不同混播方式下豆禾混播草地群落稳定性的测度与比较
2015-01-02郑伟加娜尔古丽唐高溶朱进忠
郑伟,加娜尔古丽,唐高溶,朱进忠
(1.新疆农业大学草业与环境科学学院,新疆 乌鲁木齐830052;2.新疆维吾尔自治区草地资源与生态重点实验室,新疆 乌鲁木齐830052)
人工草地(tame grassland,artificial grassland,sowing grassland)是人为结合所在地的具体生态条件和一定的经济利用目标,选择适宜草种而建立的特殊人工植物群落,其生态学本质是在人为农作措施维护下的开放性亚稳态生产系统[1],人工草地在建立之初就存在群落稳定性维持的问题。对于人工草地而言,外界干扰(农作措施)要维持其组分,从而维系稳定的生产能力[2]。人工草地的稳定性维持可理解为:在农作措施干扰下,尽量使群落组分、功能保持稳定,不向原生群落演替[2-3]。稳定性高的人工草地群落应具有以下3个特征:1)群落组分的稳定性;2)群落功能的稳定性;3)低的群落入侵性。
豆禾混播草地,以其较高的生产力、较好的适口性和较高的营养价值受到世界许多地区人们的重视[4-5]。豆禾混播牧草在形态上、生长发育上、营养资源利用上、时空生态层位上都存在着相互补充而非彼此竞争的可能[6],这也是群落组分稳定的实现基础。因此,合理的牧草种类组合、混播比例在实现豆禾混播草地稳定性上存在可能[7]。如何科学测度混播草地群落稳定性,面临着许多问题,特别是对群落稳定性的量化研究十分缺乏[8]。对于群落稳定性的量化研究,许多学者通过Godron的稳定性测定方法,对所研究植物群落中所有物种百分数与累积相对频度比值进行计算,其值越接近20/80,群落越稳定[9],但仅能提供群落稳定与否的信息[10]。也有学者通过主成分分析法对群落的稳定因素进行筛选,并结合Godron测定方法来评价群落稳定性[10];或者通过分析各稳定因素的隶属度,综合评价群落的稳定性[11]。有关混播草地群落稳定性的研究主要关注了种间相容性、环境压力和干扰活动这三个因素[1,3,12],如顾梦鹤等[13]和Dong等[14]在高寒地区对混播禾草草地进行施肥,提高了草地组分稳定性与产量稳定性,抑制了杂草的入侵;李莉等[12]在喀斯特地区对豆禾混播草地进行适当地刈割,或者刈割与放牧有规律地交替使用,有利于维持混播草地组分,减少杂草入侵。但上述研究对群落稳定与否[15]、影响群落稳定性因素的定量分析[12]、多种因素的可比性[7]等关注较少。因此,本研究以建植多年的豆科与禾本科混播草地群落为研究对象,从群落组分的稳定性、群落功能的稳定性和群落入侵性3个方面定量测度和对比分析不同混播种类和混播比例的豆禾混播草地稳定性,从群落稳定性的角度认识豆禾混播草地的演变规律和机理,为豆禾混播草地的长期利用探寻理论依据和技术支撑。
1 材料与方法
1.1 试验地概况
试验地点位于新疆维吾尔自治区伊犁哈萨克自治州昭苏盆地腹内的昭苏马场(E 81°03′-81°05′,N 42°38′-43°15′),该地区属温带山区半湿润易旱冷凉气候类型。年均温度2.7℃,≥10℃年积温1416.8℃,无霜期85~100d,年均降水量512mm,降水集中于5-8月,约占全年总降水量的68%,年均蒸发量1261.6mm,是降水量的2.5倍,积雪期158d,积雪厚度20~60cm。试验小区所处地段原为天然草地(山地草甸),2008年开垦为多年生人工草地。植被主要成分有鸭茅(Dactylisglomerata)、无芒雀麦(Bromusinermis)、新疆鹅观草(Roegneria sinkiangensis)、猫尾草(Phleumpratense)、草原糙苏(Phlomispratensis)、草原老鹳草(Geraniumpratense)、大叶橐吾(Ligulariamacrophylla)等。盖度65%~100%,草层高25~85cm,鲜草产量103905kg/hm2。土壤为黑钙土,土壤有机质含量为13.63%~14.89%,全氮、全磷、全钾量分别为9.53,2.16,12.20g/kg,碱解氮、有效磷、有效钾含量分别为459.20,144.43,473.58mg/kg[5]。
