室温干燥贮藏和天然高寒草甸层积对草玉梅种子萌发的影响
2015-01-02鱼小军王芳张婧徐长林肖红景媛媛张建文杨海磊
鱼小军,王芳,张婧,徐长林,肖红,景媛媛,张建文,杨海磊
(1.甘肃农业大学草业学院,草业生态系统教育部重点实验室,中-美草地畜牧业可持续发展研究中心,甘肃省草业工程实验室,甘肃 兰州730070;2.甘肃农业大学农学院,甘肃 兰州730070)
种子休眠是延迟植物繁殖、增大物种适合度的一种生态策略,是种子内在的固有特性之一,休眠的主要生态学意义在于可以防止种子在适宜萌发但不利于幼苗建植的环境条件下萌发,从而确保种群在风险环境中延续[1-3]。种子萌发是种子对环境条件和萌发需求二者叠合时的一种响应[4-5]。种子萌发阶段是植物幼苗成功建植的关键环节,是植物生长周期的关键时期,它直接影响着幼苗存活、个体适合度及植物生活史的表达,进而影响到种群动态和群落组成[6-9]。
种子休眠主要包括由种胚的形态后熟、生理后熟引起的胚休眠,由种壳(种皮、果皮)透水性、透气性和透光性差等因素引起的物理休眠,以及由种胚和种壳共同作用引起的复合休眠[1,5]。干贮藏能影响种子的休眠水平,控制休眠解除的速率[10]。一些植物的种子经干贮藏后才能完成后熟(after-ripening),发芽率才得以提高。骆驼蒿(Peganumnigellastrum)种子经室温干燥贮藏6个月,其发芽率由贮藏前的8%提高到98%[11];10月份成熟的长枝木蓼(Atraphaxisspinos)种子贮藏3,6,9个月后,发芽率由贮藏前的17%提高到99%[10]。层积(stratification或chilling)是一种古老而有效的破除种子休眠的方法[12]。冷层积处理高寒草甸植物种子后约有50%的种子发芽率显著增大[13]。种子萌发对寒冷的需求代表着植物的一种生存对策,可以保证种子在冷季之后的春季或早夏萌发,这有利于幼苗的存活[1,14]。层积处理为种子发芽提供了有利条件,已在打破多种植物种子休眠上取得了较好效果[11-13,15-16]。
草玉梅(Anemonerivularis)又名溪畔银莲花、虎掌草(云南)、鬼打青(四川)和水乌头(湖北)等,为毛茛科多年生草本植物[17-20]。草玉梅的根状茎和叶具清热解毒、活血舒筋之功效,可用于治疗咽喉痛、风湿痛、胃痛、疟疾、慢性肝炎和肝硬化等症[21-22];另外,草玉梅也是青藏高原高寒草甸的重要植物种,因此对草地生态系统和生物多样性的稳定、家畜的健康生产具有重要作用[23-24]。有研究表明,由于草玉梅的果皮遮光引起了草玉梅种子的休眠,除去果皮能使室温贮藏2个月的草玉梅种子在光照条件下的发芽率提高到76.0%;除去果皮后尚有部分种子仍然处于休眠状态,因此草玉梅种子(瘦果)的休眠为综合休眠类型[25-28]。然而,各种贮藏条件对草玉梅种子萌发行为的影响尚未见研究报道。这些基础信息的缺乏将不利于高寒草甸草原管理、退化草地恢复和草玉梅的栽培工作。本文通过研究室温干燥贮藏和天然高寒草甸层积对草玉梅种子萌发的影响,揭露其生活史特征,为高寒草甸草原管理、退化草地恢复和草玉梅的栽培工作提供依据。
