临安毛竹快速生长期耗水特性及其影响因子*
2015-01-17蔡先锋李洪吉袁佳丽曾莹莹温国胜
蔡先锋,李洪吉,袁佳丽,曾莹莹,温国胜
(1.浙江农林大学林业与生物技术学院,浙江 临安311300;2.浙江农林大学亚热带森林培育国家重点实验室培育基地,浙江 临安311300)
毛竹 (Phyllostachys edulis)属单子叶植物禾本科(Poaceae)簕竹族(Bambuseae)刚竹属(Phyllostachys)植物[1],目前毛竹林面积约270×104hm2[2],是最广泛使用的竹种[3]。毛竹作为竹类植物在保持水土、维持环境碳氧平衡等方面作用明显。毛竹因投资小、收益快、效益高等特点,是实现林业可持续发展和解决农村贫困的一个有效途径,被称为“农民的山中银行”[4]。
水分是植物生长发育必需的资源,是植物进行光合作用必要的元素。光合作用与蒸腾作用同时进行,两者的比值反映了水分利用效率的大小[5~6]。水分利用效率 (WUE)作为植物生理活动过程中消耗水形成有机物质的基本效率,成为确定植物生长发育过程中所需要的水分供应的最佳指标之一[7]。目前对植物WUE及Tr研究主要集中分析其日变化方面,而涉及到整个生长期WUE宏观变化及其影响因子方面的研究鲜见报道。本项研究通过分析不同竹龄毛竹在不同生长期WUE和Tr动态及其影响因子,试图从水分利用的角度深刻理解毛竹快速生长的机制,为揭示毛竹的固碳机制和制定科学合理的管理措施提供理论依据。
1 材料与方法
1.1 研究区概况
试验地为浙江农林大学毛竹生理生态定位监测站,位于浙江省临安市东部的青山镇研口村 (北纬30°14',东经119°42')。当地气候温暖湿润,属亚热带季风气候,年均气温8.5~15.2℃,最冷月气温 2.1 ~3.0℃,最热月气温 19.5 ~27.6℃,无霜期209-235天。土壤呈酸性,pH值为4.7~6.0。属低山丘陵区,土壤属红壤,土层深度均在60 cm以上。试验林为复垦纯毛竹林,分大小年经营,竹龄在1-5年,毛竹的高度范围在9~13 m,胸径在8~16 cm。
1.2 方法
毛竹一般用度来定义其竹龄,度的划分根据竹子换叶来界定[8]。新竹长成要在5月份,将2012年出笋的新竹在未换叶之前定为Ⅰ度竹,未换叶的2010年的竹子定义为Ⅱ度竹,未换叶的2008年的竹子定义为Ⅲ度竹,在毛竹林样地上沿上、中、下随机选取3个10 m×10 m的样方,统计每个样方中所有毛竹胸径、竹高、枝下高、冠幅,计算出标准竹的胸径为93 mm,竹高为13 m,枝下高为6.5 m,冠幅为2.3 m。
在毛竹爆发式生长期 (3-5月),每月中旬(天气晴朗),利用Li-6400XT便携式光合测定仪测定净光合速率 (Pn)、蒸腾速率 (Tr)、光合有效辐射 (PARi)、大气温度 (Ta)、大气 CO2浓度(Ca)、胞间 CO2浓度 (Ci)、大气相对湿度(RH)、气孔导度 (Cond)。在3株不同年龄的标准竹上选取中上部无遮挡、生长状况良好且完整的叶片,在 12个不同光强 (0,10,30,50,80,100,300,500,800,1 000,1 200,1 500)下测定4个重复,总共3×3×12×4=432组数据。计算公式为,WUE=Pn/Tr