1.2 试验材料与设计
试验为完全随机区组设计,a因素为混播种类(a=7),分别设3种牧草混播的2个处理、4种牧草混播的2个处理、5种牧草混播的2个处理和6种豆禾牧草混播;b因素为混播比例(b=3),分别为豆禾比5∶5、4∶6和3∶7;6种牧草每种各设单播作为对照(CK=6);3次重复(N=3),共81个小区。每个小区面积均为3m×4m,行距30cm。紫花苜蓿(Medicagosativa),红三叶(Trifoliumpratense)、猫尾草单播的播量为15kg/hm2,鸭茅、无芒雀麦单播的播量为30kg/hm2,红豆草(Onobrychisviciaefolia)单播的播量为60kg/hm2;混播比例按种子占单播重量的实际用价来计算,混播与单播密度相同,混播播量及播种比例见表1。2008年5月4日播种,播种当年在8月20日刈割1次。翌年返青后不施用任何肥料,试验期间不灌溉,中耕松土1次,每年人工除杂草2次。
1.3 评价指标的选择
1.3.1 指标的选择 禾草在刈割或放牧下,表现出较强的再生性,而豆科植物再生性较弱[16]。因此,在豆禾混播群落中,组分的稳定性可理解为豆科植物保持较高的比例,故以混播群落中禾草与豆科植物的相对密度(relative density,RD)及其年际间的变异系数衡量群落组分的稳定性。
禾草相对密度:RDg=Dgl/pDg。式中,Dgl是在混播条件下禾草分蘖数,Dg是单播条件下禾草的分蘖数,p是禾草在混播地中的播种比例。
豆科牧草相对密度:RDl=Dll/qDl。式中,Dll是混播条件下豆科牧草的分枝数,Dl是单播条件下豆科牧草的分枝数,q是豆科牧草在混播地中的播种比例。
RDg=1表明种内竞争和与豆科牧草的种间竞争对禾草种群数量影响相似;RDg>1表明禾草在与豆科牧草混播时扩大了自己的种群;时,表明豆科牧草的存在限制了禾草种群数量的扩展;值含义同[17]。
变异系数:CVi=(Vi/X)×100%。式中,Vi为各植物种相对密度的标准差,X为各植物种相对密度的平均值。
以牧草产量、牧草营养物质产量及其年际间变异系数衡量群落功能的稳定性。变异系数公式同上。
Davis等[18]认为植物群落中可用(或未曾利用)资源的增加都将使该群落的可入侵性(或对于入侵的易感性)增加;如果群落处于高可入侵性,必定有可用的资源。相对产量总和(relative yield total,RYT)是物种对已占有资源量的评价[19-20],故以相对产量总和衡量群落可入侵性。
相对产量总和:RYT=(Yij/Yii)+(Yji/Yjj),式中,Yij为种i与种j混播时种i的草产量;Yii为种i单播时的草产量;Yji为种j同种i混播时种j的草产量;Yjj为种j单播时的草产量[21]。RYT=1,说明在该组分下种间和种内干扰相等;RYT<1,说明在该组分下种间干扰大于种内干扰,环境资源没有得到充分利用;RYT>1说明种间干扰小于种内干扰,各个竞争种可能有某种程度的生态位分化,环境资源得到充分利用。
1.3.2 评价方法 对参评各因子值进行标准化处理,将每一数值与该参数的最大值相比,再乘以1000,即得标准化值[11]。应用模糊数学的隶属函数法对21种豆禾混播群落稳定性进行综合评价。采用的评价模型为U(Xik)=(Xik-Xkmin)/(Xkmax-Xkmin),其中,U(Xik)为第i种混播群落第k项指标的隶属度,Xik为评价指标的标准值或多项参评因子标准值的平均值;Xkmax、Xkmin分别为所有混播群落第k项指标最大值和最小值。以群落组分的稳定性、群落功能的稳定性和群落可入侵性3项指标隶属度的均值评价各混播群落稳定性大小的依据。
1.4 取样时间和样品处理
每年刈割1次,均在豆科牧草现蕾期、禾草初花期进行刈割(2008年在8月15日,2009-2012年刈割时间在6月20日左右)。测定各混播组分产量和分枝/分蘖数,并计算相对密度、相对产量总和,在整个研究期间共测定5次。禾草相对密度以分蘖数计数,豆科牧草相对密度以一级分枝数计数;草产量的测定从小区的一端开始按1 m×1m的样方(缺苗及边行不取样)取样,3次重复,样方内牧草齐地面刈割并称鲜重,取500g鲜样放入65℃恒温箱烘干称重,计算鲜干比,折算干草产量。

