云南榧树群落及其植物区系特征研究*
2015-01-17侯昭强文光玉周丹李敏敏杜凡
侯昭强,文光玉,周丹,李敏敏,杜凡
(西南林业大学,云南 昆明650224)
云南榧树 (Torreya yunnanensis)属红豆杉科(Taxaceae)榧树属 (Torreya)植物,为滇西北特有种。榧树属 (共7种,我国产4种)所有种在中国均被列为国家二级重点保护野生植物[1],云南榧树还被《中国物种红色名录》[3]确定为濒危种。
珍稀濒危植物云南榧树无论在植物系统分类学研究、种质资源方面,还是在用材、经济、科学、文化、教育等方面都具有重要价值。因此,保护这一珍稀濒危植物具有深远的意义。而目前针对该物种的相关研究较少,且多集中于云南榧树组成成分等分子层面的研究[4~6]。所以,本项研究通过实地调查,从群落学特征、物种组成以及区系特征等方面研究云南榧树群落的生态学特性,为更好地保护该地区云南榧树种质资源提供科学依据。
1 研究地概况
研究区云南榧树群落地处东经99°03'~99°23'、北纬27°01'~27°23',海拔 2 160 ~2 970 m 的石灰岩地带,坡度20°~45°,以海拔2 300~2 800 m生长最集中。
研究区位于滇西北的维西县和剑川县。参照两县城所在地气象资料,维西县城海拔2 320 m,年日照时数2 071.3 h,年平均气温11.3℃,极端低温 -6.8℃,极端高温 31.7℃,年降水量 957.5 mm,全年无霜期253天。剑川县城海拔2 200 m,年日照时数2 405.5 h,年平均气温12.2℃,极端低温-10.7℃,极端高温33.5℃,年降水量738.0 mm,全年无霜期256天[7]。研究区域位于林区,总体海拔比所在县城海拔高,因而其降雨量会稍高于县城降雨量,温度略低于县城温度。
2 研究方法
2.1 资料收集及现场调查
全面收集整理以往调查资料,将云南榧树分布点标记在地形图上。2010年3月-2013年10月,采用典型选样法设置了面积为400 m2的16个样方。记录各样方的坡位、坡向、海拔、人为影响、群落外貌和生境特征。并将群落中胸径5 cm以上的木本植株作为乔木层,进行每木调查,分别记录其种类、胸径、树高、冠幅、物候;灌木层 (包括胸径5 cm以下乔木幼树)记录种类、株 (丛)数、高度、盖度;草本层及层间植物记录种类、数量、盖度 (表1)。
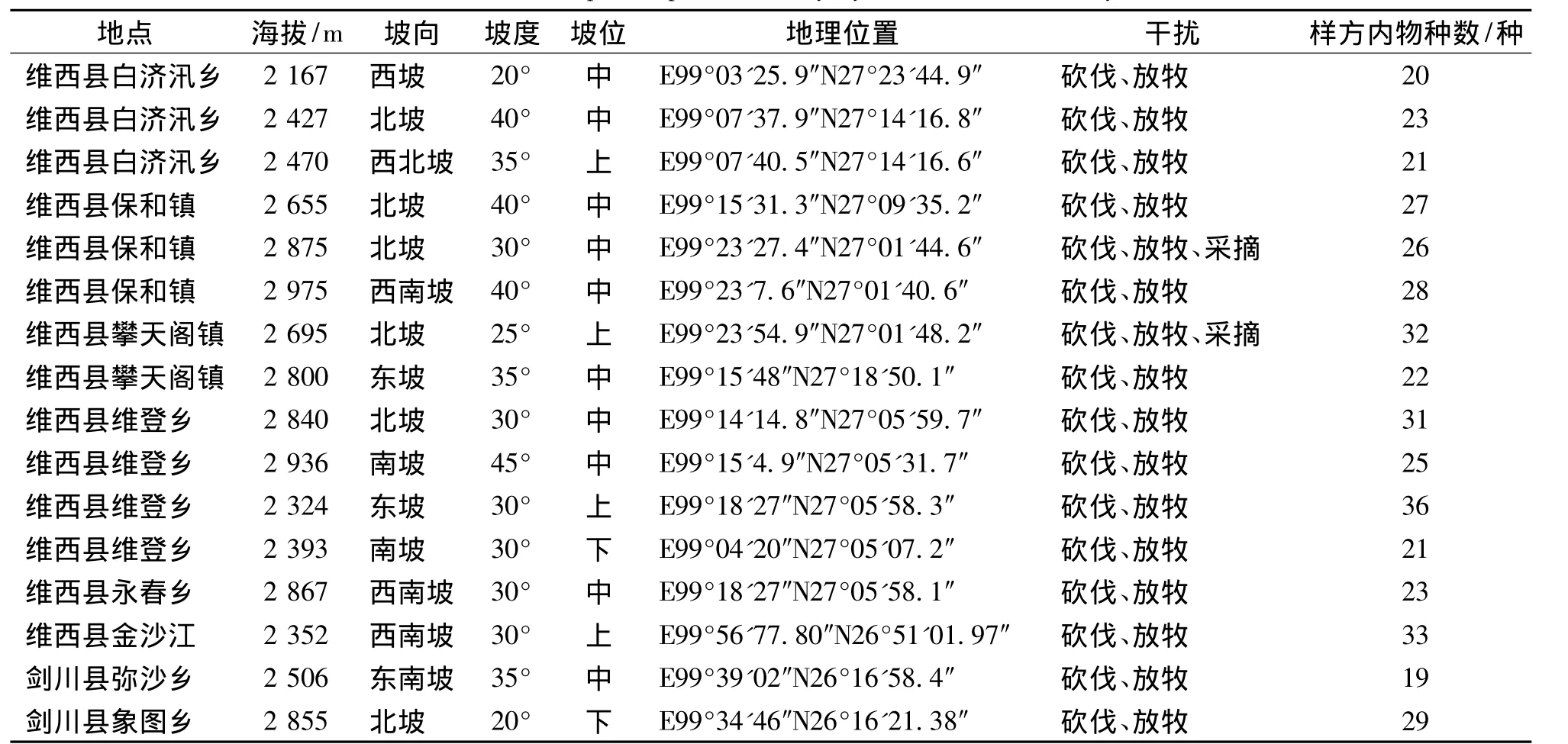
表1 云南榧树群落样地概况Tab.1 The profile plots of Torreya yunnanensis community
2.2 数据分析
按照 《中国植物志》[8]、 《云南植物志》[9]、“Flora of China”鉴定样方内所采集的标本;植物生活型谱参照 Raunkiaer的分类等级划分[10~11];属的分布区类型按照吴征镒院士的方法划分[12~13];区系次生度按照杜凡[14]提出的模型计算。
区系次生度 (F) F=a/(b+a),式中,F为区系次生度,a代表进入系统中的次生种的种数,b代表系统中保存的原生种的种数。
