千岛湖马尾松群落种群生态位与种间联结
2015-01-04王秀云朱汤军徐高福
王秀云,朱汤军,徐高福
(1. 浙江省林业科学研究院,浙江 杭州 310023;2. 新安江开发总公司,浙江 淳安 311700)
千岛湖马尾松群落种群生态位与种间联结
王秀云1,朱汤军1,徐高福2
(1. 浙江省林业科学研究院,浙江 杭州 310023;2. 新安江开发总公司,浙江 淳安 311700)
在对千岛湖马尾松(Pinus massoniana)群落野外调查基础上,采用生态位宽度、生态位重叠和基于 2×2联列表的x2检验、联结系数及共同出现百分率的方法,分析了群落主要种群间的相互关系。结果表明:群落中马尾松的生态位宽度最大,杉木的生态位宽度最小,生态位较大的物种间生态位重叠值也较高,马尾松与黄檀之间的生态位重叠值要明显高于其他种群的,Schoener指数为0.750 3。马尾松与其他种群之间的联结性较显著,而且多数为正联结。种间正联结性越强,其对环境要求越接近,生态位重叠值越大。结合联结系数及共同出现百分率指标对其种间联结性的程度测定显示:各项指标对马尾松群落主要种群之间联结性关系总体上反映了大致相同的结果,但不同的测定方法可以相互弥补由于单一方法分析的不足。
马尾松;生态位;种间联结;千岛湖
生态位与种间联结是种群生态研究的核心问题。生态位能够反映种群在群落中的时间、空间及资源利用方面的竞争关系[1~2],是物种对环境的影响、环境对物种的影响两方面的相互作用[3]。种间联结是指不同物种在空间分布上的相互联结性,通常是由于群落生境的差异影响了物种分布而引起的,是对各个物种在不同生境中相互影响、相互作用所形成的有机联系的反映,它表示种间相互吸引或排斥的性质[4]。生态位与种间联结是植物群落重要的数量和结构特征,它们作为2个物种相似性的一种尺度,对于正确认识群落的结构、功能和分类有着重要的指导意义,并能为植被的经营管理、自然植被恢复和生物多样性保护提供理论依据。近年来,对种群生态位与种间联结研究日益受到重视,如李帅锋[5]、张志勇[6]、李先琨[7]、林长松[8]等,这些研究为种群的生物多样性保护及资源可持续利用提供参考。
以马尾松(Pinus massoniana)为主的天然次生林,在千岛湖库区生态环境保护中发挥着极其重要的作用,但由于近年来人为干扰,部分马尾松群落出现了退化[9]。为此,本研究以浙江省淳安县千岛湖林区的马尾松林为研究对象,通过对马尾松群落主要种群生态位与种间关系进行研究,以期揭示主要种群在群落中的功能地位、种群间的相互作用、生态适应性及各种群对资源的利用状况,从而为马尾松的保护与资源合理利用提供科学依据。
1 研究区概况
