢活动(如分解、吸收和固氮),甚至是微生物种类数量对养分添加的响应不同,其结果可能会影响到地下生态位,进而对植物多样性产生一系列长期影响。
3.2 植物个体水平上的机理:增加植物对逆境胁迫的敏感性
3.2.1 元素失衡
系统元素限制的平衡被打破也将反映在植物体内元素失衡上。如N素的过量输入可以导致植物体内元素失衡,通常表现为植物根冠比和体内元素比(特别是Ca:Al 和 Mg:N)降低[117],进而导致净光合以及光合N利用率降低,森林生长减缓甚至树木寿命的下降[118]。Roem 和Berendse[102]在荷兰草地的研究发现,植物体内N:P 和N:K的升高对植物多样性具有负效应。目前的研究普遍认为,当某些元素的比值在一定的范围内时(表明几种养分元素共同限制),群落存在最多的植物种类[119]。就单个的物种而言,组织器官中N、P浓度高的植物在添加N或P后反应更为敏感,这些植物N、P的浓度较N:P可以更好地预测其对元素添加后的反应[120]。也有学者认为,养分添加后叶片养分的积累可能表明了营养的储存,也有可能涉及到了植物应对养分可用性的不确定性策略[121],因此仍需要更多植物生理学和计量化学的研究解释。
3.2.2 对环境变化敏感性的改变
元素的过量输入使植物对环境因子(如干旱、霜冻和真菌侵袭)敏感性增强[75, 122]。以N素为例,针叶中N含量在1.3%—1.8%(干重)时,欧洲赤松(PinussilvestrisL.)具有最佳的抗寒性,超过该范围其对于冷害敏感性也逐渐增强[123]。施N可以引起植物Ca、K等养分失衡,通过扰乱体内代谢过程和细胞膜的稳定性降低植物的抗寒能力或者通过扰乱植物的物候特征增加受到冷害的风险。植物体内的N主要以蛋白质和氨基酸的形式存在,外加N可以引起叶片中的蛋白质和游离氨基酸浓度增加,为天敌的寄生和繁殖提供有利的条件[124-125]。Nordin等[122]发现N添加(50 kg N hm-2a-1)使瑞典针叶林林下植物感染致病真菌(Podosphaera和Valdensia)的多度较周围环境提高60倍以上,植物当年茎芽受鳞翅目(Lepidoptera)幼虫损害的程度也提高了2—5倍。P和K添加使叶片P、K浓度提高,增加植食动物的取食率也有类似报道[126]。另一个方面养分元素可用性增加降低植物体内防御物质的数量,从而易于被寄生和取食[127]。Rowe等[103]发现养分添加(如NPK)增加了叶片中的含N量并降低了可溶性多酚的含量,使叶片有利于掠食者的取食。这些响应最终会导致植物群落的营养级结构和竞争格局上发生改变。
总之,多样性的维持是一个长时间多因子相互作用的复杂过程,不同生态系统中植物对养分元素添加响应的机理不尽相同。但无论是在生态系统水平上还是植物个体水平上,各种影响因素同时作用,在环境压力下,植物如果不适应养分元素添加带来的环境变化,将逐渐被淘汰。
4 目前研究存在的问题和展望
4.1 空间尺度受限
研究对象的地域和类型的不均衡。矿质元素输入影响植物多样性的现有理论主要是建立在北方和温带地区的研究基础之上,然而在热带亚热带地区,该方面的研究依然非常缺乏[7, 21, 35]。如在热带森林,虽然面积仅占全球陆地面积的7%,但却集中了50%以上的陆地物种,是陆地生物多样性的分布中心。一旦这些地区遭遇多样性退化,损失将难以估量和弥补,因此具有重要研究意义。
4.2 时间尺度受限
长期定位研究较少。除了在草地生态系统有少数长期(如>10a)的研究外,其他生态系统上的长期研究鲜有报道(表1)。就现存的少数长期研究结果显示,短期数据不能充分地预测长期植物多样性对养分动态变化的响应[60, 128]。因为随着时间的推移物种丧失的风险可能是倍增的而非累加式的。因此,今后研究需要短期的控制实验和长期定位实验相结合,生长周期和群落形成的早期晚期相结合,从而更好地从生态系统各个水平以及自然梯度上认识矿质元素添加对多样性的影响,以阐明多样性响应的机理和植物的生存策略。
4.3 研究方法的局限性
研究手段比较单一分散,难以真实反映元素变化和N沉降的自然状况。目前的研究方法主要有野外控制实验,空间梯度对比以及模型模拟的方法。野外控制实验具有目的性强和条件易操纵的优点,但不同实验的方法和干扰程度不同,限制了不同研究之间的可比性。空间梯度上的监测避免了控制实验的问题,但往往存在本底异质性给研究结果带来的不确定性因素。模型模拟的方法可以很好地预测多样性在未来的变化情况,但是需要建立在更准确的控制实验之上。森林林冠层模拟N沉降等实验方法将应用于野外控制实验,不同方法间的联系和结合运用也是发展趋势。
4.4 各生态因子的耦合研究不足
全球变化背景下,探讨不同因子(如大量元素和微量元素、降水、温度和CO2等)之间的交互作用对生态系统生物多样性的影响趋势,需要考虑研究的空间尺度(如从系统水平到基因水平)和时间尺度,以更充分了解多样性动态变化的机制。目前对这些全球变化热点问题中不同因素交互作用的研究已经受到重视[94],未来还会有更新的突破。
4.5 缺乏有关停止养分元素添加后植物多样性恢复的生态学研究
特别是有关森林生态系统元素添加停止后的监测报道极少。目前,元素添加后的恢复研究仅有草原上的个别报道。我们对养分添加所导致的长期效应认识非常局限,不足以指导恢复工作。元素添加对植物多样性产生的影响是否可以在停止添加后得到恢复、以及恢复过程中的影响因素等,还需要更加系统的研究。此外,生态系统中可能存在元素循环条件改变后的反馈适应过程,系统对于元素添加的适应性有多大,随着时间推移其群落结构调整规律如何有待解决[7, 36]。
[1] Sodhi N S, Koh L P, Brook B W, Ng P K L. Southeast Asian biodiversity: an impending disaster. Trends in Ecology & Evolution, 2004, 19(12): 654- 660.
