Pit-1基因的结构及生物信息学分析
2015-01-08王玺年陈金萍
王玺年,陈金萍
(1.甘肃畜牧工程职业技术学院,甘肃 武威733006;2.天祝藏族自治县松山镇畜牧兽医站)
垂体特异转录因子1(pituitary specific transcription factor 1)是POU 结构域中同源异性蛋白之一,是动物垂体前叶特异表达的一种具有重要功能的转录因子,具有高度保守的DNA 结合结构域。Woollard等利用原位杂交将山羊Pit-1基因定位于1q21-22染色体区域上,主要功能是正向调控促甲状腺β 亚单位(Thyroid-stimulating Hormoneβ,TSH-β)、促 乳 素(Prolactin,PRL)和 生 长 激 素(Growth Hormone,GH)基因的转录,从而对动物的生长、发育及免疫等具有重要调控作用,能够提高动物生长速度、降低脂肪沉积等。Pit-1基因的突变可以导致垂体的发育受阻,并阻碍GH、PRL、TSHβ基因的正常表达,从而使个体因多种垂体激素的缺乏而导致矮小现象(dwarfism)的出现。因此,Pit-1基因在垂体细胞的发育上扮演重要角色。本研究利用生物信息学方法对山羊Pit-1基因及其编码蛋白的理化性质、蛋白质结构以及生物学功能进行预测和分析,与人、牛和猪Pit-1蛋白基本性质与结构功能进行比较。通过本研究有助于从分子水平上理解Pit-1蛋白基本结构和生物学功能,为进一步研究Pit-1基因在种内及种间遗传分化和该蛋白的功能特性提供理论依据。
1 材料与方法
1.1 生物信息学数据库和软件
美国国立生物技术研究中心:http://www.ncbi.nlm.nih.gov;瑞士生物研究所提供的蛋白数据库:http://www.expasy.ch/tools;丹麦技术大学生物学 序 列 分 析 中 心:http://www.cbs.dtu.dk/services/;本地软件Antheprot、NDAStar、BioEdit和MEGA 等。
1.2 材料
不同物种Pit-1蛋白序列和核苷酸序列均来自NCBI,包括蛋白序列绵羊(P79364)、牛(P10036)、人(P28069)、猪(Q04788)、猕猴(Q28503)、褐 鼠(P10037)、小家鼠(Q00286)、家犬(Q9TT17)、鸡(Q9YGL7);核苷酸序列绵羊(NM_001009350)、牛(NM _174579.3)、猪(HM163575)、人(NM _000306)。
2 结果与分析
2.1 Pit-1蛋白功能分类及二级结构预测
2.1.1 Pit-1蛋白功能分类预测与分析 利用Protfun分析软件预测山羊Pit-1编码蛋白的功能。从表1可知,该蛋白具有转录调控、生长因子和转录功能的可能性分别为2.371、0.498 和1.624,作为受体的可能性仅0.035。Pit-1是编码垂体前叶特异表达的一种转录因子,在垂体细胞中对生长激素细胞的分泌及其基因转录调控过程中发挥重要作用,可能对山羊早期的生长发育起到重要的调控作用。
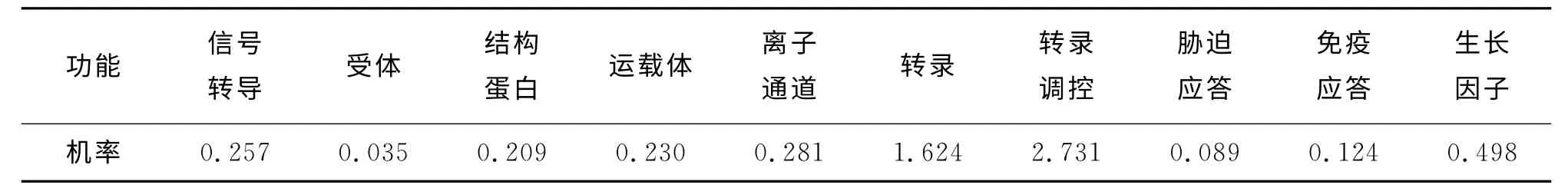