研究区位于浙江省淳安县千岛湖林区,东邻建德县,西与安徽省歙县毗连,118° 34′ ~ 119° 15′ E,29°22΄ ~ 29° 50′ N。气候属中亚热带季风气候北缘,温暖温润,雨量充沛,四季分明,光照充足,光、温、水的地域差异明显。年平均气温17℃,≥10℃积温5 410℃,年日照时数1 951 h,无霜期约263 d,年均降水量1 430 mm,相对湿度为76%。土壤主要类型有红壤,分布于海拔700 m以下,pH值5.5 ~ 6.0;黄壤分布于海拔700 m以上,pH值5.5左右;岩性土分布于海拔700 m以下,pH值7.0以上。研究区种子植物资源135科,398属,634种(含种以下分类等级)。千岛湖森林生态系统主要为暖性针叶林、常绿阔叶林、落叶阔叶林、常绿落叶阔叶混交林、针阔混交林、竹林、灌丛、灌草丛等类型。植物种类丰富,有乔木125种,灌木45种,藤本31种,草本植物133种。其代表的乔木种有马尾松(Pinus massoniana)、杉木(Cunninghamia lanceolata)、柏木(Cupressus funebris)、苦槠(Castanopsis sclerophylla)、青冈(Cyclobalanopsis glauca)、柯(Lithocarpus glaber)、枫香(Liquidambar formosana)、木荷(Schima superba)等;灌木种格药柃(Eurya muricata)、檵木(Loropetalum chinense)、短尾越桔(Vaccinium carlesii)、江南越桔(V. mandarinorum)、马银花(Rhododendron ovatum)等,草本种芒萁(Dicranopteris dichotoma)、蕨(Pteridium aquilinum var. latiusculum)等,藤本植物菝葜(Smilax china)、紫藤(Wisteria sinensis)等。
2 研究方法
2.1 样地设置
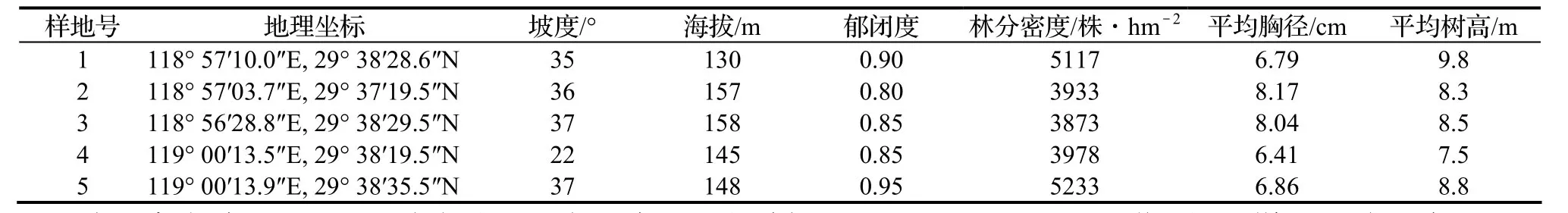
表1 各样地基本情况Table 1 General conditions of sampling plots
在千岛湖林区以马尾松为主的天然次生林中,分别在阴坡、阳坡不同立地中设置调查样地5个(表1),样地面积30 m×30 m,将样地划分为10 m×10 m的小样方,逐个样方调查所有胸径(DBH)≥3 cm的植物,记录乔木、灌木物种的名称、高度、胸径、冠幅(盖度)。同时记录各样地的海拔、坡度、坡向等立地因子。全部野外工作于2012年8-9月完成。
由于植物种类较多,将植物按照在样地中出现的频率由大到小排序,筛选出现频率较高的植物种类进行生态位与种间联结性的计算和分析,参加分析的物种有14种(乔木9种,灌木5种)。
2.2 生态位测定方法
本研究采用Levins指数测定种群的生态位宽度,采用Schoener指数与Pianka指数测度优势种群间的生态位重叠[10~11]。