次生种的确定 指通常分布于村寨边、路边、林缘等人为活动频繁的环境中的阳性树种,它们不出现在没有人为干扰的原生植物群落中。
区系次生度指人为干扰后原生态系统中进入的次生种及已丧失的原生种的种类的百分比,人为干扰强度越大该比值越高,因而该数值的大小能够反映一个生态系统 (或生境)受人为干扰的程度。
3 结果与分析
3.1 云南榧树群落学特征
从生境上看,云南榧树基本分布在北坡的中上部位,沟底和山脊处较为少见,生境较湿润、稍陡、裸岩多,土壤以黄壤和棕壤为主,林下枯落物厚2~3 cm。云南榧树所处的群落属于针-阔混交林。群落的乔木层盖度50% ~70%,高6~16 m,组成种类计48种,以云南榧树 (Torreya yunnanensis)为优势,伴生种主要是青冈 (Cyclobalanopsis glauca)、光叶高山栎 (Quercus pseudosemecarpifolia)、云南铁杉 (Tsuga dumosa)、栓皮栎 (Quercus variabilis)、漆 (Toxicodendron vernicifluum)、云南红豆杉 (Taxus yunnanensis)等;灌木层盖度20%~45%,高1~4.5 m,组成种类计68种,主要有粉枝莓 (Rubus biflorus)、宝兴茶藨子 (Ribes moupinense)、茸毛木蓝 (Indigofera stachyodes)等;草本层盖度35% ~50%,高0.01~0.6 m,种类丰富,计116种,常见天名精 (Carpesium abrotanoides)、粘冠草 (Myriactis wightii)、细穗兔儿风(Ainsliaea spicata)、蟹甲草 (Parasenecio forrestii)等;层间植物26种,高1~5 m,盖度25% ~30%,如裂叶铁线莲 (Clematis parviloba)、崖爬藤 (Tetrastigma obtectum)、云南崖爬藤 (Tetrastigma yunnanense)等。
3.2 种群结构
16个样方中记录到云南榧树92株 (丛),单个样方最多达14株。将92株云南榧树按上限排外法以5 cm为1个径级分为11个径级 (表2)。

表2 云南榧树径级结构Tab.2 The diameter-class structure of Torreya yunnanensis
从表2可知,云南榧树以胸径超过50 cm的植株最多,共68株,占总株数的73.91%;胸径46~50 cm的次之,共4株,占4.35%;胸径小于等于5 cm的幼树仅7株,占7.61%。群落中胸径最大的植株胸径超过150 cm,最小植株高仅20 cm。云南榧树种群成年大树所占比例较大,且大径级植株多为断头或中空等生长状况较差的植株,而中幼体植株比例较小,仅见7株幼苗均生长于石缝间,可见该种群属于衰退型种群。
3.3 种子植物区系地理成分分析
3.3.1 科属种组成
组成云南榧树群落的物种较多,6 400 m2的样方面积内记录到维管束植物90科199属258种;其中蕨类植物13科18属24种,裸子植物3科8属9种,双子叶植物65科144属190种,单子叶植物9科29属35种 (表3)。
