支链氨基酸(BCAA)及水产动物对其需求量的研究进展
2015-01-18王莉苹韩雨哲孙梦蕾尤宏争
■王莉苹 陈 飞 韩雨哲 孙梦蕾 暴 宁 司 滨 尤宏争
(1.大连海洋大学农业部北方海水增养殖重点实验室,辽宁大连 116023;2.天津市水产研究所,天津 300221)
蛋白质是各种生物体的重要组成物质,可占鱼类干重的65%~75%。水产动物不同于畜禽,其消化道内淀粉酶活性较低,对糖类的利用能力较低,因而蛋白质就成为其主要的功能性物质,是水产动物主要的能量来源,这就决定鱼类对蛋白质的需求高于畜禽。试验表明,鱼类的生长主要体现在鱼体肌肉蛋白质沉积量的增加,鱼类的生长性能和饲料利用率与饲料中的氨基酸水平之间存在密切的关系,适量的氨基酸可节约对蛋白质的利用,促进鱼类的生长,提高饲料利用效率。氨基酸的组成、含量及其有效性都会对饲料中蛋白质的营养价值产生影响。
研究发现,所有鱼类都需要10种必需氨基酸来维持其正常的生长发育,任何一种必需氨基酸的缺乏或不足都会导致鱼类生长下降或致使饲料转化率降低,这表明鱼类必须从食物中获取足够的氨基酸以满足其生长的需要。支链氨基酸(Branch chain amino acid,BCAA)占必需氨基酸的40%,其足够的添加量及合理的比例对动物的正常发育十分重要。本文基于对文献的综述及对以往数据的整理,对支链氨基酸的生理功能、代谢及水产动物对其需要量进行研究分析,以期为今后水产动物氨基酸适宜需求量的研究及饲料中氨基酸平衡的配比等提供参考。
1 支链氨基酸(BCAA)
支链氨基酸(BCAA)包括亮氨酸、异亮氨酸和缬氨酸,占动物和植物蛋白总氨基酸的18%~20%,它们在通过细胞膜时竞争相同的载体。另一方面,它们具有相似的化学结构,如:亮氨酸的化学结构为α-氨基异己酸;异亮氨酸化学结构为α-氨基-β-甲基戊酸;缬氨酸的结构式为α-氨基异戊酸,即α-碳链上都含有分支脂肪烃链结构,把这三种氨基酸统称为分支氨基酸或支链氨基酸。
支链氨基酸(BCAA)是动物维持生长所必需的氨基酸,不能在动物体内合成,是必须从日粮中获得的必需氨基酸,并且在代谢过程中存在较为复杂的拮抗机制。支链氨基酸可促进氮储留及蛋白质合成,在一些生物化学反应及部分动物的生长过程中发挥重要的作用。研究表明,支链氨基酸可延长真鲷鱼卵的孵化时间。金宏等(2001)认为,BCAA能改善运动骨骼肌线粒体的功能,消除运动性疲劳,提高大鼠运动耐力,BCAA在生化代谢过程中的拮抗作用也可反映到其对免疫功能的影响上。氨基酸可显著影响体液的免疫功能,而支链氨基酸在其中的表现较为明显。