Levins生态位宽度(Bi):
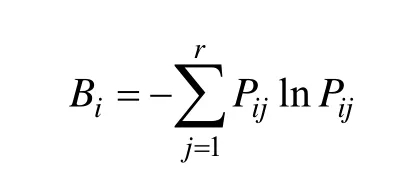
Schoener指数(Cih):
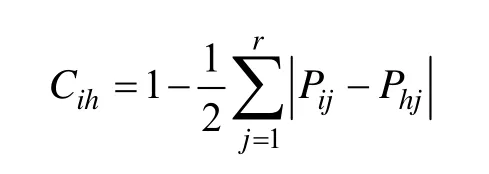
Pianka重叠指数(Np):
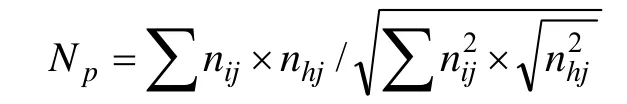
式中,Bi是种i的生态位宽度,Pij是种i对第j个资源的利用占它对全部资源利用的频率,即Pij= nij/Ni,而,nij为种i在资源j上的优势度(本文即物种的重要值),r为资源等级数;Cih为Schoener指数,表示物种i与物种h的相似程度,且有Cih= Chi;Np为Pianka重叠指数,nij和nhj为种i和种h在资源j上的优势度,本文即物种的重要值比例,重要值(IV)=(相对密度+相对显著度+相对频度)×100/3[12],Schoener指数与Pianka重叠指数的值均介于0和1之间。
2.3 种间联结测定方法
以x2统计量为基础,以联结系数与共同出现的百分率确定物种间联结性。
2.3.1 x2统计量的计算与检验 由于取样为非连续性取样,原始数据为时间存在与否的二元数据,因此非连续性数据的x2值用Yates的连续性校正公式计算[13]。
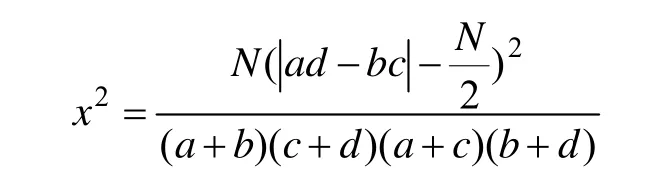
式中:a是所调查的所有样方中,物种A与物种B同时出现的样方数;b为物种A出现,物种B不出现的样方数,c为物种A不出现,而物种B出现的样方数,d为物种A与B都不出现的样方数,下同。
种间联结强弱由x2决定,若x2≥3.841 (0.01<p<0.05),表示种间联结显著;x2≥6.635(p<0.01),表示种间联结极显著;x2<3.841(p>0.05)时,认为两个种独立分布,即中性联结。当ad>bc时为正联结,ad<bc时为负联结。
2.3.2 联结系数(Y) 联结系数用来进一步检验由x2所测出的结果及说明种间联结程度,其计算公式[14]为:
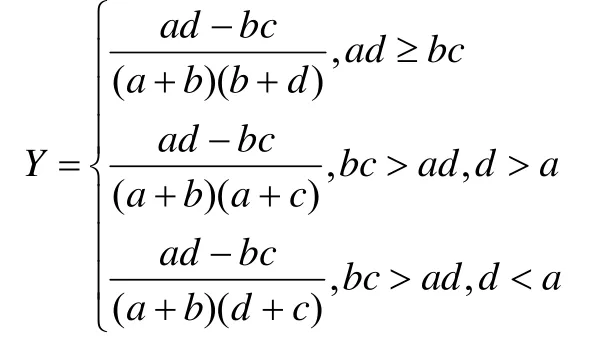
Y值域为[-1, 1],Y越接近1物种间正关联越强,反之,越接近-1物种间负关联越强,Y为0表明物种间完全独立。
2.3.3 共同出现的百分率(P) 用来测度物种间正联结强度,其计算公式[14]为:
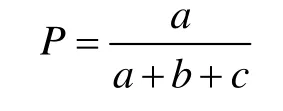
P的大小在0和1之间变动,其值越接近1,关联程度越大,表明该种对的正联结越紧密,无关联时为0。
3 结果与分析
3.1 生态位宽度