各样地乔木层树种重要值大于2的种类统计见表4。群落中有345株乔木树种,其中云南榧树85株,重要值15.58%,在所有乔木树种中数值最大,属于乔木层的优势种。青冈、华山松、光叶高山栎和云南铁杉的重要值依次减小,共同构成群落的建群种;团香果 (Lindera latifolia)、聚花桂(Cinnamomum contractum)、云南泡花树 (Meliosma yunnanensis)、五角枫 (Acer mono)、杨叶木姜子(Litsea populifolia)等在群落中少见,属于偶见种(表4)。

表3 云南榧树群落物种组成Tab.3 The species composition of Torreya yunnanensis
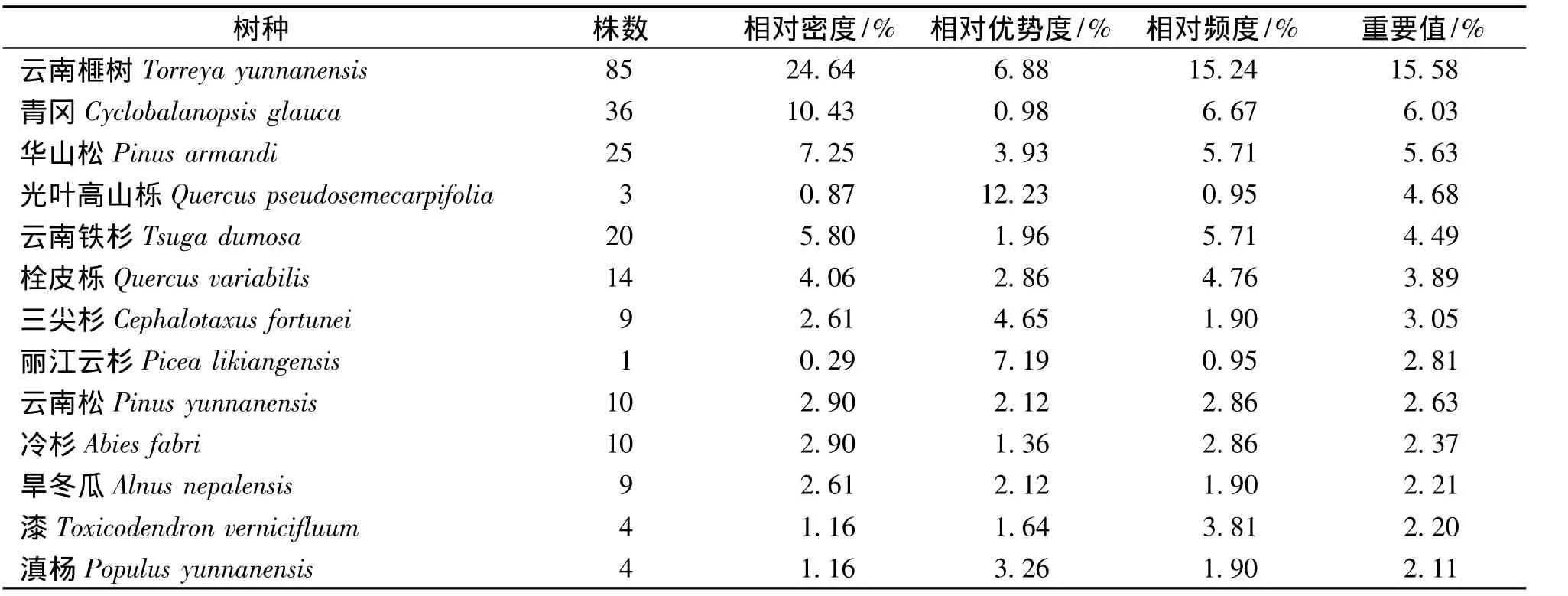
表4 群落中乔木层主要树种重要值Tab.4 The important community values of the Main tree species
3.3.2 属的分布区类型分析
按照吴征镒院士的方法[12],将群落中种子植物属划分为17个类型及亚型 (表5)。
世界分布的属有剪股颖属 (Agrostis)、银莲花属 (Anemone)、蓼属 (Polygonum)、鬼针草属(Bidens)、远志属 (Polygala)等18属,占总数的9.94%。此类型所含物种多为适应性较强的杂草。
泛热带分布的属有十大功劳属 (Mahonia)、马兜铃属 (Aristolochia)、草胡椒属 (Peperomia)等26属,占总属数 (不包括世界分布,下同)的15.95%;热带亚洲至热带非洲及其变型分布有宿苞豆属 (Shuteria)、蝎子草属 (Girardinia)等10属,占总属数的6.13%;热带亚洲分布及其变型有青冈属 (Cyclobalanopsis)、松属 (Pinus)、栎属(Quercus)、野扇花属 (Sarcococca)等12属,占总属数的7.36%。
北温带分布及其变型有桤木属 (Alnus)、杉属(Abies)、松属 (Pinus)、丽江云杉属 (Picea)、栎属 (Quercus)、小蘖属 (Berberis)等56属,占总属数的34.36%;东亚和北美洲间断分布有榧树属(Torreya)、漆树属 (Toxicodendron)、铁杉属(Tsuga)、楤木属 (Aralia)等13属,占总属数的7.98%;旧世界温带分布有淫羊藿属 (Epimedium)、荞麦属 (Fagopyrum)、瑞香属 (Daphne)等9属,占总属数的5.52%;东亚分布及其变型有溲疏属 (Deutzia)、旌节花属 (Stachyurus)、十萼花属 (Dipentodon)、棣棠花属 (Kerria)等20属,占总属数的12.27%。其中中国-喜马拉雅分布5属,占总属数的3.07%;中国-日本分布有刺楸属 (Kalopanax)、半夏 (Pinellia)3属,占总属数的 1.84%。