表1 豆禾混播草地混播比例与播量Table 1 Mixed sowing ratio and sowing quantity in legume-grass mixture
每次刈割的同时,按“十”字法随机取样500g混匀,将样品粉碎后,过0.4mm筛,利用ZDDN-Ⅱ型凯氏定氮仪测定牧草样品的粗蛋白含量(crude protein,CP)、SZF-06A 型粗脂肪仪测定粗脂肪含量(ether extract,EE)、CXC-06纤维分析仪测定中性洗涤纤维(neutral detergent fiber,NDF)含量,并根据产草量换算成CP、EE和NDF产量,具体操作参见杨胜[22]的方法。
1.5 数据处理
利用IBM SPSS Statistics 21中的One-way ANOVA对不同年份的各评价指标进行方差分析、计算标准误,并利用LSD对指标进行年际间比较,差异显著性水平P<0.05。
2 结果与分析
2.1 不同混播组合群落组分稳定性的比较
从各个年份来看,2008年组合G、H、I的RDg值较高,而组合D、E、F较低;2009年组合I、P的RDg较高,而组合J、K、L、O较低;2010年组合M、T的RDg值较高,而组合C、I较低;2011年组合L、P的RDg值较高,而组合B、E、I较低;2012年组合D、P的RDg值较高,而组合B、E、I、Q较低(图1)。从年际间的变化来看,组合A、B、C、D、E、F、I、P、Q、R、S、T的RDg呈单峰变化趋势,在2009年到达最高值,以后逐年下降;而组合G、H、J、K、M、N、O、U的RDg呈直线下降趋势,在2008年达到最高值,以后逐年下降(图1)。组合A、T的RDg距平较小,且RDg值均在1.90以上,说明其RDg变化幅度较小,禾草在群落中具有较高的稳定性(图2)。从5年均值来看,组合I、P具有较高RDg值,且具有较低的RDg变异系数;所有组合RDg值均大于1(图3)。
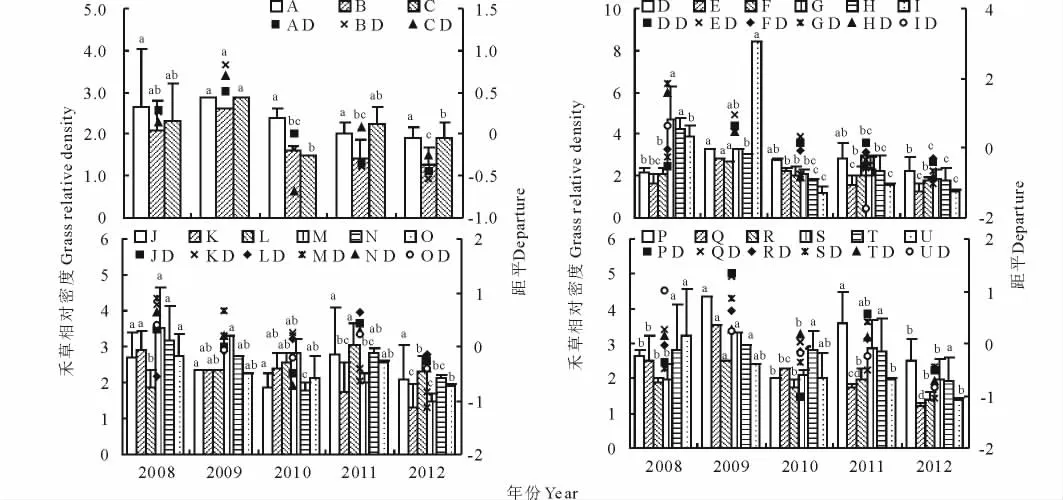
图1 不同混播组合禾草相对密度及距平的变化Fig.1 Changes of grass relative density and departure among different mixed patterns
从各个年份来看,2008年组合C、F、I、J的RDl值较高,而组合S、U较低;2009年组合C、F、O的RDl值较高,而组合G、P较低;2010年组合B、C、I的RDl值较高,而组合R、S较低;2011、2012年组合F、J、L的RDl值较高,而组合T、U的RDl值为0,表明其豆科植物完全消失(图2)。从年际间的变化来看,组合A、B、C、D、F、G、H、I、J、K、L、P、Q、R、的RDl呈直线下降趋势,在2008年到达最高值,以后逐年下降;而组合E、M、N、O、S、T、U 的RDl呈单峰变化趋势,在2009年达到最高值,以后逐年下降(图2)。组合A、B、M的RDl距平较小,且RDl值均在1.10以上,说明其RDl变化幅度较小,豆科植物在群落中具有较高的稳定性。从5年均值来看,组合F、L具有较高的RDl值,但具有较高的RDl变异系数;组合S、T、U的RDl值小于1,其他组合均大于1(图3)。

图2 不同混播组合豆科相对密度及距平的变化Fig.2 Changes of legume relative density and departure among different mixed patterns

图3 不同混播组合禾草与豆科植物相对密度的5年均值及变异系数Fig.3 Means and CV of grass and legume relative density among different mixed patterns in 2008-2012
2.2 不同混播组合群落生产性能稳定性的比较