生态位宽度是反映一个种群所利用的各种不同资源的总体能力,是度量种群对环境的适应能力或对资源利用程度的尺度。生态位宽度越大,物种对环境的适应能力越强,在群落中的分布幅度相应地越大。从表2可看出,马尾松在群落中居绝对优势,表明马尾松在本区环境资源利用最为广泛,利用资源的能力强。其结果与常绿阔叶林遭破坏后形成的次生植被,以马尾松首先侵入生长相符合。地带性落叶阔叶树种枫香、黄檀,其Bi值分别为0.558 5与 0.491 4。说明在马尾松群落中,地带性落叶阔叶树种的生态位幅度较广,并与马尾松在乔木层构成亚层,一般马尾松位于主林层,地带性阔叶树种居第二层,形成复层结构的演替过程。而且地带性乡土树种如柯、木荷、短柄枹栎其幼苗幼树耐荫且更新能力强,因此在马尾松林下能更新生长,易与马尾松形成混交林,相对适应环境能力较好。灌木树种南烛、格药柃、短尾越桔、檵木的生态位宽度Bi值分别为0.561 8、0.522 8、0.485 7和0.464 4,其适应环境能力较强,特别适合于马尾松林下生长。而且在所调查的样地中零星分布着山槐、杉木等乔灌木树种,由于研究区立地条件差,不适宜于山槐、杉木的生长,所以仅占有一个资源位,其生态位宽度最低。
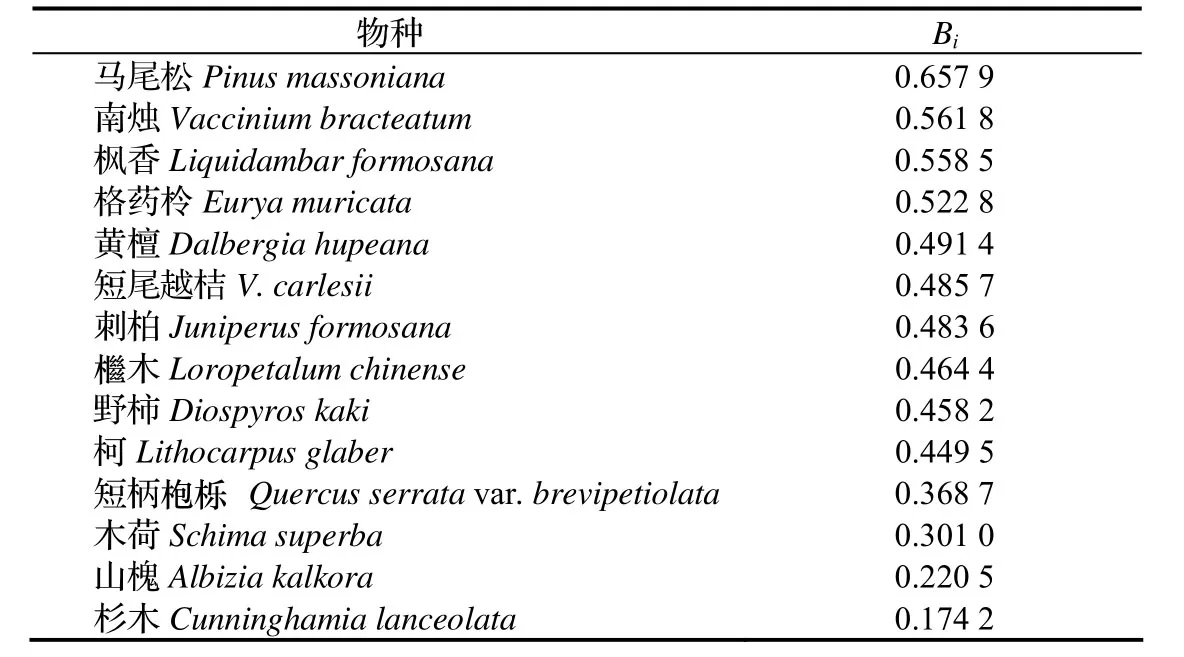
表2 马尾松群落优势种群生态位宽度Table 2 Niche breadth of dominant populations in P. massoniana communities
此时期是先锋树种生态位压缩和后期竞争种生态位扩展的过渡阶段,先锋树种马尾松与落叶阔叶树种黄檀、枫香共同控制群落的结构和功能。
3.2 生态位重叠
当两个物种利用同一资源或共同占有某一资源因素(如营养成分、空间等)时就会出现生态位重叠现象,生态位重叠较大的物种常常具有相近的生态习性或有互补性要求。Schoener指数与Pianaka指数两个生态位重叠值为0的种对占总种对数的2.20%(表3)。生态位宽度较大的物种与其它种群间的生态位重叠较大,群落中Schoener指数大于0.8的种对有4个,如南烛—檵木;但也并不意味着生态位宽度较小的物种之生态位重叠就一定小,群落中Pianaka指数大于0.8的有3个,分别为木荷—柯、木荷—杉木及石柯—杉木。马尾松与其他物种的Schoener指数在0.257 0 ~ 0.761 5,Pianaka指数在0.285 9 ~ 0.709 9。
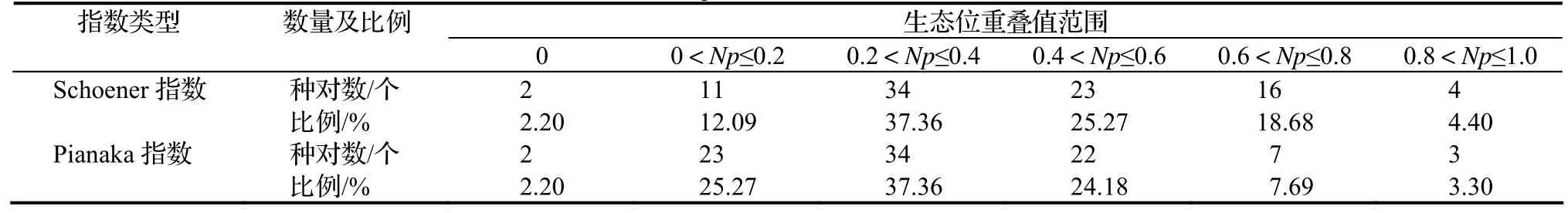
表3 马尾松群落生态位重叠值Table 3 Niche overlap values in P. massoniana communities
3.3 种间联结分析
千岛湖马尾松林主要种群间x2统计量数阵、联系系数半矩阵图以及共同出现的百分率半矩阵图分别见表4、表5和图1、图2。
