牛磺酸的生理功能及其在鱼类配合饲料中的应用
2015-01-18解文丽关燕云艾春香
■解文丽 关燕云 艾春香
(厦门大学海洋与地球学院,福建厦门 361102)
牛磺酸又称2-氨基乙磺酸(2-Aminoethanesulfon⁃ic acid)、牛胆酸、牛胆素,1827年首先从牛胆汁中分离,纯品为无色或白色晶体,无臭。牛磺酸化学性质稳定,是一种含硫游离氨基酸,不参与体内蛋白的生物合成。牛磺酸虽然不参与蛋白质合成,但它却与胱氨酸、半胱氨酸和蛋氨酸等含硫氨基酸的代谢密切相关。有关牛磺酸的研究以前主要集中在哺乳动物,研究表明,牛磺酸在动物体内发挥重要的生理功能,如调节心血管系统、参与糖类、脂类、蛋白质代谢,促进神经系统发育,提高动物繁殖性能等。近年来,牛磺酸在水产养殖中的应用研究受到人们广泛关注。研究主要集中在两个方面:一是牛磺酸对鱼类的生长性能的影响,如在以植物源蛋白为主的饲料中添加牛磺酸能显著提高肉食性鱼类的生长性能(Gaylord等,2007);二是牛磺酸对养殖鱼类生存能力的影响,饲料中适量添加牛磺酸通过提高红细胞的渗透脆性进而改善鱼类耐缺氧能力以及缓解鱼类绿肝综合征等(Yang等,2013)。鱼类对于牛磺酸的需要量与其种类、大小、食性等密切相关,但过量的牛磺酸也会引起水产养殖动物出现中毒反应(El-Sayed,2014),各种养殖鱼类的牛磺酸适宜需求量需要进一步开展深入系统的研究。本文综述了牛磺酸的结构特点和自然分布、合成与代谢以及其在鱼类中的生理功能及鱼类配合饲料中的应用。
1 牛磺酸的结构特点与自然分布
牛磺酸,是一种β型含硫氨基酸,分子式:HO3SCH2-CH2-NH2,相对分子量为125.1,常温常压下为无色四周针状结晶,无臭,味微酸,熔点310℃,不与蛋白质结合而以游离态存在于生物体内。
牛磺酸主要分布于动物的脑、卵巢、子宫、肾、骨骼、肌肉、血液等组织器官,在植物体内含量较少。牛磺酸含量除在中枢神经系统中仅次于谷氨酸外,在其它组织器官中都远高于其它氨基酸。在神经、肌肉、腺体等可兴奋组织内牛磺酸含量较高,大脑、骨骼肌、肝脏和肾脏中的牛磺酸含量占自由氨基酸总量分别约为19%、50%、25%和50%(Salze等,2015),在心血管组织、胆汁中含量也很高。牛磺酸以鲜蛤中含量最高,其次是孵化19 d左右的鸡胚中含量较多。含牛磺酸的植物种类很少,其中含量最多的是藻类,其次是坚果和豆科植物籽实(黑豆、蚕豆、嫩豌豆、扁豆及南瓜籽等),分别为15~46 nmol/g和9.2~18.7 nmol/g(任和等,2006)。值得注意的是某些动物包括含有丰富牛磺酸的动物,不能够通过自身代谢合成牛磺酸而必须通过饲料获得(Salze等,2015)。