[2] Tilman D, Lehman C. Human-caused environmental change: impacts on plant diversity and evolution. Proceedings of the National Academy of Sciences of the United States of America, 2001, 98(10): 5433- 5440.
[3] Wilson E O. The Diversity of Life. USA: Harvard University Press, 1992.
[4] Hooper D U, Adair E C, Cardinale B J, Byrnes J E, Hungate B A, Matulich K L, Gonzalez A, Duffy J E, Gamfeldt L, O′Connor M I. A global synthesis reveals biodiversity loss as a major driver of ecosystem change. Nature, 2012, 486(7401): 105- 108.
[5] MacDougall A S, McCann K S, Gellner G, Turkington R. Diversity loss with persistent human disturbance increases vulnerability to ecosystem collapse. Nature, 2013, 494(7435): 86- 89.
[6] Sala O E, Chapin F S III, Armesto J J, Berlow E, Bloomfield J, Dirzo R, Huber-Sanwald E, Huenneke L F, Jackson R B, Kinzig A, Leemans R, Lodge D M, Mooney H A, Oesterheld M, Poff N L, Sykes M T, Walker B H, Walker M, Wall D H. Global biodiversity scenarios for the year 2100. Science, 2000, 287(5459): 1770- 1774.
[7] Bobbink R, Hicks K, Galloway J, Spranger T, Alkemade R, Ashmore M, Bustamante M, Cinderby S, Davidson E, Dentener F, Emmett B, Erisman J W, Fenn M, Gilliam F, Nordin A, Pardo L, De Vries W. Global assessment of nitrogen deposition effects on terrestrial plant diversity: a synthesis. Ecological Applications, 2010, 20(1): 30- 59.
[8] Matson P, Lohse K A, Hall S J. The globalization of nitrogen deposition: consequences for terrestrial ecosystems. Ambio, 2002, 31(2): 113- 119.
[9] Magill A H, Aber J D, Currie W S, Nadelhoffer K J, Martin M E, McDowell W H, Melillo J M, Steudler P. Ecosystem response to 15 years of chronic nitrogen additions at the Harvard Forest LTER, Massachusetts, USA. Forest Ecology and Management, 2004, 196(1): 7- 28.
[10] Phoenix G K, Hicks W K, Cinderby S, Kuylenstierna J C I, Stock W D, Dentener F J, Giller K E, Austin A T, Lefroy R D B, Gimeno B S, Ashmore M R, Ineson P. Atmospheric nitrogen deposition in world biodiversity hotspots: the need for a greater global perspective in assessing N deposition impacts. Global Change Biology, 2006, 12(3): 470- 476.
[11] Marklein A R, Houlton B Z. Nitrogen inputs accelerate phosphorus cycling rates across a wide variety of terrestrial ecosystems. New Phytologist, 2012, 193(3): 696- 704.
[12] Vitousek P M, Porder S, Houlton B Z, Chadwick O A. Terrestrial phosphorus limitation: mechanisms, implications, and nitrogen- phosphorous interactions. Ecological Applications, 2010, 20(1): 5- 15.
[13] Lu X K, Mo J M, Gundersern P, Zhu W X, Zhou G Y, Li D J, Zhang X. Effect of simulated N deposition on soil exchangeable cations in three forest types of subtropical China. Pedosphere, 2009, 19(2): 189- 198.