1 材料与方法
1.1 试验材料
于2013年10月初,在甘肃省武威市天祝藏族自治县抓喜秀龙乡的甘肃农业大学天祝高山草原试验站(37°40′N,102°32′E,海拔2960m)采集完熟期的草玉梅种子(瘦果)。该地区草地为寒温潮湿类高寒草甸;该地年均气温-0.1℃,最热月7月均温12.7℃,最冷月1月均温-18.3℃,≥0℃的年积温为1380℃。年均降水量416mm,多集中于7、8、9月;年蒸发量1592mm,土壤为高山草甸土,土层厚度40~80cm,土壤pH为7.0~8.2[29]。供试完熟期的草玉梅种子在母株上容易脱落,果皮呈黑色,含水量为13.6%。
1.2 试验设计
1.2.1 室温干燥贮藏处理 草玉梅种子采集后于甘肃农业大学草业学院草地资源与生态实验室干燥贮藏,室温为20℃。分别在种子采集后0,40,80,150和180d,于25/15℃的变温条件下进行发芽试验。进行6个处理,分别为去果皮光照(光照强度为1250lx)、去果皮自然光照、去果皮黑暗,不去果皮光照、不去果皮自然光照、不去果皮黑暗。精选大小一致、籽粒饱满、无破损的草玉梅种子进行试验。黑暗条件下以脱脂棉加单层滤纸为发芽床,其余条件下以单层滤纸为发芽床。
1.2.2 高寒草甸层积处理 于2013年10月底布设在天祝高寒草甸围栏内的不放牧高寒草甸为层积不践踏处理,以及自由中度放牧的冷季牧场层积践踏处理,中度放牧强度主要参考任继周[30]和中华人民共和国农业部发布的农业行业标准“天然草地合理载畜量的计算”[31]确定,并以同期室温干燥贮藏的草玉梅种子为对照。层积处理种子埋深1cm[32],层积种子于2014年4月初取回部分种子,分别在30/20℃,25/15℃,20/10℃,20/5℃和15/5℃条件下进行发芽试验。
践踏处理的草玉梅种子50%种子留于草地,待2014年8月20日,统计草地上的发芽数和未发芽种子数。将未发芽种子带回实验室,分别在30/20℃,25/15℃,20/10℃,20/5℃和15/5℃条件下进行发芽试验。
所有发芽试验均采用纸上(TP)法,每培养皿50粒,每处理3次重复。每日统计发芽数,每天加入适量的蒸馏水,第18天结束发芽试验。
1.3 数据统计分析
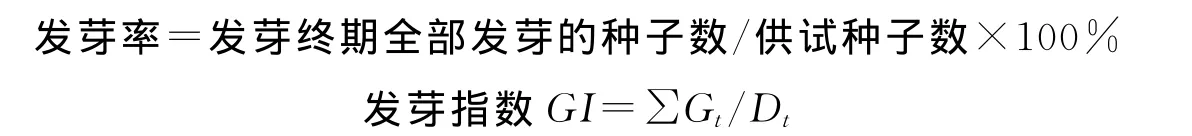
式中,Gt为t天发芽的种子数;Dt为相对应的种子发芽天数。
所有数据均以平均数±标准误表示,试验数据用SPSS 16.0进行差异显著性统计分析。
2 结果与分析
2.1 室温干燥贮藏对草玉梅种子萌发的影响
2.1.1 对初次发芽天数的影响 草玉梅种子的初始发芽天数因室温干燥贮藏的天数不同、是否去掉果皮和是否光照而异(表1)。