2 牛磺酸在鱼类体内的合成与代谢
2.1 牛磺酸的合成
研究表明,牛磺酸可由半胱氨酸、胱氨酸和蛋氨酸等含硫氨基酸在动物机体内经过一系列酶促反应合成,已知牛磺酸合成的5条途径见图1。
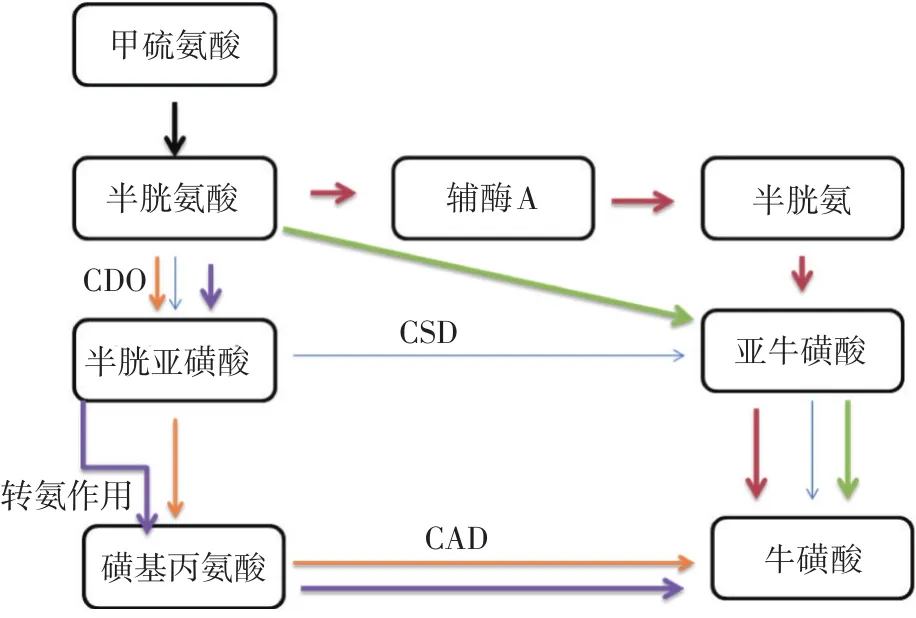
图1 牛磺酸合成的5条途径
哺乳类合成牛磺酸的主要途径是:由半胱氨酸经过半胱亚磺酸、亚牛磺酸最终生成牛磺酸。半胱氨酸经过半胱氨酸双加氧酶(cysteine dioxygenase,CDO,EC 1.13.11.20)氧化生成半胱亚磺酸,CDO是调节半胱氨酸浓度的关键酶。半胱氨酸过多会导致生物体产生中毒现象(Stipanuk等,2006),半胱亚磺酸经过半胱亚磺酸脱羧酶(cysteine sulfinate decarboxylase,CSD,EC 4.1.1.29)脱羧生成亚牛磺酸,CSD是牛磺酸合成的限速酶。CSD活性具有组织特异性,与肌肉、肾脏和肠上皮细胞相比,哺乳动物肝脏中的CSD酶活性更高,说明肝脏是哺乳动物合成牛磺酸的主要场所(Salze等,2015)。
鱼类合成牛磺酸所需要的酶活性普遍低于哺乳类。牛磺酸合成在不同鱼类体内表现出极大的差异,这种差异取决于鱼的种类、大小、摄食习惯、CSD活性以及蛋氨酸含量。牛磺酸的合成能力与鱼类的食性有关:常见的杂食性鱼类,如鲤鱼(Cyprinus carpio)体内牛磺酸含量极低(Salze等,2015),但鲤鱼却具有很强的牛磺酸合成能力,即使饲料中不添加牛磺酸,鲤鱼体内的牛磺酸保有率仍高达280%(Kim等,2008),推断鲤鱼合成牛磺酸的途径不是第一条,很可能是第二条途径。同样属于杂食性鱼类的罗非鱼(Oreochro⁃mis spp),它的CSD活性是0.558 nmol/(min·mg prot.),仍然需要饲料中添加牛磺酸,表明CSD的活性不足(Salze等,2015)。饲料中添加牛磺酸对杂食性的斑点叉尾鮰(Ietalurus Punetaus)稚鱼的生长性能影响不显著(Robinson等,1978),这表明斑点叉尾鮰稚鱼自身能够合成足够的牛磺酸。由此推测,杂食性鱼类具有一定的牛磺酸合成能力。牛磺酸合成在草食性鱼类中研究的较少,草鱼(Ctenopharyngodon idellus)虽然合成牛磺酸的能力很弱,但能满足自身生长发育所需(Yang等,2013)。团头鲂(Megalobrama amblycephala)体内检测到有较高浓度的牛磺酸,占自由氨基酸含量的32.04%(何琳等,2014)。