3.3.1 x2统计量检验 经x2统计量矩阵分析(表4),马尾松林91个种对中,正联结的种对数为48个,占总种对数的52.75%,其中极显著正关联(x2≥6.635)、显著正关联(3.841≤x2<6.635)与不显著正关联(x2<3.841)的种对数分别为3个、3个及42个。负联结的种对数为37个,占总种对数的40.66%,其中极显著负关联、显著负关联与不显著负关联的种对数分别为1个、3个及33个。种对2(南烛)—5(黄檀)、3(枫香)—11(短柄枹栎)、4(格药柃)—7(刺柏)、6(短尾越桔)—10(柯)、9(野柿)—13(山槐)及13—14(杉木)之间无联结(x2= 0),因为这些种在群落中通常只在少数的样方或局部的样方中出现,因而表现出较强的独立性。
马尾松除与枫香、南烛、柯、木荷及短柄枹栎为不显著负关联外,与其他大部分种群均为正联结,其中与杉木在0.05的水平上呈现显著的正关联(x2= 6.33),说明它们之间对生境具有相同或相似的需求与适应并相互依存。千岛湖马尾松群落总体呈现正联结,说明马尾松群落稳定性基础仍存在。
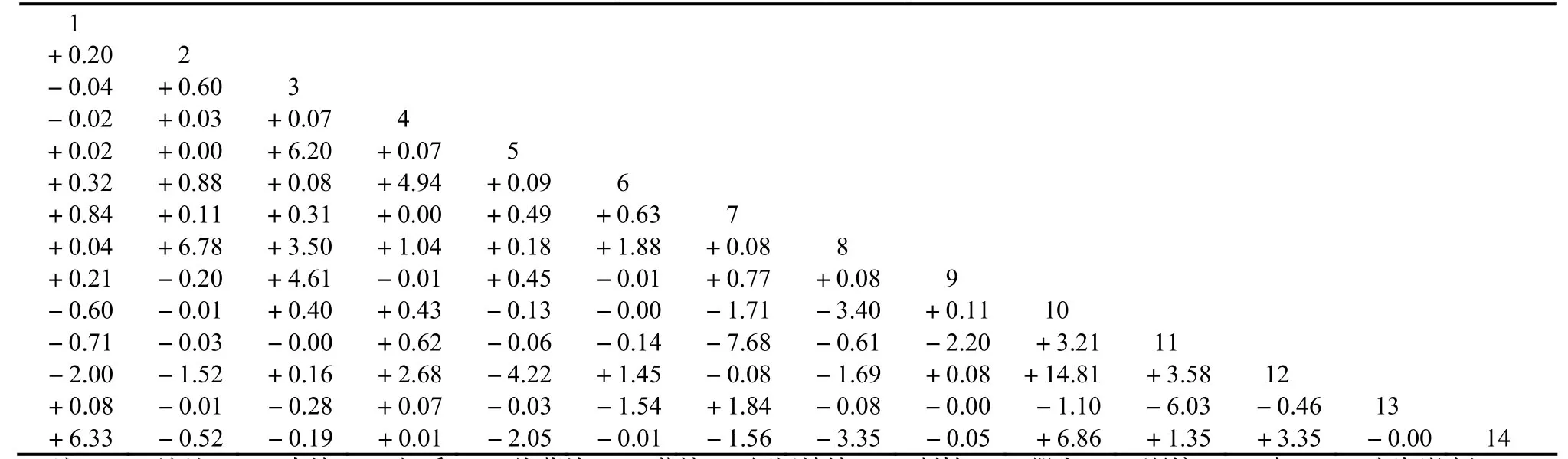
表4 千岛湖马尾松林种群间x2统计量矩阵Table 4 Matrix analysis on interspecific association by Chi-square test in P. massoniana communities
注:1—马尾松,2—南烛,3—枫香,4—格药柃,5—黄檀6—短尾越桔,7—刺柏,8—檵木,9—野柿,10—柯,11—短柄枹栎,12—木荷,13—山槐,14—杉木,下同。
3.3.2 共同出现的百分率和联结系数 虽然x2统计量值比较客观准确地判断种对联结的显著性,但是,共同出现的百分率和联结系数却能体现出x2检验不显著种对的联结性及其大小。所以在种间联结性研究中都是以x2检验为基础,结合共同出现百分率和联结系数来检验植物群落种间联结性,可以得出更为准确的结果。马尾松群落91个种对的共同出现百分率和联结系数测定和分析统计结果见表5、图1和图2。
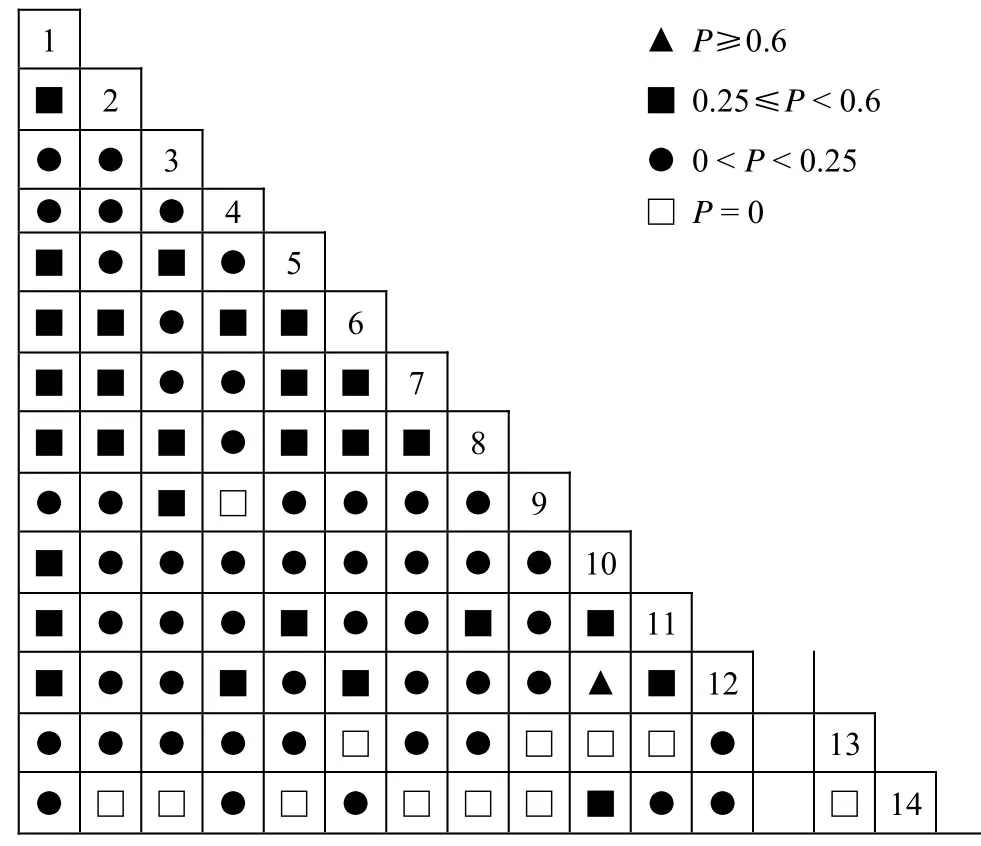
图1 千岛湖马尾松群落种群间共同出现百分率(P)关矩阵Figure 1 Semimartrices diagrams for percentage co-occurrence(P) of dominant populations in P. massoniana communities

图2 千岛湖马尾松群落种间联结系数(Y)关矩阵Figure 2 Semimartrices diagrams for association coefficient(Y) of dominant populations in P. massoniana communities