[14] Lucas R W, Klaminder J, Futter M N, Bishop K H, Egnell G, Laudon H, Högberg P. A meta-analysis of the effects of nitrogen additions on base cations: implications for plants, soils, and streams. Forest Ecology and Management, 2011, 262(2): 95- 104.
[15] Liu X J, Zhang Y, Han W X, Tang A, Shen J L, Cui Z L, Vitousek P, Erisman J W, Goulding K, Christie P, Fangmeier A, Zhang F S. Enhanced nitrogen deposition over China. Nature, 2013, 494(7438): 459- 462.
[16] Cui S, Shi Y, Groffman P M, Schlesinger W H, Zhu Y G. Centennial-scale analysis of the creation and fate of reactive nitrogen in China (1910- 2010). Proceedings of the National Academy of Sciences of the United States of America, 2013, 110(6): 2052- 2057.
[17] 谢迎新, 张淑利, 冯伟, 赵旭, 郭天财. 大气氮素沉降研究进展. 中国生态农业学报, 2010, 18(4): 897- 904.
[18] Lu X K, Gilliam F S, Yu G R, Li L H, Mao Q G, Chen H, Mo J M. Long-term nitrogen addition decreases carbon leaching in a nitrogen-rich forest ecosystem. Biogeosciences, 2013, 10(6): 3931- 3941.
[19] Aber J D, Goodale C L, Ollinger S V, Smith M L, Magill A H, Martin M E, Hallett R A, Stoddard J L. Is nitrogen deposition altering the nitrogen status of northeastern forests? BioScience, 2003, 53(4): 375- 389.
[20] 张燕, 崔学民, 樊明寿. 大气氮沉降及其对草地生物多样性的影响. 草业科学, 2007, 24(7): 12- 17.
[21] Lu X K, Mo J M, Dong S F. Effects of nitrogen deposition on forest biodiversity. Acta Ecologica Sinica, 2008, 28(11): 5532- 5548.
[22] Elser J J, Bracken M E, Cleland E E, Gruner D S, Harpole W S, Hillebrand H, Ngai J T, Seabloom E W, Shurin J B, Smith J E. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecology Letters, 2007, 10(12): 1135- 1142.
[23] Chapin F S III, Vitousek P M, Van Cleve K. The nature of nutrient limitation in plant communities. The Anmerican Naturalist, 1986, 127(1): 48- 58.
[24] Koerselman W, Meuleman A F M. The vegetation N:P ratio: a new tool to detect the nature of nutrient limitation. Journal of Applied Ecology, 1996, 33(6): 1441- 1450.
[26] Phuyal M, Artz R R E, Sheppard L, Leith I D, Johnson D. Long-term nitrogen deposition increases phosphorus limitation of bryophytes in an ombrotrophic bog. Plant Ecology, 2008, 196(1): 111- 121.
[27] Grime J P. Competitive exclusion in herbaceous vegetation. Nature, 1973, 242(5396): 344- 347.
[28] Tilman D, Kilham S S, Kilham P. Phytoplankton community ecology: the role of limiting nutrients. Annual Review of Ecology and Systematics, 1982, 13: 349- 372.
[29] Gotelli N J, Ellison A M. Nitrogen deposition and extinction risk in the northern pitcher plant,Sarraceniapurpurea. Ecology, 2002, 83(10): 2758- 2765.
[30] Suding K N, Collins S L, Gough L, Clark C, Cleland E E, Gross K L, Milchunas D G, Pennings S. Functional- and abundance-based mechanisms explain diversity loss due to N fertilization. Proceedings of the National Academy of Sciences of the United States of America, 2005, 102(12): 4387- 4392.
[31] Van Den Berg L J L, Vergeer P, Rich T C G, Smart S M, Guest D A N, Ashmore M R. Direct and indirect effects of nitrogen deposition on species composition change in calcareous grasslands. Global Change Biology, 2011, 17(5): 1871- 1883.
[32] Petchey O L, Gaston K J. Extinction and the loss of functional diversity. Proceedings of the Royal Society B: Biological Sciences, 2002, 269(1501): 1721- 1727.
[33] Chapin III F S, Zavaleta E S, Eviner V T, Naylor R L, Vitousek P M, Reynolds H L, Hooper D U, Lavorel S, Sala O E, Hobbie S E, Mack M C, Diaz S. Consequences of changing biodiversity. Nature, 2000, 405(6783): 234- 242.
[34] Lu X K, Mo J M, Gilliam F S, Yu G R, Zhang W, Fang Y T, Huang J. Effects of experimental nitrogen additions on plant diversity in tropical forests of contrasting disturbance regimes in southern China. Environmental Pollution, 2011, 159(10): 2228- 2235.
[35] Lu X K, Mo J M, Gilliam F S, Zhou G Y, Fang Y T. Effects of experimental nitrogen additions on plant diversity in an old-growth tropical forest. Global Change Biology, 2010, 16(10): 2688- 2700.