中国特有分布属仅箭竹属 (Fargesia)和短檐苣苔属 (Tremacron)2属,占总属数的1.11%。箭竹属主产喜马拉雅和横断山区。在我国北自祁连山东坡,南达海南,东起赣、湘,西迄西藏吉隆,以云南的种类最为丰富。短檐苣苔属特产我国云南西北至东部以及四川西南部,主要分布于海拔1 500~2 800 m的阴湿岩石上。
综上,群落中各类热带地理成分共有59属,占总属数的36.20%;各类温带地理成分共有104属,占总属数的63.80%,反映出了云南榧树群落强烈的温带性质。温带分布属中又以北温带分布为主,有56属,且构成云南榧树群落的主要乔木树种云南榧树、云南铁杉、冷杉等均属于北温带分布,说明该群落在植物区系上与北温带区系联系更为紧密。
3.3.3 种的分布区类型分析
根据每个种的现代地理分布格局,参照吴征镒院士的方案[13],将群落中的维管植物分为14种类型及变型 (表5)。
世界分布种8种,占总数的3.10%,如柳叶菜 (Epilobium hirsutum)、小画眉草 (Eragrostis minor)、扭黄茅 (Heteropogon contortus)等,大部分为阳性次生成分的世界性杂草。与属相比,世界广布种的比重有所下降。

表5 云南榧树群落种子植物属、种分布区类型Tab.5 The distribution types about seed plant in genus,species of Torreya yunnanensis
热带分布 (2-7)25种,占总种数的9.69%。其中泛热带分布3种,占总数的1.16%,如豆瓣绿 (Peperomia tetraphylla)、鬼针草 (Bidens pilosa)、糯米团 (Gonostegia hirta);热带亚洲至热带大洋洲分布2种,占总种数的0.78%,包括密毛蕨 (Pteridium revolutum)、金粉蕨 (Onychium siliculosum);热带亚洲至热带非洲分布3种,占总属数的1.16%,如十字苔草 (Carex cruciata)、茅叶荩草 (Arthraxon prionodes);热带亚洲分布25种,占总属数的 9.69%,如五叶草 (Geranium nepalense)、茸毛木蓝 (Indigofera stachyodes)、阔叶鳞盖蕨Microlepia platyphylla)等。
温带分布 (8-14)共217种,占总种数的84.11%;其中北温带分布 7种,占总种数的2.71%,如乔木树种光叶高山栎、栓皮栎,假升麻 (Aruncus sylvester)、野草莓 (Fragaria vesca)、香薷 (Elsholtzia ciliata)、毛葡萄 (Vitis heyneana)等;旧世界分布6种,占总种数的2.33%,如天名精 (Carpesium abrotanoides)、野青茅 (Deyeuxia arundinacea)、苦苣菜 (Sonchus oleraceus)等;温带亚洲分布3种,占总种数的1.16%,包括楼梯草 (Elatostema involucratum)、刺楸 (Kalopanax septemlobu)、败酱 (Patrinia scabiosaefolia);地中海区、西亚至中亚分布有3种,包括看麦娘 (Alopecurus