基于上述研究,可以初步推测草食性鱼类也具有一定的牛磺酸合成能力。常见的肉食性鱼类虹鳟(Oncorhynchus mykiss)CSD活性为0.67 nmol/(min·mg prot.),通常情况下饲料中添加牛磺酸能够促进其生长,表明CSD活性不足(Salze等,2015)。当摄食完全植物源蛋白质的配合饲料时,添加牛磺酸促进虹鳟的生长(Gaylord等,2006);当摄食含有鱼粉的配合饲料时,添加牛磺酸不能提高虹鳟的生长状况(Suttisak Boonyoung等,2013)。牙鲆(Parali⁃chthys olivaceus)、真鲷(Pagrosomus major)、五条鰤(Se⁃riola quinqueradiata)体内由于CSD活性较低或者缺乏,导致很少或者无法合成牛磺酸。日本比目鱼的饲料中分别添加0%、0.5%和1.5%的牛磺酸,当添加牛磺酸量为1.5%时显著提高了日本比目鱼的生长性能(Kim等,2007)。金枪鱼(Thunnus thynnus)是典型的肉食性鱼类,其体内只存在CSD,同时CSD活性只有0.001 nmol/(min·mg prot.)(王和伟等,2013),因此金枪鱼饲料中需要添加牛磺酸。鲑鱼(Salmo salar)属于淡水肉食性鱼类,本身可以通过转硫作用合成牛磺酸(El-Sayed,2014),但饲料中添加牛磺酸仍能有效地促进鲑鱼的生长,表明鲑鱼自身合成牛磺酸量不足。由此可以得出,肉食性鱼类合成牛磺酸的能力普遍较低,有的鱼类甚至无法合成牛磺酸,必须通过饲料获取。此外,鱼类的天然摄食习惯以及摄食史可能通过影响其机体内CSD活性进而影响牛磺酸的生物合成(Gaylord等,2006)。蛋氨酸也是鱼类牛磺酸合成的限制性因素之一(Davis等,2014)。目前关于鱼类牛磺酸合成的研究主要集中在几种常见的鱼类,同种鱼类不同阶段的合成能力各异。虹鳟逐渐长大,其肝脏中的CSD浓度显著增加,活性略有提高,而CDO浓度和活性逐渐下降(Wang等,2015)。但至今有关鱼类牛磺酸合成能力的研究有限,需要进一步加大研究力度。
2.2 牛磺酸的代谢
动物体内牛磺酸的合成与代谢处于动态平衡状态。当体内牛磺酸净含量超过一定水平时,牛磺酸会随尿液排出体外;当牛磺酸含量不足时,肾脏通过重吸收来降低牛磺酸的排泄量,以此维持生物体内牛磺酸的动态平衡。