2 支链氨基酸(BCAA)的生理功能
2.1 亮氨酸(Leu)
亮氨酸是生酮氨基酸,动物机体自身不能合成,作为功能性氨基酸之一,其在调节内分泌、免疫、营养等方面都发挥重要的生物学功能。早期对氨基酸的研究就发现,亮氨酸在血红蛋白合成和维持血糖水平及激素的增加方面起重要作用,并可影响肌肉应激及能量代谢。作为人体必需的氨基酸之一,亮氨酸及其代谢产物可在肌肉蛋白质的合成、骨骼肌微细损伤的修复、糖异生及骨骼肌的葡萄糖摄取方面发挥重要作用。亮氨酸能刺激肌肉蛋白质的合成并可抑制其分解,酮异己酸(KIC)虽具有同样的抑制分解之效,但无改变其合成的能力。经研究,亮氨酸可显著促进κ-酪蛋白基因的表达及蛋白质合成。对哺乳动物的研究发现,亮氨酸可在细胞蛋白质合成和分解的mTOR信号通路中起调节作用,进而对蛋白质的代谢过程产生影响。也有报道称,亮氨酸是通过增强mRNA的翻译速度来促进蛋白质合成的。
对小鼠的研究发现,高浓度的亮氨酸可促进骨骼肌中蛋白质的合成,但也可抑制蛋白质的降解。对水产动物而言,L-亮氨酸对鱼类(鲤、虹鳟等)有明显的诱食效果。日粮亮氨酸水平对吉富罗非鱼的生长性能、体营养成分的组成、生长指标(增重率、特定生长率、饲料系数、蛋白质效率等)及酶的活性、非特异性免疫均可产生显著影响。王用黎对凡纳滨对虾的研究显示,亮氨酸可显著影响凡纳滨对虾的生长性能及对饲料的利用率;还可对凡纳滨对虾血淋巴中的天冬氨酸转氨酶、丙氨酸转氨酶、超氧化物歧化酶和酚氧化酶活性产生显著影响,并可明显改变其肌肉中丙氨酸转氨酶的活性。也有学者认为,随着饲料中亮氨酸水平的升高,虹鳟的脂肪及体蛋白含量也相应升高。
2.2 异亮氨酸(Ile)
异亮氨酸的化学组成与亮氨酸相同,具有四种光学异构体,而自然界中仅存在L-异亮氨酸;L-异亮氨酸是合成激素、酶类的原料,具有促进蛋白质合成和抑制其分解的效果,在生命活动中起着重要作用。
作为水产动物必需氨基酸之一的生糖兼生酮氨基酸,异亮氨酸在日粮中的水平可显著影响吉富罗非鱼的生长性能、饲料利用率、体营养组成、消化吸收能力及非特异性免疫能力。尚晓迪在对草鱼幼鱼异亮氨酸需求量的研究中发现,日粮中适宜异亮氨酸水平能明显提高草鱼幼鱼的增重率、特定生长率和蛋白质效率,降低饲料系数;而投喂异亮氨酸水平达1.67%日粮的饲养组,草鱼的生长性能最佳;此外,异亮氨酸还可显著提高草鱼幼鱼全鱼和肌肉的蛋白含量及肌肉氨基酸总量,同时可降低全鱼和肌肉的水分,降低全鱼的脂肪含量,改善草鱼品质。还有研究显示,在血粉饲料中添加游离异亮氨酸对鲫鱼有明显的促生长作用,饲料效率也可得到提高。此外,异亮氨酸还可影响鲑科、鲤等鱼类的存活率。