从各个年份来看,2008年组合I、M牧草产量较高,而组合C、D较低;2009年组合S、T、U牧草产量较高,而组合D、K较低;2010年组合K、Q牧草产量较高,而组合S、T较低;2011年组合C、P牧草产量较高,而组合M、Q较低;2012年组合A、I牧草产量较高,而组合S、T较低(图4)。从年际间的变化来看,组合A、D、E、F、J、K、L、P、Q、R的牧草产量呈单峰变化趋势,峰值出现在2010年;而组合B、C、G、H、I、M、N、O、S、T、U的牧草产量也呈单峰变化趋势,但峰值出现在2009年(图4)。从产量距平来看,组合D、E、F、J、K、L、P、Q、R的牧草产量较低,其牧草产量距平也较小;而组合G、H、I、M、N、O、S、T、U牧草产量较高,其牧草产量距平也较大;组合A、B、C牧草产量较高,其牧草产量距平也较小,说明其牧草产量变化幅度较小,在产量方面具有较高的稳定性。
牧草营养物质产量取决于牧草产量及其营养物质含量,因此与牧草产量紧密关联。从各个年份来看,2008年组合L、M、P等CP、EE和NDF产量较高,而组合B、C、D较低;2009年组合S、T、U等CP、EE产量较高,组合D、K较低,组合H、M、O等NDF产量较高,组合J、K、L较低;2010年组合I、K、Q等CP、EE和NDF产量较高,而组合L、M、S等较低;2011年组合C、P等CP、EE和NDF产量较高,而组合 M、Q较低;2012年组合A、I等CP、EE和NDF产量较高,而组合S、T较低;其变化规律与牧草产量的基本一致(图5,图6,图7)。CP、EE和NDF产量的年际变化规律和产量距平也与牧草产量的基本一致(图5,图6,图7)。

图4 不同混播组合牧草产量及距平的变化Fig.4 Changes of forage yield and departure among different mixed patterns

图5 不同混播组合粗蛋白产量及距平的变化Fig.5 Changes of crude protein yield and departure among different mixed patterns

图6 不同混播组合粗脂肪产量及距平的变化Fig.6 Changes of ether extract yield and departure among different mixed patterns

图7 不同混播组合中性洗涤纤维产量及距平的变化Fig.7 Changes of neutral detergent fiber yield and departure among different mixed patterns
从牧草产量和营养物质产量5年均值来看,组合C具有较高牧草产量,且具有较低的牧草产量变异系数;所有组合牧草产量均大于350g/m2;组合C、I、O具有较高的CP、EE和NDF产量,且C具有较低的变异系数(图8)。
2.3 不同混播组合群落入侵性稳定性的比较
从各个年份来看,2008年组合R、S、T等相对总产量(RYT)值较高,而组合D、E、P较低;2009年组合B、C等RYT值较高,组合D、K、P较低;2010年组合I、K、Q等RYT值较高,而组合H、R、S等较低;2011年组合C、J、P等RYT值较高,而组合Q、R、U较低;2012年组合A、I、L等RYT值较高,而组合S、T、U较低;2008-2012年所有组合RYT值均高于1(图9)。从年际间的变化来看,组合A、D的RYT值呈逐年上升趋势,在2012年出现峰值;组合B、C、E、H、L、M、N的RYT值呈双峰变化趋势,在2009和2012年出现了2个峰值;组合F、J、O、P、Q、U的RYT值呈单峰变化趋势,在2009、2010或2011年出现峰值;组合G、I、K的RYT值也呈双峰变化,但其峰值出现在2010和2012年;组合R、S、T的RYT值呈逐年下降趋势,峰值出现在2008年(图9)。从5年均值来看,组合A、C、I、J、L具有较高的RYT值,且A具有较低的变异系数(图9)。