aequalis)、小蓟 (Cirsium setosum)、小铁仔(Myrsine africana),占总种数的1.16%;中亚分布4种,如越桔忍冬 (Lonicera myrtillus)、素方花(Jasminum officinale)等,占总种数的1.55%;东亚分布8种,占总种数的3.10%,如千里光 (Senecio scandens)、盐肤木 (Rhus chinensis)、米饭花(Lyonia ovalifolia)等;中国-喜马拉雅分布79种,占总种数的30.62%,如华山松 (Pinus armandii)、云南铁杉、三尖杉 (Cephalotaxus fortunei)、丽江云杉 (Picea likiangensis)、旱冬瓜 (Alnus ne-palensis)、细穗兔儿风 (Ainsliaea spicata)、梾木(Cornus macrophylla)等;中国-日本分布24种,占总种数的9.30%,如半夏 (Pinellia ternata)、青冈 (Cyclobalanopsis glauca)、叶上花 (Helwingia japonica)、乌饭 (Vaccinium bracteatum)、棣棠花(Kerria japonica)等。
该区域属于中国特有分布的种共83种,占总种数的32.17%,有云南榧树、冷杉 (Abies fabri)、云南松 (Pinus yunnanensis)、狭叶五味子(Schisandra lancifolia)、杨叶木姜子 (Litsea populifolia)、沧江新樟 (Neocinnamomum mekongense)、偏翅唐松草 Thalictrum delavayi)、宝兴淫羊藿(Epimedium davidii)等;其中云南特有种8种,仅占总种数的3.10%,有密叶小檗 (Berberis davidii)、怒江十大功劳 (Mahonia salweenensis)、紫花凤仙花 (Impatiens purpurea)、齿叶石楠 (Photinia prionophylla)等。
综上,云南榧树群落植物区系复杂,以温带成分占绝对优势:温带属104个,占总属数的63.8%,温带种217个,占总种数的84.11%,乔木层如云南榧树、云南铁杉、光叶高山栎等;灌木层如壮刺小檗 (Berberis deinacantha)、小叶栒子 (Cotoneaster microphyllus)等;草本层如假升麻 (Aruncus sylvester)、野草莓 (Fragaria vesca)、香薷 (Elsholtzia ciliata)等;层间植物如云南铁角蕨 (Asplenium yunnanense)、高山薯蓣 (Dioscorea kamoonensis)、崖爬藤 (Tetrastigma obtectum)等。温带成分贯穿乔木层级以下各层,热带成分仅在属级分布型中表现出一定的比例 (36.20%),种级热带成分很少 (12.79%),说明云南榧树群落具有强烈的温带性质。中国特有属仅2属,占总属数的1.23%,特有种83种,占总种数的32.17%。
3.4 生活型谱