随着贮藏天数的增加,去掉果皮草玉梅种子的初始发芽天数呈不同程度的减小。去掉果皮的草玉梅种子,不论是光照、培养箱自然光,还是黑暗条件下,贮藏40~180d处理的初次发芽天数显著低于贮藏0d处理(P<0.05)(表1)。在光照或自然光照的情况下,去掉果皮的草玉梅种子初次发芽天数,贮藏40,80,150和180d间的差异不显著(P>0.05)。去掉果皮的草玉梅种子,在黑暗条件下,贮藏180d的初次发芽天数均显著小于贮藏40,80和150d的处理(P<0.05)。没有去掉果皮的草玉梅种子,光照条件下,贮藏40~180d处理的初次发芽天数均显著低于贮藏0d处理(P<0.05)(表1);自然光情况下,贮藏40和80d处理的初次发芽天数和对照差异不显著,贮藏150和180d处理的初次发芽天数均显著低于对照和贮藏40,80d处理;黑暗条件下,0~150d的贮藏处理下的草玉梅种子均在第18天未能发芽,贮藏180d的处理在17d有种子发芽。
光照不同程度地降低了去掉果皮草玉梅种子的初始发芽天数。刚收获的去掉果皮的草玉梅种子,在光照条件下的初始发芽天数为14d,显著低于自然光(>18d)和黑暗处理(>18d)(P<0.05)(表1)。经过40~180d的室温干燥贮藏,光照和自然光照情况下去掉果皮草玉梅种子初始发芽天数间的差异不显著(P>0.05),但均显著低于黑暗处理(P<0.05)。贮藏40~180d后,未去掉果皮的草玉梅种子,光照条件下的初始发芽天数均显著小于自然光照和黑暗处理(P<0.05)。贮藏0~80d,未去掉果皮的草玉梅种子在自然光照和黑暗条件下初始发芽天数间的差异不显著,均为>18d。
除了贮藏0d的自然光照和黑暗处理,其他去掉果皮处理均显著降低了草玉梅种子的初始发芽天数(P<0.05)(表1)。同为光照条件,贮藏40,80,150和180d后去掉果皮草玉梅种子的初次发芽天数分别为第7.0,6.0,6.0和5.7天,显著高于贮藏同期未去掉果皮处理(初次发芽天数分别为14.3,12.0,12.0和13.0d)(P<0.05)。同为自然光照条件,贮藏40,80,150和180d后去掉果皮草玉梅种子的初次发芽天数分别为第8.3,6.0,6.0和6.0天,贮藏同期未去掉果皮处理的初次发芽天数分别为>18.0,>18.0,15.0和15.0(P<0.05),前4者皆显著低于后4者(P<0.05)。同为黑暗条件,贮藏40,80,150和180d后去掉果皮草玉梅种子的初次发芽天数分别为第10.3,10.3,10.0和9.0天,贮藏同期未去掉果皮处理的初次发芽天数分别为>18.0,>18.0,>18.0和17.0,前4者皆显著低于后4者(P<0.05)。
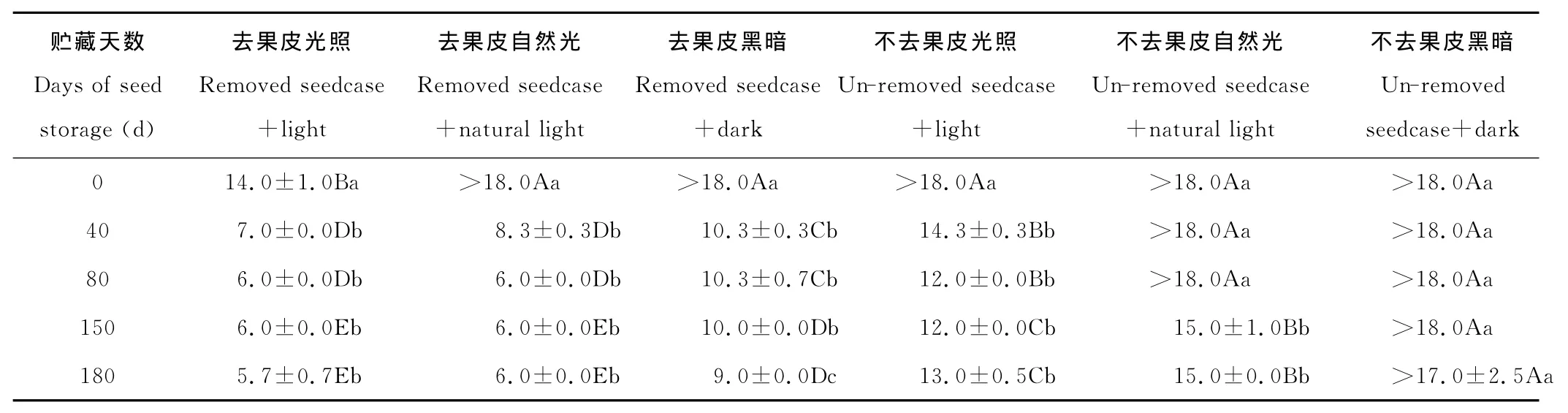
表1 室温贮藏不同天数下草玉梅种子的初次发芽天数Table 1 The initial germination day of A.rivularis seeds stored at room temperature for different days d