[36] Siddique I, Vieira I C G, Schmidt S, Lamb D, Carvalho C J R, Figueiredo R de O, Blomberg S, Davidson E A. Nitrogen and phosphorus additions negatively affect tree species diversity in tropical forest regrowth trajectories. Ecology, 2010, 91(7): 2121- 2131.
[37] Wu J P, Liu W F, Fan H B, Huang G M, Wan S Z, Yuan Y H, Ji C F. Asynchronous responses of soil microbial community and understory plant community to simulated nitrogen deposition in a subtropical forest. Ecology and Evolution, 2013, 3(11): 3895- 3905.
[38] VanderSchaaf C L, Moore J A, Kingery J L. The effect of multi-nutrient fertilization on understory plant diversity. Northwest Science, 2000, 74(4): 316- 324.
[39] Johansson O, Palmqvist K, Olofsson J. Nitrogen deposition drives lichen community changes through differential species responses. Global Change Biology, 2012, 18(8): 2626- 2635.
[40] Gilliam F S, Hockenberry A W, Adams M B. Effects of atmospheric nitrogen deposition on the herbaceous layer of a central Appalachian hardwood forest. Journal of the Torrey Botanical Society, 2006, 133(2): 240- 254.
[41] Talhelm A F, Burton A J, Pregitzer K S, Campione M A. Chronic nitrogen deposition reduces the abundance of dominant forest understory and groundcover species. Forest Ecology and Management, 2013, 293: 39- 48.
[42] Thomas S C, Halpern C B, Falk D A, Liguori D A, Austin KA . Plant diversity in managed forests: understory responses to thinning and fertilization. Ecological Applications, 1999, 9(3): 864- 879.
[43] Jacobson T K, Bustamante M M D C, Kozovits A R. Diversity of shrub tree layer, leaf litter decomposition and N release in a Brazilian Cerrado under N, P and N plus P additions. Environmental Pollution, 2011, 159(10): 2236- 2242.
[44] Bustamante M M C, de Brito D Q, Kozovits A R, Luedemann G, de Mello T R B, Pinto A de S, Munhoz C B R, Takahashi F S C. Effects of nutrient additions on plant biomass and diversity of the herbaceous-subshrub layer of a Brazilian savanna (Cerrado). Plant Ecology, 2012, 213(5): 795- 808.
[45] Huenneke L F, Hamburg S P, Koide R, Mooney H A, Vitousek P M. Effects of soil resources on plant invasion and community structure in Californian serpentine grassland. Ecology, 1990, 71(2): 478- 491.
[46] Bobbink R. Effects of nutrient enrichment in Dutchchalk grassland. Journal of Applied Ecology, 1991, 28(1): 28- 41.
[47] Jarchow M E, Liebman M. Nitrogen fertilization increases diversity and productivity of prairie communities used for bioenergy. Globle Change Biology: Bioenergy, 2013, 5(3): 281- 289.
[48] Alhamad M N, Alrababah M A, Gharaibeh M A. Impact of burning and fertilization on dry Mediterranean grassland productivity and diversity. Acta Oecologica, 2012, 40: 19- 26.
[49] Wang C T, Long R J, Wang Q L, Liu W, Jing Z C, Zhang L. Fertilization and litter effects on the functional group biomass, species diversity of plants, microbial biomass, and enzyme activity of two alpine meadow communities. Plant and Soil, 2010, 331(1/2): 377- 389.
[50] Bai Y F, Wu J G, Clark C M, Naeem S, Pan Q M, Huang J H, Zhang L X, Han X G. Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: evidence from inner Mongolia Grasslands. Global Change Biology, 2010, 16(1): 358- 372.
[51] Theodose T A, Bowman W D. Nutrient availability, plant abundance, and species diversity in two alpine tundra communities. Ecology, 1997, 78(6): 1861- 1872.
[52] Song L, Bao X M, Liu X J, Zhang F S. Impact of nitrogen addition on plant community in a semi-arid temperate steppe in China. Journal of Arid Land, 2012, 4(1): 3- 10.
[53] Fang Y, Xun F, Bai W M, Zhang W H, Li L H. Long-Term nitrogen addition leads to loss of species richness due to litter accumulation and soil acidification in a temperate steppe. PLoS ONE, 2012, 7(10): e47369.
[54] Oerlemans J, von Boberfeld W O, Wolf D. Impact of long-term nutrient supply on plant species diversity in grassland: an experimental approach on conventionally used pastures. Journal of Applied Botany and Food Quality-Angewandte Botanik, 2007, 81(2): 151- 157.
[55] Wedin D A, Tilman D. Influence of nitrogen loading and species composition on the carbon balance of grasslands. Science, 1996, 274(5293): 1720- 1723.
[56] Clark C M, Tilman D. Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands. Nature, 2008, 451(7179): 712- 715.
[57] Mrkvicka J, Vesela M. The influence of long-term fertilization on species diversity and yield potential of permanent meadow stand. Rostlinna Vyroba, 2002, 48(2): 69- 75.