植物的生活型是指有机体对环境及其节律变化长期适应而形成的一种形态表现,是依据生态适应划分的,而生活型谱反映的是某一地区或某一群落植物与环境,尤其是与气候间的相互关系。
群落中高位芽种数占优势,有83种,占32.17%,其中又以矮高位芽为主,有48种,占18.60%,说明研究区云南榧树群落生活的环境温暖而湿润。在这样温暖而湿润的地区大于30 m的大高位芽为零,说明群落中缺乏高大乔木,由此可以看出群落中的大乔木,如云南榧树、华山松、云南铁杉等缺乏并非不适宜生长在这样的环境,而是遭人为砍伐的结果。地面芽有76种,占29.46%;地上芽植物有51种,占19.77%;地下芽植物有11种,占4.26%;1年生植物37种,占14.34%(表6)。可以看出高位芽和地面芽在群落中都占优势,但二者相比高位芽优势不明显,且高位芽中以矮高位芽为主,表明云南榧树群落明显的温带性质。
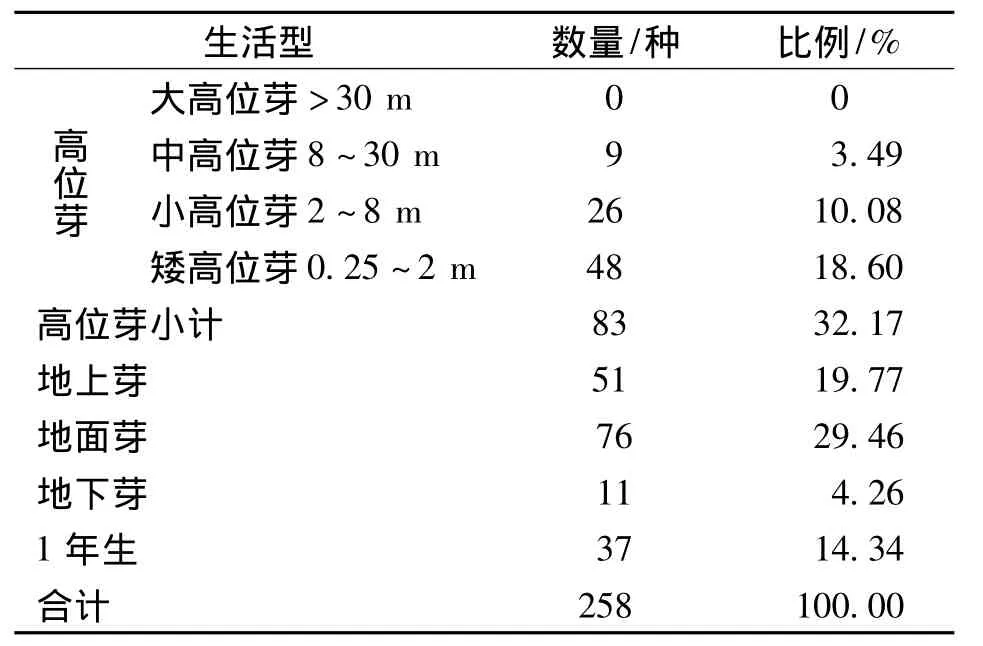
表6 云南榧树群落生活型统计Tab.6 Torreya yunnanensis Plant Community statistics
3.5 区系次生度
区系次生度是通过分析植物群落中次生种(包括外来种)的多少,以及原生种 (土著种)的丧失程度,反映该生态系统中生物多样性受干扰程度。原生种是维持当地植物群落和生态系统稳定性的主要功能群;次生种是从一个生态系统进入另一个生态系统的种类[12],属于林缘的阳性树种即外来入侵种。在植物群落受到砍伐等人为影响后,群落郁闭度降低,有利于阳性物种及外来物种的入侵和生长。按次生种的确定方法确定云南榧树群落次生种116种,占绝对优势 (表7)。
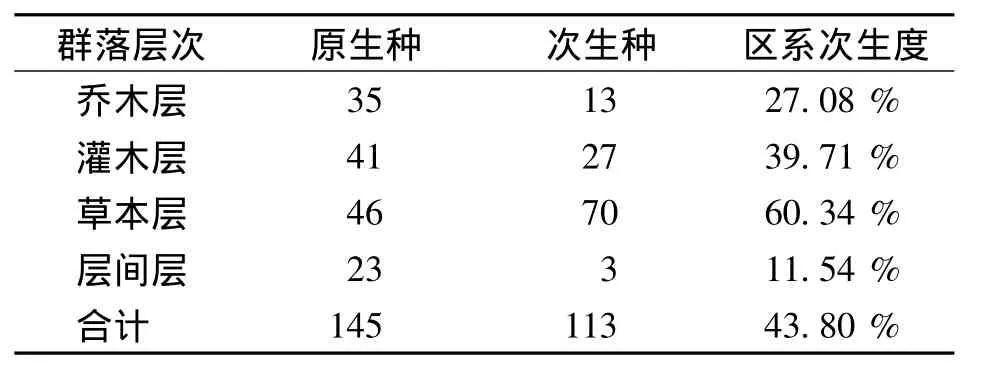
表7 原生种、次生种组成及区系次生度Tab.7 The flora secondary index in native species and secondary species
乔木层有35种原生种,如云南榧树、云南铁杉、青冈、齿叶石楠 (Photinia prionophylla)等。次生种有13种,如枫杨 (Pterocarya stenoptera)、滇杨 (Populus yunnanensis)、毛叶合欢 (Albizia mollis)等。区系次生度为27.08%。
灌木层中原生种41种,如中甸十大功劳 (Mahonia bracteolata)、纤细雀梅藤 (Sageretia gracilis)、短萼海桐 (Pittosporum brevicalyx)、梁王茶 (Nothopanax delavayi)等。次生种有27种,如红刺悬钩子(Rubus rubrisetulosus)、金丝桃 (Hypericum forrestii)、粉枝莓、垂序木蓝 (Indigofera pendula)、序叶苎麻(Boehmeria clidemioides var.diffusa)等。区系次生度为 39.71%。