图8 不同混播组合牧草产量及营养物质产量5年均值与变异系数Fig.8 Means and CV of forage,crude protein,ether extract and neutral detergent fiber yield among different mixed patterns in 2008-2012
2.4 模糊综合评价
综合统计各个混播组合群落稳定性评价指标的标准值或多项参评因子标准值的平均值,分别计算群落组分稳定性、群落功能稳定性和群落可入侵性的隶属度,以这3个隶属度的均值综合评价各混播组合的群落稳定性。由表2可知,组合C、D、F、J、L、M、T的稳定性较高,其隶属度在0.45~0.50之间;组合 H、A、R的稳定性较低,其隶属度均小于0.38;其他组合稳定性处于中间,其隶属度介于0.40~0.45(表2)。

表2 豆禾混播草地群落稳定性指标的隶属度及其均值Table 2 Subordinate function values of three stability indexes and their means of different mixed patterns
3 讨论
3.1 混播种类和比例对群落稳定性的影响
稳定性是植物群落结构与功能的一个综合特征,生态学家一直试图发展多样性与稳定性关系的通用理论[23],并将物种或功能群数量作为群落稳定性的重要特征[24-25]。虽然也有很多学者认为物种或功能群数量的增加导致群落稳定性增加需要考虑一定前提[26],或者说真正导致稳定性的不是物种或功能群数量,而是群落功能群的能力[27]。从人工草地群落的角度去考虑,由多个种群组成的群落要比单一种群群落更能有效地利用环境资源,维持长期较高的生产力和稳定性[28]。但人工草地物种间的互补对生产力的正效应被认为在很低的物种丰富度水平上就可以达到饱和[29],因此豆禾混播草地不可能无限地追求混播种类的多样性[30]。本研究中,随着混播种类数量的增加,牧草产量的隶属度呈增加趋势(混3为0.41,混4为0.42,混5为0.43,混6为0.45);但综合了组分稳定性、功能稳定性和群落可入侵性的群落整体稳定性却不是上述趋势,而是随着混播种类数量的增加,其群落稳定性呈下降趋势(表2),这与安丽娟等[10]、高润梅等[11]研究结果一致。因此,物种多样性可能仅仅表征了群落生产力这一方面的特征,无法由物种多样性高低推断群落稳定性的高低。另外,豆禾比为4∶6的牧草产量隶属度小于豆禾比5∶5和3∶7,豆禾比为4∶6的禾草RD隶属度大于其他混播比例,豆科植物RD隶属度随豆禾比的增加呈下降趋势,而综合隶属度3者之间无差异。由此可见,混播种类与混播比例分别影响的是群落稳定性的某一方面,并不能简单依据混播种类的多少、混播比例的高低来推断群落稳定性的高低。
3.2 豆禾混播草地群落稳定性的时间尺度特征
豆禾混播草地随着利用年限的延长,群落组分、结构和功能不可避免的会发生变化。对于自然生态系统而言,这种群落内部的变化往往不与群落稳定性联系起来[8,31-32],也有学者认为不同的演替阶段也存在稳定性的问题[33],各演替阶段的稳定性取决于生境和干扰格局[11]。对于豆禾混播草地而言,群落内部组分的消长、更替,以及由此产生的生产功能变化是其稳定性变化的根源,外界的干扰(相应的农作措施)则是阻止这种变化,维持其农艺性状稳定的保障。那么依据时间序列,以各指标的变异系数和弹性(距平)分析得到的群落稳定性可以看成是混播种类、混播比例与刈割干扰综合影响下的群落抵抗力和恢复力。而且这种稳定性是具有一定时间尺度的:在较短时间尺度上(5年以内)生产功能(牧草产量、营养物质产量)和豆科植物变化明显,而群落可入侵性和禾草稳定性较高(图1,图3,图9);在较长时间尺度上(5年以上)群落盖度能保持较高稳定性[12]。豆科植物的竞争力和耐刈割性均弱于禾草[14],在群落中属于竞争力弱、持久性差的组分,而禾草属于竞争力强,持久性好的组分。由此可见,群落在较短时间尺度上表现出的组分、功能不稳定性主要是豆科植物消长造成的。因此,在需要建植稳定性较高的豆禾混播草地时,需要考虑豆科植物的竞争力、持久性(生长寿命)及适应性等问题。而在评价豆禾混播草地稳定性时,时间尺度及相应的评价指标显得很重要:在较短时间尺度上,需要群落内组分及其功能变化的指标;在较长时间尺度上,则需要整个群落特征及其功能变化的指标,这样才能保证进行稳定性的比较时,所受干扰的时间尺度与讨论稳定性概念所隐含的时间尺度是一致的[2]。