表1 山羊Pit-1编码蛋白功能分析结果
2.1.2 Pit-1蛋白的二级结构预测 采用GOR 法预测山羊Pit-1 编码蛋白的二级结构(图1),山羊Pit-1编码蛋白是由44.8%α-螺旋(Helix)、5.1%β-折叠(Sheet)、16.1% 转角(Turn)和34.0%无规则卷曲(Coil)组成。由此可推测,α-螺旋和无规则卷曲是山羊Pit-1最主要的蛋白质二级结构元件。通过物种间比较发现(表2),山羊Pit-1蛋白中α-螺旋比人和猪少3.4%、2.7%,转角比人和猪多1.8%、0.9%,无规则卷曲比人和猪多1.9%、2.8%。说明山羊Pit-1蛋白在空间结构上与猪和人的相比发生了一定变异。
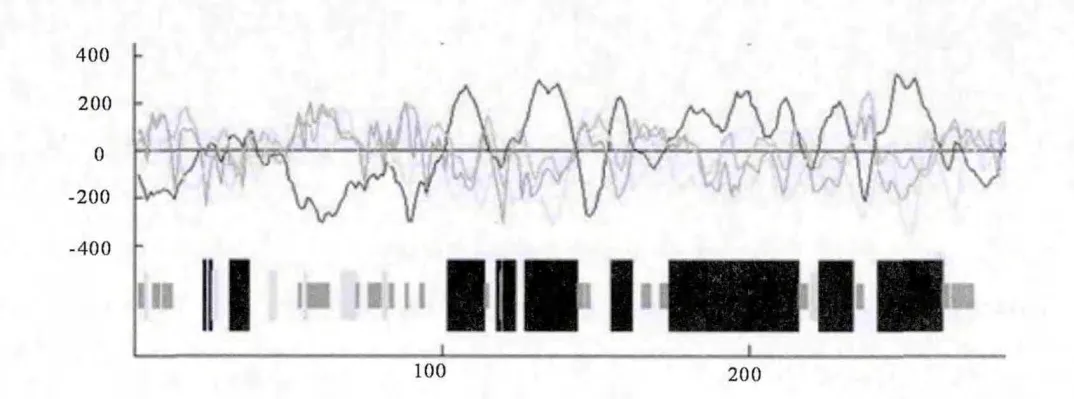
图1 山羊Pit-1蛋白二级结构预测
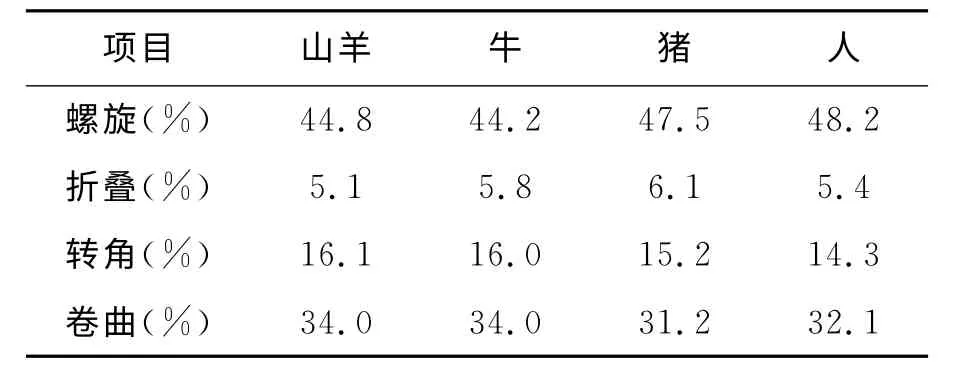
表2 山羊与其他物种Pit-1蛋白二级结构的比较
2.2 Pit-1编码蛋白的基本性质分析
通过ORF finder软件分析发现,山羊Pit-1编码蛋白长度为291个氨基酸残基,其理论分子式为C1446H2299N411O439S15,分子量约为32 946.5D,理论等电点为8.36。统计山羊Pit-1分子氨基酸组成发现,亮氨酸(Leu)、谷氨酸(Glu)含量最为丰富,色氨酸(Trp)含量最低。总的带负电荷残基总数(Asp+Glu)为35个,带正电荷残基总数(Arg+Lys)为38个。比较不同物种氨基酸组成,发现各物种除了在丙氨酸(Ala)、脯氨酸(Pro)、丝氨酸(Ser)和苏氨酸(Thr)的比例存在差异外,其余氨基酸组成基本一致,且都属于不稳定蛋白质(图2)。
2.3 Pit-1编码蛋白跨膜结构分析
跨膜蛋白的预测基于统计学模型和神经网络,跨膜结构域是膜内蛋白与膜脂结合的主要部位,一般有20个左右的疏水氨基酸组成。跨膜结构的预测,对正确认识蛋白质的功能及其在细胞中的作用有重要意义。
本研究通过分析发现(图3),山羊Pit-1蛋白的N 端可能在外侧,并且在第63~79个氨基酸之间可能存在一个由外向内的横跨膜拓扑结构。

图2 不同物种Pit-1基因CDS序列的氨基酸组成

图3 山羊Pit-1蛋白跨膜区分析
2.4 Pit-1编码蛋白磷酸化位点分析
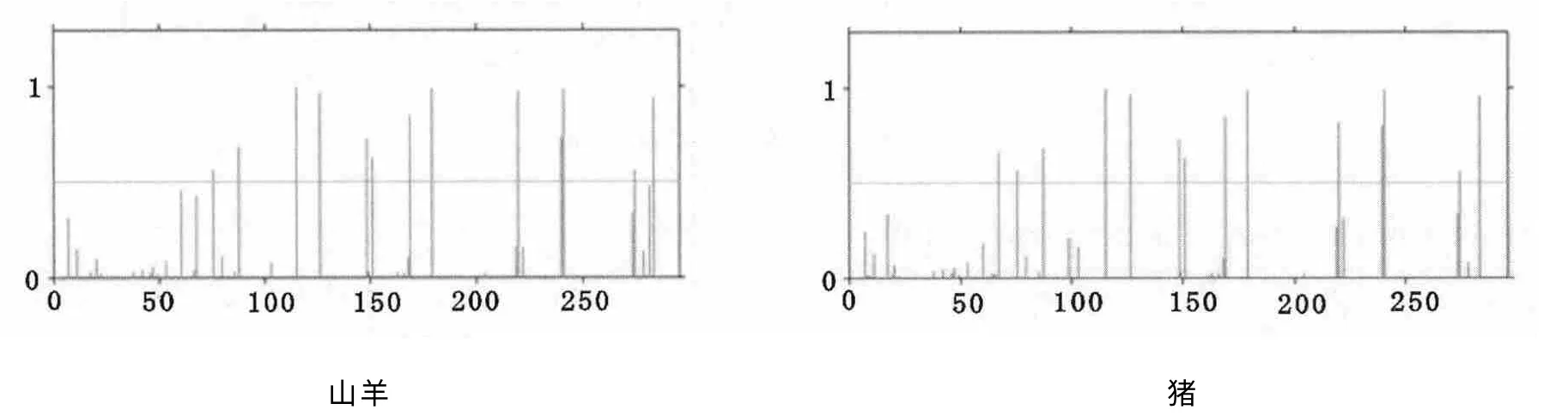
图4 Pit-1蛋白磷酸化作用位点分析
利用神经网络预测法(NetPhos)对Pit-1编码蛋白进行磷酸化位点预测,发现山羊Pit-1 编码蛋白有8个Ser、4个Thr和2个Tyr可能是磷酸化位点。与牛、猪和人对比发现,人与猪Pit-1蛋白缺少一个位于多肽链67位的Tyr磷酸化位点(图4)。
2.5 Pit-1编码蛋白的卷曲螺旋分析