草本层原生种有46种,如狭叶凤尾蕨 (Pteris henryi)、狗脊蕨 (Woodwardia japonica)、长根金星蕨 (Parathelypteris beddomei)、狗脊蕨 (Woodwardia japonica)、杏叶茴芹 (Pimpinella candolleana)等。次生种有70种,最优势的是粘冠草、野棉花(Anemone vitifolia)、獐牙菜 (Swertia bimaculata)、荩草 (Arthraxon hispidus)等。区系次生度为60.34%。
层间层原生种有23种,如棕鳞瓦韦 (Lepisorus scolopendrum)、石韦 (Pyrrosia lingua)、藏木通 (Aristolochia griffithii)、云南清风藤 (Sabia yunnanensis)、高山薯蓣 (Dioscorea kamoonensis)等。次生种有3种,紫金龙 (Dactylicapnos lichiangensis)、裂叶铁线莲、大花花椒 (Zanthoxylum macranthum)。区系次生度为11.54%。
云南榧树群落草本层次生度较为明显,其次是灌木层,再次是乔木层,层间层也有一定的次生性。群落次生度为43.80%,与同区滇西北部迪庆藏区受藏族宗教采集利用影响的天然侧柏林(27.10%)[13]相比较,明显偏高。区系次生度较高的比值反映出云南榧树群落受人为干扰程度大,原生种面临外来种挑战的情况不容乐观。
4 结论与讨论
研究地区为滇西北高原,属于青藏高原和横断山区的南部区域。地理区位决定了种子植物属和种的分布区类型以温带成分为主要特色。温带成分属104属,占总属数的63.80%,远高于全国同类成分40.87%的比例[15];种的温带成分更加显著,有217个种,占该区总种数的84.11%。所有温带成分种中,以中国特有分布占多数,83种,占总种数的32.17%,其次是中国-喜马拉雅分布种,为79种,占总种数的30.62%,反映了该区与喜马拉雅地区的密切联系。
滇西北是中国三大特有植物起源和分化的中心之一[16],而地处该区的云南榧树群落中国特有属仅2属,种仅83种,占对应比例分别为1.10%、32.17%,特有性偏低,与《云南碧塔海自然保护区》[17]特有属 (21属,占我国特有属的8.64%)、种 (1 232种,占保护区总种数的54.2%)相比相差较大,也是中国种子植物特有属、种高度集中的3个中心地区之一的“川西-滇西北中心”(101属,占我国特有属243属的41.56%)[16]偏低。原因可能是,(1)“川西-滇西北中心”特有种中大部分为草本 (草本属65属,约占64%)[17],本研究调查的群落类型仅为云南榧树群落,没有涉及到草本植物较丰富的草甸、稀树灌草丛等类型。(2)与滇西北中心区域范围[18]相比,云南榧树群落区域很狭窄;(3)砍伐、践踏、放牧等人为活动已经对云南榧树群落的物种组成带来了很大影响;(4)云南榧树群落的物种组成以草本最多,而草本又是最易受到破坏的植物,因此云南榧树群落也极易受影响。一旦群落遭到破坏,原生草本容易散失,次生草本也容易进入。