4 结论
从群落组分稳定性、功能稳定性和可入侵性3个方面测度与比较了21个豆禾混播组合的群落稳定性。从组分稳定性来看,含红豆草的混5-2、混4-2、混3-2等组合具有较低的豆科RD隶属度,豆科植物稳定性较差;而不含红豆草的混5-1、混4-1、混3-1等组合具有较高的豆科RD隶属度,豆科植物稳定性较高。从功能稳定性来看,随着混播种类数量的增加,牧草产量、CP产量、EE产量等的隶属度呈增加趋势,豆禾比为4∶6的牧草产量、CP产量、EE产量等隶属度小于豆禾比5∶5和3∶7。从可入侵性来看,混播种类对其影响较小,豆禾比5∶5具有较高的RYT隶属度。综合组分稳定性、功能稳定性和可入侵性3个方面来看,豆禾比为4∶6具有较低的群落稳定性,而混播种类对群落稳定性影响较小。由此可见,混播种类、混播比例仅仅能影响到群落稳定性的某一方面,对群落稳定性不起决定性的作用。如何科学测度豆禾混播草地群落的稳定性,可能还需要考虑时间尺度、空间尺度及对应的敏感指标,寻找适当的“标尺”,理清稳定性所隐含的内容,才能达到科学、准确、客观的目的。
Reference:
[1]Wang Y S,Jiang W L,Hong F Z,etal.A review of studies on stability of artificial mixed-pasture.Grassland of China,2005,27(4):58-64.
[2]Dong S K,Hu Z Z.Research advancement on the stability of artificial grassland and the mechanisms of its maintenance.Grassland and Turf,2000,3:3-8.
[3]Zhou H K,Zhao X Q,Zhao L,etal.The community characteristics and stability of theElymusnutansartificial grassland in alpine meadow.Chinese Journal of Grassland,2007,29(2):12-24.
[4]Ma C H,Han J G,Li H X,etal.The dynamical studies on biomass,qualities and interspecific competition of the rye and vetch mixture.Acta Prataculturae Sinica,1999,8(4):56-64.
[5]Zheng W,Zhu J Z,Jianaerguli.A comprehensive evaluation of the productive performance of legume—grass mixture under different mixed sowing patterns.Acta Prataculturae Sinica,2012,21(6):242-251.
[6]Xie K Y,Zhao Y,Li X L,etal.Relationships between grasses and legumes in mixed grassland:a review.Acta Prataculturae Sinica,,2013,22(3):284-296.
[7]Han D L,He S J,Chen C,etal.The community stability of the mixed grassland with leguminous and gramineous forages.Ecology and Envi-ronment,2008,17(5):1974-1979.