1.3 统计分析
运用SPSS 19.0统计分析软件对数据进行相关性分析,Excel 2003制图。
2 结果与分析
2.1 不同生长期不同竹龄WUE及Tr动态分析
不同生长期WUE变化呈“中间高,两边低”的趋势 (图1),即毛竹WUE在生长中期最大,前期稍弱,后期最小;而不同生长期Tr则呈“中间低,两边高”趋势,即毛竹Tr在生长中期最低,前期其次,后期最大,WUE及Tr变化趋势相反。
在相同生长期内,WUE基本上是Ⅱ度竹最大,Ⅰ度竹稍次,Ⅲ度竹最低。这应该是因为Ⅰ度竹各器官发育尚不成熟,水分吸收及输送能力不足,导致WUE稍弱,而Ⅲ度竹则相反,各器官老化,导致其WUE能力下降,Ⅱ度竹各器官发育完善,从而WUE最大;Tr在中后期基本上是随着竹龄的增加而逐渐减小的,即Ⅰ度竹最大,Ⅱ度竹其次,Ⅲ度竹最小。

图1 不同生长期不同竹龄WUE及Tr变化Fig.1 Changes of different ages and different growth on WUE and Tr
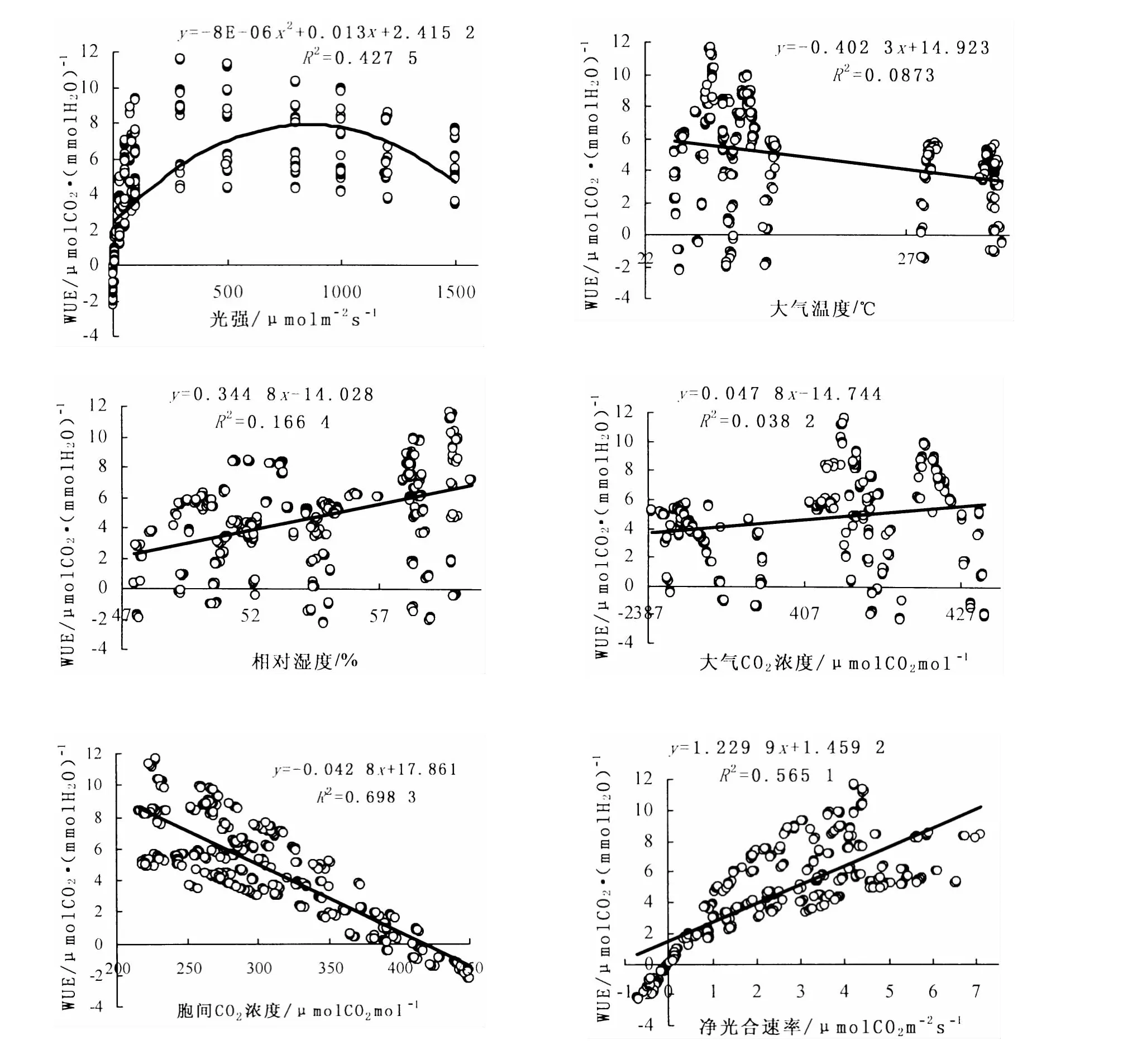
图2 WUE与环境因子关系Fig.2 Relationship between WUE and environmental factors
2.2 WUE的影响因子分析
在0~800光强下,WUE随着光强的增强逐渐增大 (图2)。约在800光强下达到最大值,随后随着PARi的继续增加,WUE则减小,主要原因则是叶片本身光合酶活性随着光强的增强而受到抑制,导致Pn下降,进而使WUE减小;WUE与Ta具有较弱的负相关关系;WUE与RH、Ca呈显著正相关,随着RH及Ca的逐渐增大,WUE逐渐增大;WUE与Ci呈显著负相关,主要是因为在Pn增大时,WUE增加而Ci减小所致;Pn作为反映WUE变化的重要指标之一,显著影响了WUE的变化,表现为WUE随着Pn的增大而明显增大。
2.3 Tr的影响因子分析
Tr与PARi、Ta、Cond呈正相关,与 Ca、Ci、RH呈负相关 (图3)。PARi对Tr作用明显,随着PARi的逐渐增强,Tr逐步增大,这与WUE变化趋势不同,说明0~800光强下Pn及Tr同时增大,且Pn增幅比Tr明显,而在800光强时Pn达到最大值,随着光强的继续增强,Pn逐渐减小,Tr则继续增大,导致WUE减小,从而有WUE呈先增大后减小的趋势;在快速生长期,Ta主要集中在22~29℃,温度波动不剧烈,对Tr影响较为微弱;Cond作为控制叶片水气交换的通道,其开度变化直接影响到Tr的大小,Tr随着Cond的逐渐增大显著增大;Tr与Ca呈负相关,主要原因是气孔开度减小引起的水分蒸发的大幅度减小,导致Tr的减小;Tr随着Ci的逐渐增大而逐渐减小,说明Ci的增大是由气孔开度减小导致的,气孔开度减小,Tr必然减小;Tr受RH影响较大,呈先增后降变化趋势,这主要是因为随着RH逐渐增大,叶片气孔开度逐渐减小,导致Tr减小。