卷曲螺旋主要存在于一些转录因子、结构蛋白以及酶中,在细胞与外界环境之间的物质与信息交换中可以发挥重要功能。Pit-1作为一种转录因子,对GH、PRL 和TSH 基因的转录表达起到正向调控作用。因此,有必要对其卷曲螺旋结构进行信息学分析。利用在线工具COILS预测Pit-1蛋白卷曲螺旋结构(图5)。山羊第110、135、195、250和275个氨基酸附近存在卷曲螺旋可能性很大,可能对正向调控促乳素和生长激素基因的转录表达具有一定的作用。与人、牛和猪对比发现猪缺少一个位于多肽链135位氨基酸附近的卷曲螺旋结构,但含有此卷曲螺旋结构的概率很低,仅为0.089,推测Pit-1蛋白在垂体细胞中发挥其转录调控功能在物种间具有相似性。
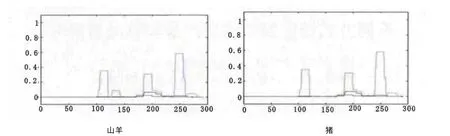
图5 Pit-1蛋白卷曲螺旋预测
2.6 Pit-1编码蛋白亚细胞定位分析
利用TargetP1.1 和PSORTⅡ软件分析山羊Pit-1蛋白的亚细胞定位,结果表明(表3)该蛋白最终定位于细胞核的可能性为70%,定位于微体细胞的可能性为30%,定位溶酶体的可能性为10%。预测山羊Pit-1蛋白最终定位于细胞核内,在细胞核内发挥生物学作用,即这种蛋白为核靶向蛋白。

表3 山羊Pit-1蛋白质亚细胞定位结果
3 讨论
通过对山羊Pit-1 基因的生物信息学分析发现,山羊Pit-1蛋白的分子质量为32 946.5D,整条多肽链表现为亲水性,其二级结构以α-螺旋和无规则卷曲为主要的结构元件,该蛋白最终定位于细胞核的可能性为70%,可能在细胞核中发挥生物学作用。山羊Pit-1编码蛋白含有5个卷曲螺旋结构,14个磷酸化位点,这些位点的出现可能有利于蛋白质信号的传导,来诱导神经系统对外界刺激的调控,如调节生长激素基因、促乳素基因的转录表达。并且在第35个氨基酸附近有一个可能的剪切位置,在第63~79个氨基酸区域可能存在一个由外向内的横跨膜拓扑结构,这些剪切位点和跨膜结构可能会导致编码序列氨基酸结构的改变进而影响该基因的结构和功能分类。
Pit-1基因在垂体发育和激素表达中起关键作用,对调节细胞分泌和细胞发育起主导作用,能够识别特异的基因顺序,并与之结合,从而引起细胞内基因的转录,促使垂体前叶生长激素、催乳素、促甲状腺素β细胞及其基因的转录和表达,从而影响动物生长发育。近年来关于Pit-1 基因的研究较多,对人类和小鼠的研究表明,Pit-1基因突变与小鼠矮小基因座(dwarf locus)和人类综合性垂体激素缺陷症(CPHD)相关;在猪上其多态性与胴体组成性状、生长性状有关联;在牛上该基因多态位点与牛的生长和胴体性状存在关联,是标记早期生长性状最理想的候选基因。Song等初步证实,Pit-1 基因的遗传突变及其与家畜生产性能的相关性可作为MAS(marker assistant selection)的 有效候 选基因。本研究利用多种生物信息学软件对山羊Pit-1基因编码蛋白质的理化性质、结构和功能进行初步探索,并同人、牛和猪的Pit-1 蛋白进行了比对,发现山羊Pit-1蛋白的20种氨基酸组成比例、基本性质及编码蛋白的功能与牛、猪Pit-1蛋白具有相似的功能,为进一步对该基因的功能研究提供理论依据提示Pit-1 基因可能是山羊生长性状的数量性状位点(quantitative trait loci,QTL)或者其与生长性状的QTL存在紧密连锁。