群落次生度高达43.80%,说明人为活动的影响使得不少其他生态系统物种进入到云南榧树群落。突出表现为大径级和通直的植株被砍伐,群落郁闭度降低,原来分布在林缘的次生物种大量进入林内,改变了群落原有的区系成分。云南榧树幼苗较少,且都位于石缝间。推测云南榧树种子为某种或某几种动物所喜爱的食物,在每年种子成熟时对其进行搬运、啃食。将野外成熟掉落的种子带回实验室经激素处理后置于培养箱萌发,未见萌发,10个月后种子霉烂。说明种子萌发困难是云南榧树幼苗少的原因之一。另外,人为影响使得生境长期退化,也会导致幼苗减少。云南榧树幼苗减少,种群更新存在困难,种群属于衰退型种群,最终导致云南榧树极其濒危。
综上,人为活动的强烈干扰,如放牧、采集、偷伐等严重,导致次生物种大量进入,原生物种逐渐消失,次生度增大,使得适合云南榧树生存的生境逐渐减少,幼苗变少,种群更新困难,物种面临灭绝的挑战。
致谢:维西县林业局及各乡镇林业站的领导和工作人员在外业调查中给予了极大的支持,同窗研究生在野外调查、标本鉴定中共同工作,在此表示感谢!
[1]王艇,苏应娟,黄超,等.红豆杉科植物RAPD分析及其系统学意义[J].西北植物学报,2000,20(2):243-249.
[2]周其兴.红豆杉科及其相关类群的分支分析[J].大连大学学报,2001,22(6):30-36.
[3]汪松,解焱.中国物种红色名录[M].北京:高等教育出版社,2004:308.
[4]马长乐,胥辉,白琼,等.ICP-OES法测定云南榧树不同部位矿质元素[J].东北林业大学学报,2012,40(7):30-36.
[5]马长乐,黄晓霞,艾星梅,等.云南榧内生菌的分离及抗植物病原菌活性筛选[J].西部林业科学,2014,43(4):10-14.
[6]陈振德,谢立,何英,等.云南榧子中多糖的含量测定及其高效毛细管电泳指纹图谱分析[J].中国药房,2005,16(23):1817-1818.
[7]云南省气象局.云南气候图册[M].昆明:云南人民出版社,1982.
[8]中国科学院中国植物志编辑委员会,中国植物志[M].北京:科学出版社,1991.
[9]中国科学院昆明植物研究所.云南植物志[M].北京:科学出版社,2003.
[10]王荷生.植物区系地理[M].北京:科学出版社,1992:9-17.
[11]宋永昌.植被生态学[M].上海:华东师范大学出版社,2001:99-116.
[12]吴征镒,周浙昆,李德铢,等.世界种子植物科的分布区类型系统[J].云南植物研究,2003,25(3):245-257.
[13]吴征镒.中国种子植物属的分布区类型[J].云南植物研究,1991(增刊,Ⅳ):1-139.
[14]杜凡,杨宇明,王娟,等.区系次生度和生态次生度在生物多样性评价中的应用研究[J].西部林业科学,2012,41(1):41-45.
[15]王娟,马钦彦,杜凡.云南大围山国家级自然保护区种子植物区系多样性特征[J].林业科学,2006,41(1):7-15.
[16]应俊生,张玉龙.中国种子植物特有属[M].北京:中国科学出版社,1994.
[17]周伟,陈宝昆.云南碧塔海自然保护区[M].昆明:云南科技出版社,2010:36-56.
[18]李锡文.中国特有种子植物属在云南的两大生物多样性中心及其特征[J].云南植物研究,1994,16(3):221-227.