异亮氨酸在动物的生长过程中起到重要的作用,异亮氨酸的不足可对机体产生很大的影响。当异亮氨酸缺乏时,支链氨基酸间的平衡被打破,机体产热增加,并可激活脂肪内激素敏感性脂肪酶(HSL),使脂肪分解加快,进而减少脂肪的沉积。而投喂完全不含异亮氨酸的日粮会使动物体重显著下降,同时也会导致血浆和尿液的氨基酸氮降低。
2.3 缬氨酸(Val)
缬氨酸(Val)属支链氨基酸,可参与蛋白质和胺神经递质血清素的合成,最早是从动物的胰脏浸提液中分离而来。缬氨酸是生糖氨基酸,其在免疫球蛋白中所占的比例高于其他氨基酸,当缬氨酸缺乏时会显著影响胸腺及淋巴组织的生长,并可抑制白细胞的增生。研究表明,缬氨酸可刺激前T淋巴细胞和骨骼T淋巴细胞前体分化为成熟的T淋巴细胞。
缬氨酸在水产动物也有一定的研究,在饲料中添加适量的L-缬氨酸,对大西洋鲑、虹鳟、鲤等有明显的诱食效果。王用黎认为,饲料中的缬氨酸可显著影响凡纳滨对虾的生长性能和饲料利用率,同时可提高肌蛋白的沉积;饲料中缬氨酸水平可对凡纳滨对虾血淋巴和肌肉中的丙氨酸转氨酶和天冬氨酸转氨酶活性产生显著影响,还可显著影响血淋巴中超氧化物歧化酶和肝胰腺中碱性磷酸酶的活性;此外,饲料中缬氨酸水平在一定程度上可提高机体的免疫能力。有研究显示,在饲料中添加缬氨酸在一定程度上可提高草鱼、异育银鲫等动物的生长性能。
缬氨酸除可在正常生命活动中发挥重要作用及对部分水产动物产生影响外,还有研究发现,随着饲料中缬氨酸含量的增加,感染了新城疫病毒的初生雏鸡,其抗体滴度及血凝素滴度也逐渐升高;此外,雏鸡的发育和饲料转化率也可得到改善。也有研究认为,给小鼠腹腔注射缬氨酸可增加其绵羊红细胞及免疫后的脾IgM分泌细胞数。综合上述支链氨基酸的生理功能,其对动物的影响见表1。
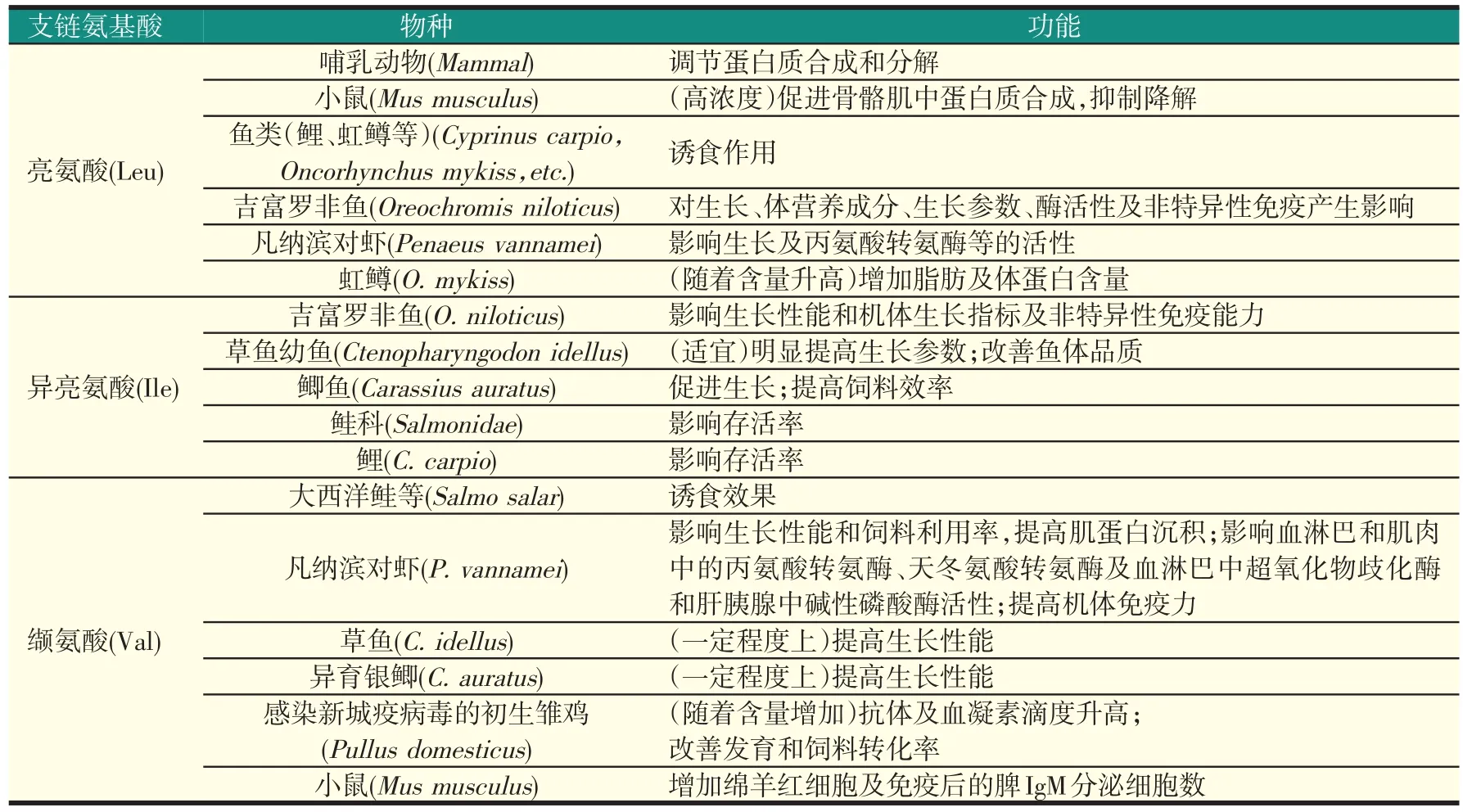
表1 BCAA的生理功能
2.4 支链氨基酸的交互作用