牛磺酸在动物体内的代谢可以分为以下5个途径:①生成结合胆汁酸,胆汁酸盐不仅能够促进脂肪和脂溶性维生素的消化吸收,而且能够增加胆固醇的溶解与排出。②生成牛磺氯胺,牛黄氯胺的产生使细胞免遭氧化损伤。另外,牛黄氯胺通过减少一氧化氮和肿瘤坏死因子的产生来抑制炎症反应的发生,这已经成为牛磺酸生物学功能研究的热点。③生成脒基牛磺酸,在无脊椎动物中,脒基牛磺酸可以作为神经递质的调节剂,其中在很多海洋无脊椎动物中脒基牛磺酸类似于哺乳动物的肌酸。④生成牛磺脲酸,有关牛磺脲酸的研究较少,其生物功能还不清楚。⑤生成羟乙基牛磺酸。研究表明,哺乳动物不能合成羟乙基牛磺酸,其体内的羟乙基牛磺酸可能是由其肠道内细菌合成(王和伟等,2013)。
3 牛磺酸在鱼类中的生理功能
3.1 参与蛋白质、糖类、脂类的代谢调节
牛磺酸可以促进胃液的分泌,提高蛋白酶活力,从而促进蛋白质的分解。牛磺酸能够引起鼠胃酸的分泌(Huang等,2011)。饲料中添加牛磺酸能够显著提高鲤鱼血清甲状腺激素含量,血清甲状腺激素控制着鱼类生长激素的表达和合成,生长激素可以促进鱼类蛋白质的合成。牛磺酸能提高军曹鱼早期幼鱼淀粉酶和胰蛋白酶的活性,从而促进军曹鱼幼鱼对淀粉和蛋白质的利用(Salze等,2012)。当灌喂草鱼的牛磺酸浓度为0.8 mg/ml时,肠道蛋白酶在2 h后达到最大,当牛磺酸浓度过量时,消化酶的活性受到抑制(龙勇等,2004)。
牛磺酸能够促进机体对葡萄糖、淀粉等糖类的利用。胰腺由内外分泌腺两部分组成,其中内分泌腺由大小不同的细胞团——胰岛所组成,胰岛主要由4类细胞组成:A细胞、B细胞、D细胞、PP细胞。A细胞分泌胰高血糖素,升高血糖,B细胞分泌胰岛素,降低血糖。进入胰腺B细胞的牛磺酸能够引起胰岛素的释放,从而使血糖含量降低。当灌喂草鱼的牛磺酸浓度为1 mg/ml时,肠道淀粉酶活性在2 h后达到最高,当牛磺酸过量时淀粉酶活性受到抑制,适量添加牛磺酸能够提高草鱼淀粉酶的活性(龙勇等,2004)。牛磺酸(600~800 mg/kg)显著提高草鱼肝胰脏和肠道脂肪酶和淀粉酶活性(P<0.05)(罗莉等,2006);牛磺酸(800~1 200 mg/kg)显著提高黄河鲤鱼肝胰脏和肠道蛋白酶、脂肪酶和淀粉酶活性(高春生等,2007)。
胆汁酸以胆汁酸盐的形式参与脂肪的消化吸收,而牛磺酸和甘氨酸能够与胆酸和鹅脱氧胆酸等游离胆酸结合形成牛磺胆酸、牛磺鹅脱氧胆酸等结合胆汁酸,进而以胆汁酸盐的形式进入消化道中,促进脂肪分解消化。日本比目鱼体内的牛磺酸是唯一能与胆酸结合的氨基酸,并且在仔鱼和稚鱼期,饲料中牛磺酸的摄食量影响结合胆酸的组成(Kim等,2008)。饲料中添加牛磺酸能够增加草鱼稚鱼的肠系膜脂肪系数,说明牛磺酸与脂肪代谢密切相关(Yang等,2013)。牛磺酸可以降低脂肪在鱼体内的积累,Marit Espe等通过对大西洋鲑幼鱼研究证明,高植物蛋白的饲料中低浓度的牛磺酸能够作用于脂类的代谢和储存(Marit等,2012)。