表5 马尾松林主要种群共同出现百分率(P)和联结系数(Y)检验与x2检测比较分析Table 5 Analysis of association co efficient(Y) and percentage co-occurrence (P) with x2-test of domina nt populations in P. massoniana communities
由图1及表5可见,马尾松群落主要物种91个种对间的共同出现百分率检测结果,P≥0.25的种对有29个,所占比例为31.87%,其中种对10(柯)—12(木荷)的P值为0.6,关联程度最高;经P值测定,相互间无关联的种对有12个(P = 0),即这12个种对在45个样方中从未同时出现,所占比例为13.19%;关联度介于0和0.25之间的种对有50个,所占比例为54.94%。与马尾松种群联结程度较高的种群有檵木、短柄枹栎、黄檀、刺柏等。
从图2和表5可见,Y≥0.3的显著关联的种对有26个,占总种对数的28.57%,比x2检测的显著关联种对数多,尤其是显著负关联的种对较为明显,其中正关联的只有3对,负关联的23个种对。Y≤-0.6的种对多达16个,其中有12个种对P= 0,显示种对相互独立。可见通过Y值测定,可以更准确地检测出那些关联度较高但经x2和P检测为关联不显著的种对,但另一方面也可能夸大了种间的负联结性程度;经Y值测定,相互间无关联的种对有2个(Y = 0),分别为种对1(马尾松)—6(短尾越桔)、5(黄檀)—6(短尾越桔)。
4 结论与讨论
生态位宽度是衡量植物种群对资源环境利用状况的尺度[3],一般来说,生态位宽度越大表明物种对环境的适应能力越强,对资源的利用越充分[15],同时也表明该物种的特化程度越小,倾向于泛化种,而且这些物种在群落中往往处于优势地位,反之,则为竞争力明显较弱的特化种[16]。马尾松群落是亚热带地区的重要植被类型,马尾松是次生演替的先锋树种,数量多,分布广,对群落内环境起着决定性作用,对资源的利用程度最高,生态位宽度最大。枫香、黄檀、刺柏等种群的数量和分布相对于马尾松要小,对资源的利用能力和对群落环境的决定作用都次于马尾松,因此生态位宽度小于马尾松种群;但这些种群是马尾松林中重要的优势种,是群落的重要伴生物种,为群落中的泛化种,生态幅较广,在创造群落环境中起到重要的作用,所以它们的生态位宽度也较大。马尾松群落中光照较足,喜阳但耐荫树种如木荷、柯、短柄枹栎长势良好,对资源利用比较充分,生态位宽度较高。灌木种南烛、格药柃、檵木及短尾越桔在群落中分布范围较广,多出现在马尾松林下,所以生态位宽度较高。而山槐和杉木种群数量上不是很多,分布范围较小,且分布不均匀,在群落中的地位较低,对环境资源的利用能力较弱,生态位宽度很小。