[58] Smits N A C, Willems J H, Bobbink R. Long-term after-effects of fertilisation on the restoration of calcareous grasslands. Applied Vegetation Science, 2008, 11(2): 279- 286.
[59] Pierik M, van Ruijven J, Bezemer T M, Geerts R H E M, Berendse F. Recovery of plant species richness during long-term fertilization of a species-rich grassland. Ecology, 2011, 92(7): 1393- 1398.
[60] Silvertown J, Poulton P, Johnston E, Edwards G, Heard M, Biss P M. The park grass experiment 1856- 2006: its contribution to ecology. Journal of Ecology, 2006, 94(4): 801- 814.
[61] Xu Z G, Yan B X, He Y, Song C C. Nutrient limitation and wetland botanical diversity in Northeast China: can fertilization influence on species richness? Soil Science, 2007, 172(1): 86- 93.
[62] Bergamini A, Pauli D. Effects of increased nutrient supply on bryophytes in montane calcareous fens. Journal of Bryology, 2001, 23(4): 331- 339.
[63] Pauli D, Peintinger M, Schmid B. Nutrient enrichment in calcareous fens: effects on plant species and community structure. Basic and Applied Ecology, 2002, 3(3): 255- 266.
[64] Kirkham F W, Mountford J O, Wilkins R J. The effects of nitrogen, potassium and phosphorus addition on the vegetation of a Somerset peat moor under cutting management. Journal of Applied Ecology, 1996, 33(5): 1013- 1029.
[65] Goldberg D E, Miller T E. Effects of different resource additions on species diversity in an annual plant community. Ecology, 1990, 71(1): 213- 225.
[66] Rajaniemi T K, Allison V J, Goldberg D E. Root competition can cause a decline in diversity with increased productivity. Journal of Ecology, 2003, 91(3): 407- 416.
[67] Zhao X F, Xu H L, Zhang P, Fu J Y, Tu W X, Zhang Q Q. The effects of nutrient addition on plant species diversity in desert grassland, Xinjiang, northwest China. Quaternary International, 2013, 298: 152- 160.
[68] Brooks M L. Effects of increased soil nitrogen on the dominance of alien annual plants in the Mojave Desert. Journal of Applied Ecology, 2003, 40(2): 344- 353.
[69] Chiarucci A, Maccherini S, Bonini I, De Dominicis V. Effects of nutrient addition on species diversity and ground cover of “serpentine” vegetation. Plant Biosystems, 1998, 132(2): 143- 150.
[70] Roem W J, Klees H, Berendse F. Effects of nutrient addition and acidification on plant species diversity and seed germination in heathland. Journal of Applied Ecology, 2002, 39(6): 937- 948.
[71] Huberty L E, Gross K L, Miller C J. Effects of nitrogen addition on successional dynamics and species diversity in Michigan old-fields. Journal of Ecology, 1998, 86(5): 794- 803.
[72] Stevenson F C, Légère A, Simard R R, Angers D A, Pageau D, Lafond J. Weed species diversity in spring barley varies with crop rotation and tillage, but not with nutrient source. Weed Science, 1997, 45(6): 798- 806.
[73] Yin L C, Cai Z C, Zhong W H. Changes in weed community diversity of maize crops due to long-term fertilization. Crop Protection, 2006, 25(9): 910- 914.
[74] Jonasson S. Plant-responses to fertilization and species removal in tundra related to community structure and clonality. Oikos, 1992, 63(3): 420- 429.
[75] Robinson C H, Wookey P A, Lee J A, Callaghan T V, Press M C. Plant community responses to simulated environmental change at a high arctic polar semi-desert. Ecology, 1998, 79(3): 856- 866.
[76] Nordin A, Strengbom J, Witzell J, Nasholm T, Ericson L. Nitrogen deposition and the biodiversity of boreal forests: implications for the nitrogen critical load. Ambio, 2005, 34(1): 20- 24.
[77] Rainey S M, Nadelhoffer K J, Silver W L, Downs M R. Effects of chronic nitrogen additions on understory species in a red pine plantation. Ecological Applications, 1999, 9(3): 949- 957.
[78] Bauer G A, Bazzaz F A, Minocha R, Long S, Magill A, Aber J, Berntson G M. Effects of chronic N additions on tissue chemistry, photosynthetic capacity, and carbon sequestration potential of a red pine (PinusresinosaAit.) stand in the NE United States. Forest Ecology and Management, 2004, 196(1): 173- 186.
[79] Seftigen K, Moldan F, Linderholm H W. Radial growth of Norway spruce and Scots pine: effects of nitrogen deposition experiments. European Journal of Forest Research, 2013, 132(1): 83- 92.
[80] Isbell F, Tilman D, Polasky S, Binder S, Hawthorne P. Low biodiversity state persists two decades after cessation of nutrient enrichment. Ecology Letters, 2013, 16(4): 454- 460.