对支链氨基酸之间相互关系的研究,Choo等(1991)发现日粮中过量的亮氨酸可抑制动物的生长,并影响缬氨酸及异亮氨酸的代谢情况。在常规配制饲料中,亮氨酸的过量通常会引起异亮氨酸和缬氨酸的缺乏。据Wilson等(1980)报道,饲料中亮氨酸含量的变化可显著影响血清中异亮氨酸和缬氨酸的浓度,并认为亮氨酸可促进叉尾鮰组织对支链氨基酸的吸收与代谢。Colvin等(1977)研究认为,在对虾饲料中添加亮氨酸或异亮氨酸可促进其生长。对大鳞大麻哈幼鱼的研究发现,日粮中亮氨酸水平影响其对异亮氨酸的需求量,此外,投喂含过量异亮氨酸及次于最适水平亮氨酸的日粮可降低大鳞大麻哈幼鱼的生长率。亮氨酸的拮抗似乎可引起湖鳟对缬氨酸需求量增加,但对异亮氨酸的需求量没有影响;当日粮中异亮氨酸过量时并不能引起湖鳟对日粮中亮氨酸和缬氨酸的需求量产生影响,而在日粮中同时添加亮氨酸可使湖鳟对缬氨酸的需求量增加。
以上研究都认为,日粮中的亮氨酸水平可对异亮氨酸和缬氨酸产生影响,而这三种氨基酸之间的相互关系远不止这些。Aschkenasy认为,日粮中高亮氨酸水平可引起大鼠的免疫抑制,而添加异亮氨酸和缬氨酸可缓解这一现象。其它研究也发现,在饲料中添加适量的异亮氨酸或缬氨酸可以大大减弱过量亮氨酸所造成的免疫抑制作用。异亮氨酸和缬氨酸一起还可促进垂体切除大鼠的膈肌蛋白质合成,但对正常禁食大鼠的肠肌蛋白质合成无促进作用。此外,适宜的异亮氨酸水平可促进支链氨基酸间的平衡,支链氨基酸作为氮和碳骨架的载体,可为谷氨酸及谷氨酰胺的合成提供氮源和碳源。
已证实亮氨酸和缬氨酸可以对牙鲆的生长参数(终体重、体增重和特定生长率)产生交互影响;不仅可在高日粮亮氨酸水平发生拮抗作用,在低亮氨酸水平的条件下增加日粮中缬氨酸的含量也能产生促进作用;这两种氨基酸在饲料中的水平可对日本牙鲆的饲料转化率产生明显的交互作用,也可影响实验鱼血浆中乳酸脱氢酶(LDH)和谷丙转氨酶(GPT)的活性;用低亮氨酸(1.60%)高缬氨酸含量的日粮饲养的鱼可以保护细胞免受氧化影响。
此外,亮氨酸可以与缬氨酸和异亮氨酸一起保护肌肉组织并能为其提供能量,它是一种重要的产血红蛋白的氨基酸,包括血糖水平及对压强耐受力。也有学者认为,缬氨酸与赖氨酸、蛋氨酸、色氨酸和丝氨酸的协同使用可节约罗非鱼对蛋白质的使用量。支链氨基酸的相互关系见表2。从表2可以看出,支链氨基酸之间有的存在拮抗作用,有的则表现为节约作用,对支链氨基酸之间的相互关系有必要做进一步的研究。

表2 BCAA的相互关系
3 各支链氨基酸(BCAA)的代谢
支链氨基酸代谢的重要特征在于转氨酶及脱氢酶活力在各组织中的分布状况。一般来说,谷氨酸脱氢酶及转氨酶联合脱氨基作用,是肝脏内氨基酸分解代谢过程的途径。而研究却发现,在构成机体蛋白质的20种氨基酸中,支链氨基酸是唯一不局限于肝脏而可在全身组织特别是肌肉组织进行分解代谢的必需氨基酸。