生态位重叠是群落物种多样性与群落结构的决定因素之一,生态位宽度能较好地反映出植物种群的竞争重叠状况[17]。群落中大多数种群间的生态位重叠较大,Schoener指数高于0.4的种对有43个,占总种对数的47.25%,Pianaka指数大于0.4的种对有32个,占全部种对的35.16%,说明群落内种群间对环境资源具有相似的生态需求,所产生的资源利用性竞争激烈,如马尾松—黄檀、黄檀—枫香等种对;Pianaka指数显示出生态位宽度较小的物种之间也可能有较大的生态位重叠值,这与生态位较小的种群对环境的适应力和资源需求存在较大差别及立地条件有关[18],本研究群落内部有较多的岩石裸露,坡度较大,地形较为复杂,造成物种分布的斑块性与环境资源较高的空间异质性,常常使物种有较高的聚集度,导致有较小生态位宽度的物种出现生态位重叠较高的现象,如木荷—柯、木荷—杉木。群落中其他物种与马尾松的生态位重叠值最大,其次是南烛,前者由于是群落乔木层的优势树种,后者则是灌木层的优势物种,生态位宽度与物种间生态位重叠具有密切关系,生态位宽度较大的树种之间生态位重叠机会也较大。
群落中正值的联结系数反映出两个物种对环境差异有相似的反应,负值则反映物种间对所需环境条件不同或是一个种存在对另一个种不利而产生排斥。千岛湖马尾松群落主要种群总体上表现为正联结,可以认为千岛湖马尾松群落稳定的基础依然存在,但在总的种对数中极显著和显著正联结的种对数较少,这说明马尾松群落稳定性较差,仍然处于不断的发展演替过程之中。在森林间伐、择伐等抚育过程中,依据森林植物群落种间联结性特点,凡与马尾松有显著正关联的树种应当作为经营目标树加以保留;相反,负关联的树种可以伐除。在对地带性植被进行改造或经营时,要充分考虑植物群落种间联结性,有目标地选择性进行间伐作业。
在进行种间联结性研究时,不宜根据单一指标作出结论,最好多个指标配合使用,以说明种间联结程度。经比较共同共同出现的百分率、联结系数测定与x2检验结果,对马尾松群落主要种群间联结性关系总体上反映了大致相同的结果,但不同的测定方法针对性有所差别。x2统计量由于有明确的指标,能比较准确客观地表现种间联结性,但x2检验只是对种联结性给出了一个概括性的结论;共同出现的百分率值和联结系数值虽能反映种间联结性的相对强弱,但对联结性强弱的等级划分缺少统一的标准,并且在物种出现次数较少,特别是当a = 0时,共同出现的百分率值和联结系数值均会夸大物种间的联结性,甚至会得出不同的结论。共同出现的百分率值由于忽视了d值的作用,a值较小时,无形中夸大了a、b、c值在联结性测定中的作用。因此,以x2检验为基础测定物种之间联结性的显著性,结合共同出现百分率和联结系数来测定物种间的关联度,能够得出比较全面和更为准确的结论。
[1] 王祥福,郭泉水,巴哈尔古丽,等. 崖柏群落优势乔木种群生态位[J]. 林业科学,2008,44(4):6-13.
[2] 王伯荪,李明光,彭少麟. 植物种群学[M]. 广州:中山大学出版社,1989.