[81] 李禄军, 曾德慧, 于占源, 艾桂艳, 杨丹, 毛瑢. 氮素添加对科尔沁沙质草地物种多样性和生产力的影响. 应用生态学报, 2009, 20(8): 1838- 1844.
[82] 张杰绮, 李奇, 任正炜, 杨雪, 王刚. 氮素添加对青藏高原高寒草甸植物群落物种丰富度及其与地上生产力关系的影响. 植物生态学报, 2010, 34(10): 1125- 1131.
[83] 苏洁琼, 李新荣, 回嵘, 赵洋, 刘艳梅. 氮沉降对荒漠化草原草本植物物种多样性和群落组成的影响. 西北植物学报, 2012, 32(4): 795- 801.
[84] Gilliam F S, Yurish B M, Adams M B. Ecosystem nutrient responses to chronic nitrogen inputs at Fernow Experimental Forest, West Virginia. Canadian Journal of Forest Research, 1996, 26(2): 196- 205.
[85] Henkin Z, Sternberg M, Seligman N G, Noy-Meird I. Species richness in relation to phosphorus and competition in a Mediterranean dwarf-shrub community. Agriculture, Ecosystems and Environment, 2006, 113(1/4): 277- 283.
[86] Janssens F, Peeters A, Tallowin J R B, Bakker J P, Bekker R M, Fillat F, Oomes M J M. Relationship between soil chemical factors and grassland diversity. Plant and Soil, 1998, 202(1): 69- 78.
[87] Wassen M J, Venterink H O, Lapshina E D, Tanneberger F. Endangered plants persist under phosphorus limitation. Nature, 2005, 437(7058): 547- 550.
[88] Ceulemans T, Merckx R, Hens M, Honnay O. Plant species loss from European semi-natural grasslands following nutrient enrichment - is it nitrogen or is it phosphorus? Global Ecology and Biogeography, 2013, 22(1): 73- 82.
[89] Crawley M J, Johnston A E, Silvertown J, Dodd M, de Mazancourt C, Heard M S, Henman D F, Edwards G R. Determinants of species richness in the park grass experiment. The American Naturalist, 2005, 165(2): 179- 192.
[90] Ren Z W, Li Q, Chu C J, Zhao L Q, Zhang J Q, Ai D, Yang Y B, Wang G. Effects of resource additions on species richness and ANPP in an alpine meadow community. Journal of Plant Ecology, 2010, 3(1): 25- 31.
[91] McLaughlin S, Wimmer R. Calcium physiology and terrestrial ecosystem processes. New Phytologist, 1999, 142(3): 373- 417.
[92] Cotrufo M F, Ineson P, Scott A. Elevated CO2reduces the nitrogen concentration of plant tissues. Global Change Biology, 1998, 4(1): 43- 54.
[93] Reich P B. Elevated CO2reduces losses of plant diversity caused by nitrogen deposition. Science, 2009, 326(5958): 1399- 1402.
[94] Newman E I. Competition and diversity in herbaceous vegetation. Nature, 1973, 244(5414): 310- 311.
[95] Hautier Y, Niklaus P A, Hector A. Competition for light causes plant biodiversity loss after eutrophication. Science, 2009, 324(5927): 636- 638.
[96] Weiner J, Thomas S C. Size variability and competition in plant monocultures. Oikos, 1986, 47(2): 211- 222.
[97] Oksanen J. Is the humped relationship between species richness and biomass an artifact due to plot size? Journal of Ecology, 1996, 84(2): 293- 295.
[98] Rajaniemi T K. Why does fertilization reduce plant species diversity?Testing three competition-based hypotheses. Journal of Ecology, 2002, 90(2): 316- 324.
[99] Callahan H S, Del Fierro K, Patterson A E, Zafar H. Impacts of elevated nitrogen inputs on oak reproductive and seed ecology. Global Change Biology, 2008, 14(2): 285- 293.
[100] Ochoa-Hueso R, Manrique E. Nitrogen fertilization and water supply affect germination and plant establishment of the soil seed bank present in a semi-arid Mediterranean scrubland. Plant Ecology, 2010, 210(2): 263- 273.
[101] Slik J W F, Raes N, Aiba S I, Brearley F Q, Cannon C H, Meijaard E, Nagamasu H, Nilus R, Paoli G, Poulsen A D, Sheil D, Suzuki E, van Valkenburg J L C H, Webb C O, Wilkie P, Wulffraat S. Environmental correlates for tropical tree diversity and distribution patterns in Borneo. Diversity and Distributions, 2009, 15(3): 523- 532.
[102] Roem W J, Berendse F. Soil acidity and nutrient supply ratio as possible factors determining changes in plant species diversity in grassland and heathland communities. Biological Conservation, 2000, 92(2): 151- 161.