支链氨基酸间的代谢。各支链氨基酸结构相似,它们在代谢途径中的转氨基和脱氢作用可共用支链氨基酸转氨酶和脱氢酶,因此它们在代谢过程中会出现拮抗作用,且拮抗程度在各动物间存在差异,最终影响动物的代谢及生长。研究认为,在日粮中添加异亮氨酸和缬氨酸可以防止由过量亮氨酸造成的免疫抑制作用的影响。也有学者发现,各支链氨基酸混合物可抑制蛋白质降解,但异亮氨酸和缬氨酸仅仅抑制垂体切除大鼠膈肌蛋白质的降解。亮氨酸和异亮氨酸之间的相互作用表现为:在实验日粮中加入过量的亮氨酸可抑制家禽对蛋氨酸的生长利用,当补充异亮氨酸后这种抑制现象可消除,表现为对蛋氨酸的生长反应。据报道,当日粮中的异亮氨酸为限制性氨基酸时,高含量亮氨酸可引起肉用仔鸡生长迟缓。此外,Taylor等发现各支链氨基酸在猪中也存在拮抗作用。
支链氨基酸参与蛋白质代谢。研究表明,支链氨基酸在动物蛋白质代谢过程中主要参与蛋白质的合成和降解两个方面。调节蛋白质合成的支链氨基酸主要是亮氨酸,其通过促进肌蛋白多肽链合成的起始促进蛋白质的合成。同时,亮氨酸的代谢产物酮异己酸可促进胰岛素的分泌,并可为谷氨酰胺的合成提供底物,进而抑制糖原异生,减缓肌蛋白的分解。在蛋白质分解过程中支链氨基酸起抗分解作用。
支链氨基酸可作为能源物质。除参与机体蛋白质的合成外,支链氨基酸在体内可作为重要的能源物质,其中每天摄入支链氨基酸的20%被氧化分解,支链氨基酸被氧化生成ATP的效率高于其他氨基酸。当异亮氨酸和缬氨酸缺乏时会引起脂肪动员和棕色脂肪组织(BAT)产热效能增加,也会抑制mTOR、激活AMPK信号通路进而增强胰岛素敏感性。在某些特殊生理时期(运动、饥饿、泌乳等),支链氨基酸可作为体内重要的能量来源,用于分解供能。支链氨基酸在泌乳母畜乳腺中有较高的氧化率,同时可改善母畜的泌乳性能,提高产奶量和乳中干物质乳蛋白和乳脂含量。因此,除氧化分解供能外,支链氨基酸还可影响母畜的泌乳与繁殖性能及动物的免疫与健康状况。如缬氨酸可在乳腺生长发育方面发挥重要作用,缬氨酸与赖氨酸的协同作用可影响母猪的泌乳过程。
支链氨基酸与芳香族氨基酸。支链氨基酸与芳香族氨基酸(苯丙氨酸、酪氨酸和色氨酸)在通过血脑屏障时需要同一个转运载体。Jose等(1979)的研究结果与上述一致,当异亮氨酸与芳香族氨基酸通过血脑屏障时可发生竞争作用,参与神经递质(5-羟色胺、多巴胺及酪胺)生成的调节;高浓度的支链氨基酸能抑制芳香族氨基酸进入脑中,反之,当支链氨基酸浓度降低时,芳香族氨基酸的竞争力增强。