图3 Tr与环境因子关系Fig.3 Relationship between Tr and environmental factors
2.4 WUE及Tr与影响因子相关性分析
由表1可知,WUE与Pn、PARi、Ci、Ta、Ca、RH、Cond呈极显著相关,且与 PARi、Ca、RH、Cond呈正相关,与Ci、Ta呈负相关;Tr与PARi、Ta、Cond呈极显著正相关,与Ca、RH、Ci呈极显著负相关。这与图1、图2散点图结果一致。

表1 WUE及Tr与影响因子相关性分析Tab.1 Correlation analysis between WUE,Tr and impact factors
3 结论与讨论
3.1 结论
(1)毛竹WUE在快速生长中期最高,前期其次,后期最小;Tr在快速生长中期最小,前期其次,后期最大。在相同生长期内:Ⅱ度竹WUE最大,Ⅰ度竹其次,Ⅲ度竹最差;而Tr随着竹龄增加逐渐减小。
(2)毛竹在快速生长过程中 WUE、Tr与PARi、Ta、Ca、Ci、RH、Cond都呈极显著相关。WUE主要影响因子是PARi及Ci,且与前者呈正相关,与后者呈负相关;Tr主要影响因子是Cond、PARi、Ci及Ca,且与PARi及Cond呈正相关,与Ci及 Ca 呈负相关。这与龙明秀[9]、赵英等[10]的研究结果一致。
3.2 讨论
水分是植物生长发育的主要因素之一。水分利用率可反映植物生长与水分利用之间的关系,常为引进、选育高水分利用率的植物种类品种提供理论依据和筛选指标[11]。叶片水分利用率在一定程度上反映了植物的耗水性和抗旱性[12]。植物吸收的水分有大部分消耗在叶片蒸腾作用上。因此,如何提高水分利用效率和将无效耗水变为有效蒸腾关系着植物抗旱性、成材性的提高。毛竹快速生长期是毛竹生长发育中极其关键的时期,分析不同竹龄毛竹WUE、Tr及其影响因子对于了解它们在毛竹林分生态系统中的作用、固碳机制和水分输送动态具有十分重要的意义,同时重点研究WUE、Tr与各生态因子之间的关系,有利于促进农林复合经营管理体系的提高,为培养高WUE作物提供科学理论指导。
该试验的不足之处在于缺乏对快速生长不同时期WUE及Tr日变化的同步测定,同时对于新竹长成成竹后的全年变化应进行进一步跟踪测定。Vertessy等[13]认为Tr受植物本身生态结构和环境因子多方面影响;Osmond等[14]认为环境蒸汽压变化对Tr有较大影响;Morecroft等[15]认为叶温对植物WUE有影响。因此关于其他因子如蒸汽气压亏缺及叶温对WUE和Tr的影响有待进一步研究。
[1]郭起荣,杨光耀,杜天真,等.毛竹学名百年之争[J].世界植物通讯,2006,4(1):18-20.