[103] Rowe E C, Healey J R, Edwards-Jones G, Hills J, Howells M, Jones D L. Fertilizer application during primary succession changes the structure of plant and herbivore communities. Biological Conservation, 2006, 131(4): 510- 522.
[104] Azevedo L B, van Zelm R, Hendriks A J, Bobbink R, Huijbregts M A J. Global assessment of the effects of terrestrial acidification on plant species richness. Environmental Pollution, 2013, 174: 10- 15.
[105] Lee J A. The calcicole-calcifuge problem revisited. Advances in Botanical Research, 1998, 29: 1- 30.
[106] Bidwell S, Attiwill P M, Adams M A. Nitrogen availability and weed invasion in a remnant native woodland in urban Melbourne. Austral Ecology, 2006, 31(2): 262- 270.
[107] Bubier J L, Smith R, Juutinen S, Moore T R, Minocha R, Long S, Minocha S. Effects of nutrient addition on leaf chemistry, morphology, and photosynthetic capacity of three bog shrubs. Oecologia, 2011, 167(2): 355- 368.
[108] 王满莲, 冯玉龙. 紫茎泽兰和飞机草的形态、生物量分配和光合特性对氮营养的响应. 植物生态学报, 2005, 29(5): 697- 705.
[109] Baruch Z, Goldstein G. Leaf construction cost, nutrient concentration, and net CO2assimilation of native and invasive species in Hawaii. Oecologia, 1999, 121(2): 183- 192.
[110] van der Welle M E W, Vermeulen P J, Shaver G R, Berendse F. Factors determining plant species richness in Alaskan arctic tundra. Journal of Vegetation Science, 2003, 14(5): 711- 720.
[111] Gilliam F S. Response of the herbaceous layer of forest ecosystems to excess nitrogen deposition. Journal of Ecology, 2006, 94(6): 1176- 1191.
[112] Gundale M J, Metlen K L, Fiedler C E, DeLuca T H. Nitrogen spatial heterogeneity influences diversity following restoration in a Ponderosa Pine Forest, Montana. Ecological Applications, 2006, 16(2): 479- 489.
[113] Bartels S F, Chen H Y H. Is understory plant species diversity driven by resource quantity or resource heterogeneity? Ecology, 2010, 91(7): 1931- 1938.
[114] Johnson N C, Wolf J, Koch G W. Interactions among mycorrhizae, atmospheric CO2and soil N impact plant community composition. Ecology Letters, 2003, 6(6): 532- 540.
[115] Liu L, Gundersen P, Zhang T, Mo J M. Effects of phosphorus addition on soil microbial biomass and community composition in three forest types in tropical China. Soil Biology and Biochemistry, 2012, 44(1): 31- 38.
[116] Bhadalung N N, Suwanarit A, Dell B, Nopamornbodi O, Thamchaipenet A, Rungchuang J. Effects of long-term NP-fertilization on abundance and diversity of arbuscular mycorrhizal fungi under a maize cropping system. Plant and Soil, 2005, 270(1): 371- 382.
[117] Whytemare A B, Edmonds R L, Aber J D, Lajtha K. Influence of excess nitrogen deposition on a white spruce (Piceaglauca) stand in southern Alaska. Biogeochemistry, 1997, 38(2): 173- 187.
[118] Aber J D, Magill A, McNulty S G, Boone R D, Nadelhoffer K J, Downs M, Hallett R. Forest biogeochemistry and primary production altered by nitrogen saturation. Water, Air and Soil Pollution, 1995, 85(3): 1665- 1670.
[119] Gusewell S. Responses of wetland graminoids to the relative supply of nitrogen and phosphorus. Plant Ecology, 2005, 176(1): 35- 55.
[120] Mamolos A P, Vasilikos C V, Veresoglou D S. Vegetation in contrasting soil water sites of upland herbaceous grasslands and N: P ratios as indicators of nutrient limitation. Plant and Soil, 2005, 270(1/2): 355- 369.
[121] Ostertag R. Foliar nitrogen and phosphorus accumulation responses after fertilization: an example from nutrient-limited Hawaiian forests. Plant and Soil, 2010, 334(1/2): 85- 98.
[122] Nordin A, Nasholm T, Ericson L. Effects of simulated N deposition on understory vegetation of a boreal coniferous forest. Functional Ecology, 1998, 12(4): 691- 699.
[123] Aronsson A. Frost hardiness in Scots pine (PinussilvestrisL.) II. Hardiness during winter and spring in young trees of different mineral nutrient status. Studia Forestalia Suecica, 1980, 155: 1- 27.
[124] Fluckiger W, Braun S. Nitrogen and its effect on growth, nutrient status and parasite attacks in beech and Norway spruce. Water, Air and Soil Pollution, 1999, 116(1/2): 99- 110.
[125] Strengbom J, Nordin A, Näsholm T, Ericson L. Parasitic fungus mediates change in nitrogen-exposed boreal forest vegetation. Journal of Ecology, 2002, 90(1): 61- 67.