4 水产动物对各支链氨基酸的需求量
氨基酸是构成生物体蛋白质的基本单位,对机体的营养、免疫、生存等有极其重要的作用,是生物有机体的重要组成部分,在生命现象中起着至关重要的作用,也在生命体内物质的代谢调控、信息传递等方面扮演重要角色。
研究认为,各氨基酸的平衡可以提高氨基酸的保持率、氮利用率及促生长作用。试验表明,在饲料中添加适宜含量的支链氨基酸,可显著影响大黄鱼和鲈鱼的生长及摄食。然而,任何动物对氨基酸的需要量都有一个限度,支链氨基酸作为一类氨基酸,其过低或过高的含量都会对机体产生不同程度的影响:对成人的研究显示,缺乏异亮氨酸会降低机体蛋白质的合成,但肝脏蛋白质的合成受此影响并不明显。而缬氨酸的缺乏可使血凝素、补体C3和运铁蛋白水平显著下降。研究发现,当饲料中亮氨酸水平高于生物的最适需要量时,可使体蛋白的沉积率降低。也有报道显示,当饲料亮氨酸水平过量时,会引起大黄鱼和凡纳滨对虾的摄食量下降、生长受阻。
破坏氨基酸平衡会降低动物机体的蛋白质合成,影响动物的采食量及营养物质的利用等,进而影响动物的生长和饲料的转化,可见适宜含量的氨基酸对动物的生长发育至关重要。研究发现,饲料中适宜水平的亮氨酸可提高机体的抵抗力并促进凡纳滨对虾的生长。日粮中适宜的异亮氨酸水平能促进草鱼幼鱼蛋白质的合成,降低脂肪合成及沉积,促进其生长。适宜水平的缬氨酸可促进幼建鲤肠道和肝胰脏的生长发育、提高肝胰脏和肠道消化酶及肠道吸收酶活力、影响幼建鲤肠道微生物菌群的平衡,进而提高机体的消化吸收能力。然而,因各水产动物的机体需要量及其所处外界环境等的不同造成水产动物对各支链氨基酸的需求量有很大差异(见表3)。
由表3可以看出,在淡水鱼类对亮氨酸的需求量中,团头鲂的需要量最低,为2.02%~2.17%;鳗鲡的需求量相对高于大多鱼类(吉富罗非鱼、大黄鱼除外)对亮氨酸的需求,占饲料蛋白的5.30%;而上述水产动物(包括淡水和海水动物)对亮氨酸需求量的研究中,吉富罗非鱼的需求量明显高于其它物种,占饲料蛋白的7.08%~7.27%。淡水鱼类对异亮氨酸需求量的研究中,草鱼幼鱼和吉富罗非鱼的需求量明显高于其他动物;团头鲂对异亮氨酸的需求量显著低于其他鱼类。而在对缬氨酸的研究中发现,虹鳟的需要量明显较低,草鱼幼鱼的需求量显著高于其他淡水鱼种。