[1] Rosenfeld M G.POU-domain transcription factors:powerful developmental regulators[J].Genes Development,1991,5:897-907.
[2] Woollard J,Tuggle C K,Ponce de Leon F A.Rapid communication:Localization of POU1F1to bovine,ovine and caprine[J].Anim Sci,2000,78:242-243.
[3] 滕勇,经荣斌,宋成义.哺乳动物POU1F1基因对其生长发育 的作用[J].西 北 农 业学报,2004,13(2):11-15.
[4] Primus E,Mullis M D.Genetics of growth hormone deficiency[J].Endocrinol Metab Clin N Am,2007,36(1):17-36.
[5] Li S,Crenshaw E B,Rawson E J,et al.Dwarf locus mutants lacking three pituitary cell types result from mutations in the POU-domain gene Pit-1[J].Nature,1990,347:528-533.
[6] 宋成义,赵芹,高波.猪POU1F1基因启动子区多态分析及其与生长形状的关联[J].中国农业科学,2011,44(24):5067-5072.
[7] Zhao Q,Davis M E,Hines H C.Associations of polymorphisms in the Pit-1gene with growth and carcass traits in Angus beef cattle[J].Journal of Animal Science,2004,82(8):2229-2233.
[8] 刘波,陈宏,蓝贤勇.秦川牛及其杂种牛POU1F1 基因多态与生长性能相关性[J].中国农业科学,2005,38(12):2520-2525.
[9] Song C Y,Teng Y,Gao B,et al.MspⅠpolymorphisms in the 3rd intron of the swine POU1F1(Pit-1)gene and its correlation with growth performance[J].Animal Biotechnology Bulletin,2004,9(1):38-45.
[10] Xue Kai,Chen Hong,Wang Shan,et al.Effect of Genetic Variations of the POU1F1 Gene on Growth Traits of Nanyang Cattle[J].遗 传 学 报,2006,33(10):901-907.
[11] 罗卫星,蔡惠芬,王兴群,等.山羊垂体转录因子POU1F1基因多态性及其与屠宰性状相关性研究[J].中国畜牧杂志,2011,47(9):5-9.