[126] Santiago L S, Wright S J, Harms K E, Yavitt J B, Korine C, Garcia M N, Turner B L. Tropical tree seedling growth responses to nitrogen, phosphorus and potassium addition. Journal of Ecology, 2012, 100(2): 309- 316.
[127] Throop H L, Lerdau M T. Effects of nitrogen deposition on insect herbivory: implications for community and ecosystem processes. Ecosystems, 2004, 7(2): 109- 133.
[128] Mueller K E, Hobbie S E, Tilman D, Reich P B. Effects of plant diversity, N fertilization, and elevated carbon dioxide on grassland soil N cycling in a long-term experiment. Global Change Biology, 2013, 19(4): 1249- 1261.
Responses of terrestrial plant diversity to elevated mineral element inputs
MAO Qinggong1,2, LU Xiankai1,*, CHEN Hao1,2, MO Jiangming1
1KeyLaboratoryofVegetationRestorationandManagementofDegradedEcosystems,SouthChinaBotanicalGarden,ChineseAcademyofSciences,Guangzhou510650,China2UniversityofChineseAcademyofSciences,Beijing100039,China
Biodiversity is the basis for ecosystems normal functioning and for the existence and development of human beings. During the past decades, however, anthropogenic activities have greatly altered the direction and magnitude of mineral elements (such as nitrogen [N], phosphorus [P], and potassium [K]) cycling in terrestrial ecosystems, causing great impact on the structure and functioning of these systems and further declining biodiversity therein. It is projected that N deposition will become the third largest driver of biodiversity loss by the year 2100, following land-use change and climate change. With the rapid development of industry and intensification of agriculture, the rates of nutrient inputs are expected to continue increasing in the future. The responses of terrestrial plant diversity to elevated mineral element inputs are of increasing global concern, especially with the globalization of atmospheric N deposition. However, information regarding the effect of mineral element input on terrestrial biodiversity is very limited in China. Based on available literature, we summarized the effect of nutrient element inputs on plant diversity in terrestrial ecosystems. The objective is to enhance our understanding about the effects of mineral element inputs on terrestrial biodiversity and to provide scientific base for sustainable ecosystem management. This review includes the following four aspects. (1) The concept of “limiting nutrient” and the coupling relationships between nutrient availability and plant diversity. Increase in productivity after nutrient addition is used as a common mode to determine if an individual or a community is limited by certain element. In general, nutrient enrichment changes the balance of elements in plants and affects their growth rate. The competition mechanism is commonly suggested to be responsible for the changes in biodiversity for the long term. Meanwhile, loss of diversity can affect mineral nutrient cycling by changing both nutrient absorption and release processes. (2) We summarized the results from the experimental studies about the effects of individual nutrient addition (especially for N, P, and K) or the combined nutrient addition on terrestrial plant diversity. Plant diversity generally declines in response to excessive nutrient inputs, depending on the quality and duration of nutrient inputs and on the ecosystem types. Compared to N, P availability may have stronger effect on the plant diversity, at least in some temperate regions. Mineral elements may also interact with some other factors such as CO2and further shape the responses in biodiversity. Lichens and mosses are suggested to be the most sensitive to external nutrient inputs, followed by herbaceous plants, shrub, and trees. (3) We discuss the mechanisms responsible for altering plant diversity induced by nutrient addition, at the ecosystem level and at the individual level. At the ecosystem level, competitive exclusion is widely accepted, because some fast-growing species become dominant in a plant community under nutrient enrichment. By decreasing the heterogeneity of soil nutrients, element inputs reduce the possibility of coexistence of different species. Soil acidification and subsequent aluminum toxicity are also suggested to decline plant diversity. In addition, nutrient enrichment can have indirect effects on local/regional plant diversity through promoting alien species invasion. At the individual level, elevated element inputs may lead to nutrient imbalance and increase sensitivity to biotic and abiotic stresses. The above mechanisms may also work together to cause species loss in the long term. (4) Lastly, we point out the limitations of the current research and present the potential research prospects in view of biodiversity protection and ecosystem management in the future.
mineral elements; nutrient limitation; nitrogen deposition; plant biodiversity; terrestrial ecosystem; global changes
国家973计划项目(2014CB954400); 国家自然科学基金项目 (31370498, 41273143)
2013- 11- 02;
日期:2014- 11- 03
10.5846/stxb201311022648
*通讯作者Corresponding author.E-mail: luxiankai@scbg.ac.cn
毛庆功, 鲁显楷, 陈浩, 莫江明.陆地生态系统植物多样性对矿质元素输入的响应.生态学报,2015,35(17):5884- 5897.
Mao Q G, Lu X K, Chen H, Mo J M.Responses of terrestrial plant diversity to elevated mineral element inputs.Acta Ecologica Sinica,2015,35(17):5884- 5897.