对海水鱼的研究发现,牙鲆和大菱鲆对亮氨酸的需求量相对较低,其他几种鱼对亮氨酸的需求量没有明显差别;从表3还可看出,在对异亮氨酸和缬氨酸的研究中,大菱鲆对这两种支链氨基酸的需要量相对都较低,而鲈鱼对这两种氨基酸的需要量则明显高于其他动物。在对三种虾(斑节对虾、日本对虾、凡纳滨对虾)支链氨基酸需求量的研究中发现,日本对虾对亮氨酸和缬氨酸的需要量明显低于斑节对虾和凡纳滨对虾,凡纳滨对虾对此两种氨基酸的需求量明显高于其他两种对虾;而水产虾类对异亮氨酸需求量研究的数据不足,不能就此对其差异做出准确判断。
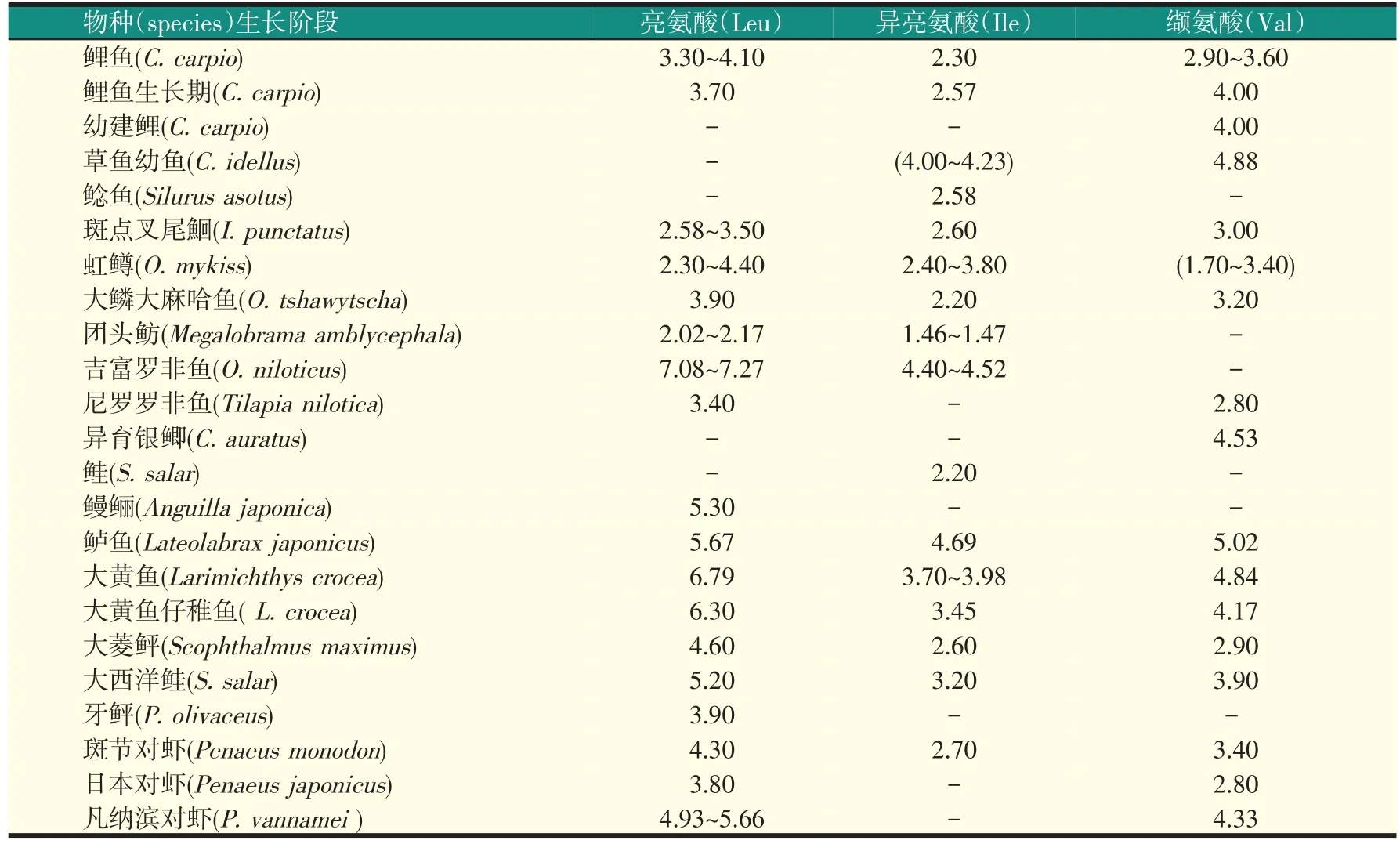
表3 不同水生生物对各支链氨基酸的需求量(%饲料蛋白)
总之,同一水生动物对不同支链氨基酸的需要量不同;同一动物的不同生长阶段对同一支链氨基酸的需要量也不尽相同,这可能是因为不同生长阶段的动物对蛋白的需要不同使得氨基酸需要量产生很大的差异。据Portz等研究发现,不同时期大口黑鲈的机体氨基酸含量不同,且随着生长的进行其氨基酸需要量也会发生变化;不同水产动物对同一支链氨基酸的需求量也各不相同,这可能是因为不同动物对氨基酸的消化吸收率不同所致。整体来看,海水鱼类对各支链氨基酸的需求量相对高于淡水鱼类;而虾对各支链氨基酸的需求量与淡水鱼相比差别不明显,与海水鱼相比其需要量稍低。
以上对水产动物支链氨基酸需求量的分析结果之所以表现出很大的差异,是因为水产动物对氨基酸的需要量受多方面因素的限制,动物的种类和生理状况、年龄和生长发育阶段、食性、水温、水质、饲养密度、饲料蛋白的种类及投喂方法等的不同都会对这种结果产生影响。除此之外,氨基酸的种类和氨基酸之间的拮抗作用及评价指标等也会导致水产动物对各支链氨基酸需求量的差异。
5 研究方向
支链氨基酸在机体生命活动中发挥至关重要的作用,虽然早在很多年前就已有对支链氨基酸的研究,但对支链氨基酸在水产动物方面的研究依旧很有限,特别是对支链氨基酸交互作用的研究。国内在猪、绵羊、大鼠等畜禽方面对支链氨基酸的研究展开的比较多,主要集中在其对动物机体的生产性能、蛋白质合成、免疫、母猪泌乳、代谢、供能等方面;而对水产动物支链氨基酸需要量的研究与国外相比较少。
支链氨基酸在调节动物各种生理机制中发挥重要作用。对支链氨基酸需要量的研究可调整支链氨基酸间及支链氨基酸与其它营养物质的比例,从而不断提高动物的生产性能,以发挥其最佳生产潜力。研究水产动物对支链氨基酸的需求量可以为研究氨基酸代谢提供支撑,为节约蛋白资源、降低饲料成本、科学使用蛋白质日粮提供试验性参考依据;还可进一步完善对氨基酸的需求,为全价配合饲料的研发提供理论基础。
(参考文献85篇,刊略,需者可函索)