[3] 李帅锋,刘万德,苏建荣,等. 滇西北云南红豆杉群落物种生态位与种间联结[J]. 植物科学学报,2012,30(6):568-576.
[4] 张志勇,陶德定,李德珠. 五针白松在群落演替过程中的种间联结性分析[J]. 生物多样性,2003,11(2):125-131.
[5] 李先琨,黄玉清,苏宗明. 南方红豆杉群落主要树木种群间联结关系初步研究[J]. 生态学杂志,1999,18(3):10-14.
[6] 林长松,李玉英,左经会,等. 珍稀植物十齿花群落乔木优势种群种间联结性[J]. 生态学杂,2008,27(2):178-184.
[7] 洪利兴,王泳,杜国坚,等. 我国南方马尾松林生态系统的退化特征和改造对策研究[J]. 浙江林业科技,2000,20(3):1-9.
[8] 李德志,石强,臧润国,等. 物种或种群生态位宽度与生态位重叠的计测模型[J]. 林业科学,2006,42(7):95-103.
[9] 胡正华,钱海源,于明坚. 古田山国家级自然保护区甜槠林优势种群生态位[J]. 生态学报,2009,29(7):3 670-3 677.
[10] 汤景明,艾训儒,易咏梅,等. 鄂西南木林子常绿落叶阔叶混交林恢复过程中优势树种生态位动态[J].生态学报,2012,32(20):6 334 -6 342.
[11] 王乃江,张文辉,陆元昌,等. 陕西子午岭森林植物群落种间联结性[J]. 生态学报,2010,30(1):67-78.
[12] 欧祖兰,李先琨,苏宗明,等. 元宝山冷杉群落主要树木种群间联结关系的研究[J]. 生态学杂志,2002,21(1):14-18.
[13] 张金屯. 数量生态学方法[M]. 北京:科学出版社,1995.
[14] 李菁,骆有庆,石娟,等. 阿尔山地区兴安落叶松林下植物种群生态位[J]. 林业科学研究,2011,24(5):651-658.
[15] 李生,陈存及,曹永慧,等.乳源木莲天然林主要种群生态位研究[J].江西农业大学学报,2003,25(3):374-378.
[16] Weider, L J. Niche breadth and life history variation in a hybrid Daphnia complex[J]. Ecology, 1993,74(3):935-943.
[17] Paoli G D, Curran L M, Zak D R. Soil nutrients and beta diversity in the Bornean Dipterocarpaceae: Evidence for niche partitioning by tropical rain forest trees[J]. J Ecol, 2006(94):157-170.
[18] McKane R B, Johnson L C, Shaver G R, et al. Resource-baced niches provide a basis for plant species diversity and dominance in arctic tundra[J]. Nature, 2002(415):68-71.
Niche and Interspecific Association of Species of Pinus massoniana Communities in Qiandao Lake
WANG Xiu-Yun1,ZHU Tang-Jun1,XU Gao-fu2
(1. Zhejiang Forestry Academy, Hangzhou 310023, China; 2. Xin’anjiang Development Corporation of Zhejiang, Chun’an 311700, China)
Field data collection was based on 5 plots (30m×30m) in Pinus massoniana communities distributed in Qiandao lake, Zhejiang province. Methods including niche breadth, niche overlap, x2-test, association coefficient(AC) and percentage co-occurrence (PC) based on 2×2 contingency table were used for analyzing the interaction between the species population. The niche breadth of P. massoniana was the largest, while that of Cunninghamia lanceolata was the smallest. Simultaneously, niche overlap increased with niche breadth. The niche overlap of species pairs of between P. massoniana and Dalbergia hupeana was larger than species pairs between P. massoniana and other populations. The Schoener index was 0.7503. The inter-specific associations between P and other species were comparatively significant, and mostly were of positive. The more intense the positive association, the more similar the environmental demand required, and the larger the value of niche overlap was. The degree of inter-species association based on x2-test, together with the association coefficient and percentage co-occurrence, showed that the indices applied could reflect the approximately same results, and multiple indices could remedy the insufficiency analyzed by a single index.
Pinus massoniana.; Niche; Interspecific Association; Qiandao Lake
S718.5
A
1001-3776(2015)03-0036-07
2014-09-16;
2015-04-21
浙江省科技厅公益项目(2014C32118);浙江省省院合作林业科技计划项目(2012SY11)
王秀云(1978-),女,内蒙古凉城人,博士,副研究员,从事林业碳汇与森林生态研究;*通讯作者。