[2]江泽慧.在第四届中国竹子之乡联谊会暨全国竹(藤)业经济发展研讨会开幕式上的致辞[J].竹子研究汇刊,2000,19(3):2-3.
[3]林福兴,林如青,林强.不同绿竹种源含水率与持水量研究[J].水土保持应用技术,2007(2):6-8.
[4]胡小洲.毛竹林发展前景及其丰产栽培综合措施[J].安徽农学通报,2013,19(8):105-106.
[5]李俊,于沪宁,刘苏峡.冬小麦水分利用效率及其环境影响因素分析[J].地理学报,1997,52(6):551-560.
[6]张岁岐,山仑.植物水分利用效率及其研究进展[J].干旱地区农业研究,2002,20(4):1-5.
[7]王会肖,刘昌明.作物光合、蒸腾与水分高效利用的试验研究[J].应用生态学报,2003,14(10):1632-1636.
[8]葛宏立,周国模,刘恩斌,等.浙江省毛竹直径与年龄的二元 Weibull分布模型[J].林业科学,2008,44(12):15-20.
[9]龙明秀.紫花苜蓿光能及叶片水分利用效率影响因子分析[J].草业科学,2009,26(11):73-77.
[10]赵英,张斌,赵华春,等.农林复合系统中南酸枣蒸腾特征及影响因子[J].应用生态学报,2005,16(11):2035-2040.
[11]梁宗锁,康绍忠.植物水分利用率及其提高途径[J].西北农林学报,1996,16(6):79-84.
[12]李国泰.8种园林树种光合作用特征与水分利用效率比较[J].林业科学研究,2002,15(3):291-296.
[13]Vertessy R A,Benyon R G,Sullivan S K,et al.Relationships between stem diameter,sapwood area,leaf area and transpiration in a young mountain ash forest[J].Tree Physiol,1995,15:559-567.
[14]Osmond C B,Björkman Ö.Simultaneous measurement of O2effects on net photosynthesis and glycolate metabolism in C3 and C4 species of Atriplex[J].Carnegie Institution of Washington Yearbook,1972,77:141-148.
[15]Morecroft M D,Woodward F I.Experimental investigations on the environmental determination of δ13C at different altitude[J].Journal of Experimental Botany,1990,41(231):1303-1308.