[8]Zhang J Y,Zhao H L.An example for study on vegetation stability in sandy desertification land:determ ination and comparison of resistance among communities under a short period of extremely aridity disturbance.Acta Ecologica Sinica,2010,30(20):5456-5465.
[9]Zheng Y R.Comparison of methods for studying stability of forest community.Scientia Silvae Sinicae,2000,36(5):28-32.
[10]An L J,Zhu Z H,Wang X A,etal.Stability analysis of major communities in malan forest region of Ziwuling Mountain.Acta Botanica Boreali-Occidentalia Sinica,2007,27(5):0859-0863.
[11]Gao R M,Shi X D,Guo Y D.Community stability evaluation of riparian forest of the upper reaches of Wenyuhe in Shanxi,China.Chinese Journal of Plant Ecology,2012,36(6):491-503.
[12]Li L,Wang Y S,Hong F Z,etal.Effects of long-term utilizing systems on community stability of Karst pasture.Ecology and Environment,2011,20(8-9):1204-1208.
[13]Gu M H,Wang T,Du G Z.Effects of fertilization on productivity and stability of cultivated grassland in an alpine region.Journal of Lanzhou University(Natural Sciences),2010,46(6):59-64.
[14]Dong S K,Kang M Y,Hu Z Z,etal.Performance of cultivated perennial grass mixtures under different grazing intensities in the alpine region of the Qinghai-Tibetan plateau.Grass and Forage Science,2004,59(3):298-306.
[15]Zhao Q S,Hou X Y,Duan J J,etal.A dynamic study on community diversity of leguminous grass mixed artificial pasture in different grazing systems.Chinese Journal of Grassland,2013,35(5):80-85.
[16]Zheng W,Zhu J Z,Kuerban,etal.Dynamics of interspecific competition of legume—grass mixture under different mixed sowing patterns.Acta Agresia Sinica,2010,18(4):568-575.
[17]Wang P,Zhou D W,Zhang B T.Coexistence and inter-specific competition in grass-legume mixture.Acta Ecologica Sinica,2009,29(5):2560-2567.
[18]Davis M A,Grime J P,Thompson K.Fluctuating resources in plant communities:ageneral theory of invisibility.Journal of Ecology,2000,88:528-534.
[19]Zhang Y L,Wang J L,Hu Z Z.Interspecific competition and community stability of variegated Alfalfa+Awnless Brome mixture.Acta Agresia Sinica,2007,15(1):43-49.
[20]Zhang H Y,Yang H S,Li C H,etal.Research on the dynamics of productive forces of Alfalfa+awnless brome man-made grassland under the different patterns of mixed sowing.Journal of Inner Mongolia University for Nationalities,2008,23(1):55-58.
[21]Jonathan W S.Introduction to Plant Population Ecology[M].Published in the United States of America,1982:147-155.
[22]Yang S.Feed Analysis and Feed Quality Detection Technology[M].Beijing:China Agricultural University Press,1999:19-61.
[23]MacArthur R H.Fluctuations of animal populations,and a measure of community stability.Ecology,1955,36:533-536.
[24]Tilman D,Downing J A.Biodiversity and stability in grassland.Nature,1994,367(27):363-365.
[25]Bengtsson J.Which species?What kind of diversity?Which ecosystem function?Some problems in studies of relations between biodiversity and ecosystem function.Applied Soil Ecology,1998,10:191-199.
[26]Zhang L M,Chen B,Li Z Y.Analysis of the species diversity and community stability in local-community using the Neutral Theory.Acta Ecologica Sinica,2010,30(6):1556-1563.
[27]Cardinale B J,Srivastava D S,Duffy J E,etal.Effects of biodiversity on the functioning of trophic groups and ecosystems.Nature,2006,443:989-992.
[28]Li B.Ecology[M].Beijing:Higher Education Press,1999:233-237.
[29]Vandermeer J H.The Ecology[M].Cambrige University Press,1989:98-102.
[30]Jiang W L,Chu A C P,Ren J Z.Botanical composition dynamics and herbage mass in pasture mixtures in the mountainous regions of Guizhou province,China.Proceedings of theⅩⅦInternational Grassland Congress[C].New Zealand and Australia,1994:72-75.