3.2 抗氧化
牛磺酸及其衍生物能够清除机体活性氧及其他氧化性有害物或增强机体抗氧化系统的抗氧化能力,从而保护动物免遭氧化伤害。饲料中添加1%的牛磺酸能够减少石首鱼肝脏氧化的伤害(Isaura Bañuelos-Vargas等,2014)。当饲料中牛磺酸浓度为1%时,欧洲鲈鱼(Dicentrarchus labrax)前肠中的过氧化氢酶活性与对照组相比增加(Feidantsis等,2014)。
3.3 免疫调节
牛磺酸能够在一定程度上提高动物的免疫功能。饲料中添加牛磺酸在4周时可以提高虹鳟仔鱼的免疫力,但长时间添加(16周)却降低虹鳟的免疫力(徐奇友等,2007)。
3.4 调节渗透压
牛磺酸可以作为生物膜的稳定剂,调节细胞膜的流动性,使其成为理想的渗透压调节物质。牛磺酸渗透调节作用的发挥主要依靠牛磺酸对于渗透调节蛋白ENVZ的修饰作用(Moeukemann 等,1999)。牛磺酸是主要的渗透压调节物质之一。饲喂缺乏牛磺酸饲料的五条鰤(Serio⁃la quinqueradiata),血浆牛磺酸浓度和血清渗透压显著降低(Takagi S等,2006)。研究广盐性的罗非鱼(Sarotherodon mossambicus)发 现,牛磺酸是罗非鱼(Sarotherodon moss⁃ambicus)肌肉和肝脏中的主要氨基酸,当肌肉细胞皱缩时,牛磺酸浓度急剧升高(Assem等,1983)。
3.5 提高繁殖能力
近年来,牛磺酸对于鱼类繁殖性能的影响逐渐受到重视。配制牛磺酸含量分别为0%、0.5%、1%的3种配合饲料投喂五条鰤,结果表明,牛磺酸能够提高五条鰤的产卵量,且随着牛磺酸含量的提高,五条鰤卵细胞生长显著提高,最终测定雌鱼的产卵率分别是0、1/6~6/7(Mat⁃sunari等,2006)。当添加牛磺酸水平为10 g/kg饲料时,罗非鱼亲鱼表现出最好的繁殖性能(El-Sayed,2014)。
3.6 提高动物的耐缺氧能力
牛磺酸能改善鱼类的耐缺氧能力。牛磺酸含量分别为6‰和8‰时均能显著提高黄河鲤鱼抗缺氧能力(P<0.05)(张书松等,2008)。用牛磺酸含量分别为 0、0.5、1、1.5、2、2.5 g/kg的饲料饲喂草鱼稚鱼,随着饲料中牛磺酸含量的增加,草鱼在极度缺氧环境下生存时间延长,并且最高牛磺酸含量有最好的耐缺氧能力(Yang等,2013)。牛磺酸含量至少为1.5 g/kg时红细胞渗透脆性显著提高,当含量上升到2.5 g/kg时红细胞渗透脆性大幅提高。