[31]Li Q,Zhou D W,Song Y T.The distribution features of two widespread legumes and their relationships with soil factors in Songnen grassland.Acta Prataculturae Sinica,2014,23(1):31-40.
[32]Wang J B,Zhang D G,Cao G M,etal.Regional characteristics of the alpine meadow degradation succession on the Qinghai—Tibetan Plateau.Acta Prataculturae Sinica,2013,22(2):1-10.
[33]Dang C L,Wang C Y,Wang B R,etal.Succession and stability in plant community.Chinese Journal of Ecology,2002,21(2):30-35.
[1]王元素,蒋文兰,洪绂曾,等.人工混播草地群落稳定性研究进展.中国草地,2005,27(4):58-64.
[2]董世魁,胡自治.人工草地群落稳定性及其调控机制研究现状.草原与草坪,2000,3:3-8.
[3]周华坤,赵新全,赵亮,等.高山草甸垂穗披碱草人工草地群落特征及稳定性研究.中国草地学报,2007,29(2):12-24.
[4]马春晖,韩建国,李鸿祥,等.冬牧70黑麦+箭筈豌豆混播草地生物量、品质及种间竞争的动态研究.草业学报,1999,8(4):56-64.
[5]郑伟,朱进忠,加娜尔古丽.不同混播方式豆禾混播草地生产性能的综合评价.草业学报,2012,21(6):242-251.
[6]谢开云,赵云,李向林,等.豆-禾混播草地种间关系研究进展.草业学报,2013,22(3):284-296.
[7]韩德梁,何胜江,陈超,等.豆禾混播草地群落稳定性的比较.生态环境,2008,17(5):1974-1979.
[8]张继义,赵哈林.短期极端干旱事件干扰下退化沙质草地群落抵抗力稳定性的测度与比较.生态学报,2010,30(20):5456-5465.
[9]郑元润.森林群落稳定性研究方法初探.林业科学,2000,36(5):28-32.
[10]安丽娟,朱志红,王孝安,等.子午岭马栏林区主要森林群落的稳定性分析.西北植物学报,2007,27(5):0859-0863.
[11]高润梅,石晓东,郭跃东.山西文峪河上游河岸林群落稳定性评价.植物生态学报,2012,36(6):491-503.
[12]李莉,王元素,洪绂曾,等.喀斯特地区长期草地利用制度对群落稳定性的影响.生态环境学报,2011,20(8-9):1204-1208.
[13]顾梦鹤,王涛,杜国桢.施肥对高寒地区多年生人工草地生产力及稳定性的影响.兰州大学学报(自然科学版),2010,46(6):59-64.
[15]赵青山,侯向阳,段俊杰,等.不同放牧制度下混播人工草地群落多样性动态.中国草地学报,2013,35(5):80-85.
[16]郑伟,朱进忠,库尔班,等.不同混播方式下豆禾混播草地种间竞争动态研究.草地学报,2010,18(4):568-575.
[17]王平,周道玮,张宝田.禾-豆混播草地种间竞争与共存.生态学报,2009,29(5):2560-2567.
[19]张永亮,王建丽,胡自治.杂花苜蓿与无芒雀麦混播群落种间竞争及稳定性.草地学报,2007,15(1):43-49.
[20]张宏宇,杨恒山,李春辉,等.不同混播方式下苜蓿+无芒雀麦人工草地生产力动态研究.内蒙古民族大学学报(自然科学版),2008,23(1):55-58.
[22]杨胜.饲料分析及饲料质量检测技术[M].北京:中国农业大学出版社,1999:19-61.
[26]张立敏,陈斌,李正跃.应用中性理论分析局域群落中的物种多样性及稳定性.生态学报,2010,30(6):1556-1563.
[28]李博.普通生态学[M].北京:高等教育出版社,1999:233-237.
[31]李强,周道玮,宋彦涛.松嫩草地两种广布豆科植物分布特征及其与土壤因子关系.草业学报,2014,23(1):31-40.
[32]王建兵,张德罡,曹广民,等.青藏高原高寒草甸退化演替的分区特征.草业学报,2013,22(2):1-10.
[33]党承林,王崇云,王宝荣,等.植物群落的演替与稳定性.生态学杂志,2002,21(2):30-35.