红细胞渗透脆性测试试验中溶血率和极度缺氧条件下的生存时间之间存在明显的相关性(P=0.023<0.05),牛磺酸的细胞膜稳定性提高了草鱼稚鱼极度耐缺氧能力(Yang等,2013)。牛磺酸通过提高鲫鱼的血红蛋白含量提高鲫鱼耐缺氧能力(邱小琮等,2006)。当牛磺酸浓度为1.5%时麦穗鱼存活时间最长(吕广祺等,2015)。
4 牛磺酸在鱼类配合饲料中的适宜需求量
各种鱼类的牛磺酸合成能力不同,同种鱼类不同生长阶段牛磺酸合成能力变化也很大。此外,饲料原料组成、蛋白源、养殖鱼类的生长阶段及其所处的环境条件也显著影响牛磺酸的适宜需求量。近几年有关鱼类牛磺酸的需求量研究结果见表1。
5 牛磺酸在鱼类配合饲料中的应用
5.1 牛磺酸在鱼类配合饲料中的应用现状与存在的问题
近年来,鱼粉资源的短缺,导致其价格持续攀升,推动了植物源蛋白替代鱼粉的研究。植物源蛋白除存在营养素不均衡、缺乏限制性氨基酸、抗营养因子、适口性差等不足外,绝大多数植物源蛋白均缺乏牛磺酸,因此,研究植物性蛋白源替代鱼粉时更需要密切关注牛磺酸的应用,特别是在肉食性鱼类饲料中的应用。
研究表明,牛磺酸在某些鱼体中不能合成或者合成量不足,为条件性必需氨基酸。其中半胱亚磺酸脱羧酶(CSD)为含硫氨基酸转化生成牛磺酸的最大限制酶,因此CSD在鱼体内的活性在一定程度上代表了鱼类合成牛磺酸的能力。但是CSD活性在不同鱼类及同种鱼的不同发育时期差异显著:CSD活性相对较高的鱼类,虹鳟、大西洋鲑鱼、罗非鱼等;CSD活性相对较低的鱼类,真鲷、牙鲆、鲤鱼、金枪鱼等;CSD几乎没有活性的鱼类,黄尾鱼、鲽鱼(Pleuronichthys cornutus)等(于道德等,2010)。
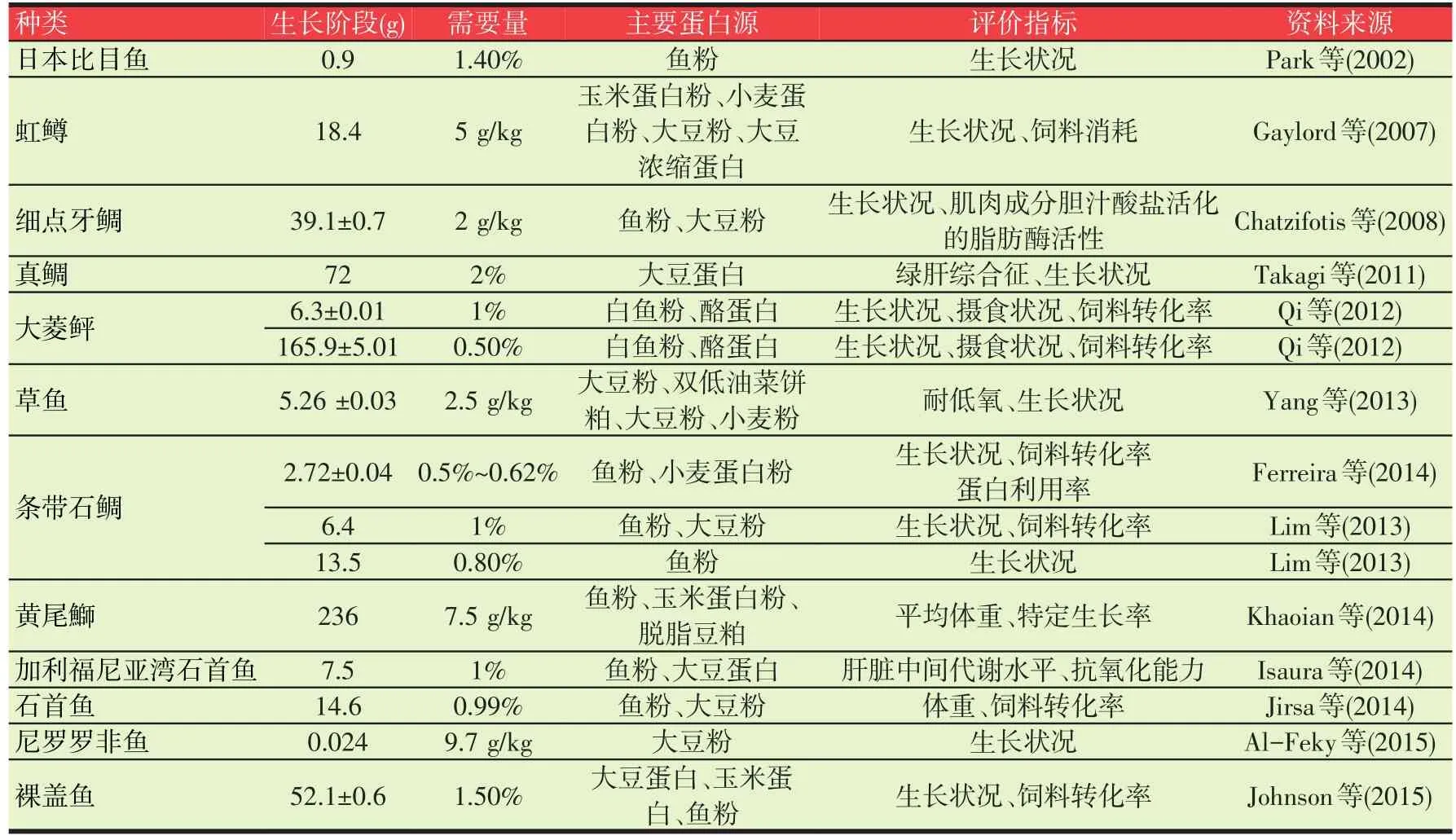
表1 鱼类牛磺酸的需要量
牛磺酸作为一种重要的鱼类饲料添加剂,其应用已有较多的报道,取得了良好的效果:①作为诱食剂。牛磺酸作为鱼类饲料添加剂能够显著提高鱼类的摄食率;②作为营养强化剂。随着饲料中牛磺酸含量的增加,五条鰤肌肉中牛磺酸含量成比例增长,且试验最初3周时添加牛磺酸的五条鰤稚鱼的生长性能显著提高,在3~6周期间补充牛磺酸的五条鰤重量得到提高(Matsunari等,2005);③作为植物源蛋白替代动物源蛋白的饲料添加剂。以高玉米蛋白粉为基础饲料中,添加1%的牛磺酸能够显著促进军曹鱼的摄食、生长以及饲料利用率(骆艺文等,2013)。以大豆蛋白作为全部蛋白源,补充牛磺酸能提高虹鳟的生长、饲料转化率、蛋白储积率、能量储积率(Gaylord等,2007)。以五条鰤为试验对象发现,以大豆浓缩蛋白完全替代鱼粉,则饲料中添加牛磺酸对于维持五条鰤正常的生理条件和生长状况是必要的(Takagi S等,2008);④增强鱼类的生理功能。牛磺酸是牙鲆中与胆酸结合的唯一氨基酸,结合形成牛磺胆酸和牛磺鹅脱氧胆酸,进而与胆汁结合进入消化道促进脂肪等的消化吸收(Kim等,2007);⑤提高鱼类的生存能力。真鲷缺乏牛磺酸会发生绿肝综合征,因为牛磺酸缺乏引起红细胞渗透脆性增加,导致红细胞破裂,逸出血红蛋白,血红蛋白的降解使胆色素大量生成,不能从肝脏排泄到胆囊,最终导致胆色素淤积(王和伟等,2013)。真鲷饲喂大豆蛋白和玉米蛋白代替鱼粉的饲料,发现这些真鲷的绿肝综合征发病率提高,推测肝脏牛磺酸含量下降是引发真鲷绿肝综合征的原因之一(Goto等,2001);⑥改善鱼肉品质。饲料中添加牛磺酸能够减少大西洋鲑鱼幼鱼脂肪的积累,同时不影响蛋白质的增加,并且饲料中添加牛磺酸能够增加肝脏中的自由氨基酸含量(Espe等,2012)。
目前鱼类对牛磺酸需求量的研究还远远不能满足养殖需求,许多研究结果只是说明某种鱼某一阶段对牛磺酸的需要量。此外,过量牛磺酸添加导致鱼类出现中毒现象。饲料中牛磺酸的添加水平不仅要考虑鱼的种类、生长阶段,还要注意饲料蛋白源及其水平、含硫氨基酸的种类和数量,同时饲料中的牛磺酸转运拮抗剂(如脒基牛磺酸、β-丙氨酸)会降低牛磺酸的使用效果(王和伟等,2013)。
5.2 牛磺酸在鱼类配合饲料中的应用前景
海洋渔业资源逐渐减少,导致鱼粉价格不断上升,这已成为水产养殖业发展的瓶颈,植物蛋白替代鱼粉已成为水产配合饲料的研究热点,鱼类特别是肉食性鱼类在植物性蛋白源替代鱼粉等动物性蛋白源时,添加牛磺酸对鱼的生长有促进作用。牛磺酸的应用在当今鱼粉紧缺的情况下前景越来越广阔。但相关基础研究还缺乏,需要加大相关研究,特别是不同鱼类种类、不同阶段以及不同鱼类配合饲料配方中的牛磺酸需求量研究,以推进其应用,促进鱼类健康养殖产业的发展。
6 结语
牛磺酸虽然结构简单,却在生物体内发挥重要作用。目前牛磺酸在哺乳类动物中的作用研究得较为全面,而在鱼类中的功能研究得相对较少。已知鱼类缺乏牛磺酸会导致自身生长状况下降、存活率降低、抵抗力下降、生殖能力受到影响等,在真鲷等种类中会增加患绿肝综合征的几率。一方面,牛磺酸在生物体内发挥作用的机理还有待进一步研究,牛磺酸作为水产配合饲料添加剂的用量还需进一步完善,同时牛磺酸与其他营养物质的代谢关系还需要进一步明确。另一方面,牛磺酸具有分布广泛、天然无害、功能全面等特点,相信一定会在促进水产养殖健康、可持续发展中发挥重要作用。