2.1.2 对发芽率的影响 同初次发芽天数一致,草玉梅种子的发芽率因室温干燥贮藏天数不同、是否去掉果皮和是否光照而异(表2)。
随着室温干燥贮藏天数的增加,去掉果皮草玉梅种子的发芽率呈不同程度的增加(表2)。室温干燥贮藏40,80,150和180d的草玉梅种子在去皮光照处理下发芽率间的差异不显著(P>0.05),但皆显著高于贮藏0d处理(P<0.05)(表2)。室温干燥贮藏180d的草玉梅种子在去掉果皮自然光处理下的发芽率为72.7%,显著高于室温干燥贮藏40,80和150d后同条件处理(P<0.05),后三者间差异不显著(P>0.05),但皆显著高于贮藏0d处理(P<0.05)。不同贮藏天数的草玉梅种子在去皮黑暗处理下的发芽率与去掉果皮自然光处理相似。未去掉果皮贮藏180d的草玉梅种子,光照条件下的发芽率显著高于贮藏80和150d处理(P<0.05),后两者显著高于贮藏40d处理(P<0.05),贮藏40d的又显著高于贮藏0d的处理(P<0.05)。贮藏150和180d的草玉梅种子,在未去掉果皮自然光照下的发芽率为2.0%和16.7%,显著高于贮藏0,40和80d处理(P<0.05)。贮藏0~180 d的草玉梅种子,未去果皮并在黑暗下培养,能发芽的只有贮藏180d的处理,贮藏0~150d的处理均未能发芽。
光照不同程度地提高了去掉果皮草玉梅种子的发芽率(表2)。贮藏0d的草玉梅种子,去掉果皮光照下的发芽率为2%,显著高于去果皮自然光和黑暗处理(P<0.05),后两者发芽率皆为0。贮藏40,80,150和180d的草玉梅种子,去掉果皮光照和自然光处理下的发芽率,皆显著高于去掉果皮黑暗处理(P<0.05),同期去掉果皮光照和自然光处理下的发芽率差异不显著(P>0.05)。贮藏40,80,150和180d的草玉梅种子,未去掉果皮光照处理下的发芽率显著高于自然光和黑暗处理(P<0.05);贮藏0~80d未去掉果皮的草玉梅种子在自然光照和黑暗处理下发芽率为0。
除了贮藏0d的自然光照和黑暗处理,其他去掉果皮处理均显著提高了草玉梅种子的发芽率(P<0.05)(表2)。贮藏40,80,150和180d后去掉果皮草玉梅种子光照条件下的发芽率分别为63.3%,63.3%,60.0%和65.3%,显著高于贮藏同期的未去掉果皮光照处理(其发芽率分别为8.7%,11.0%,14.7%和39.3%)(P<0.05)。同为自然光照条件,贮藏40,80,150和180d后去掉果皮草玉梅种子的发芽率分别为57.3%,53.0%,53.0%和72.7%,显著高于贮藏同期的未去掉果皮处理(其发芽率分别为0.0%,0.0%,2.0%和16.7%)(P<0.05)。同为黑暗条件,贮藏40,80,150和180d后去掉果皮草玉梅种子的发芽率分别为9.3%,10.7%,12.7%和28.0%,显著高于贮藏同期的未去掉果皮处理(P<0.05)。
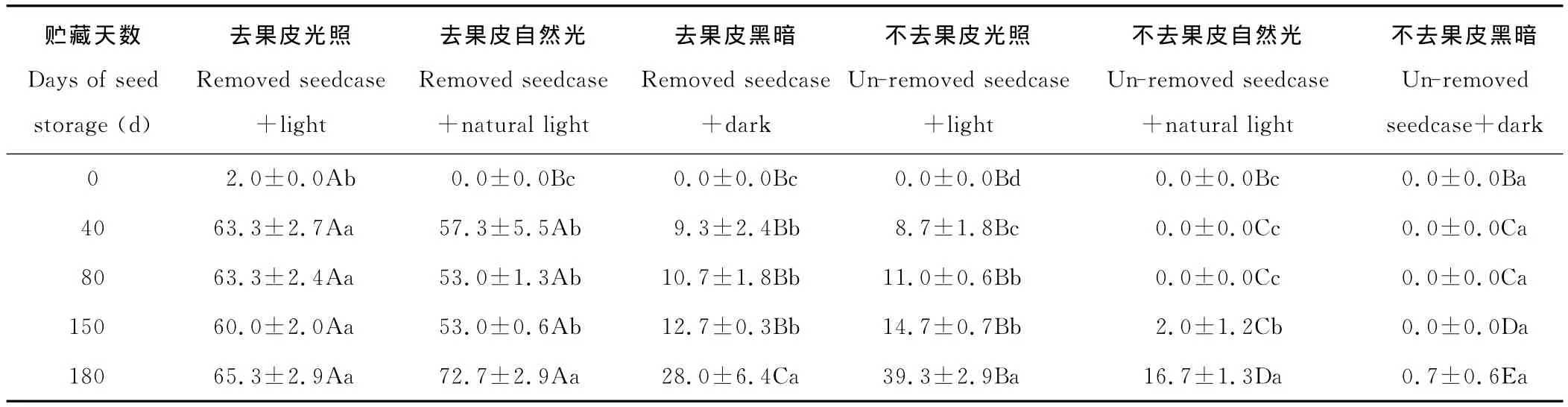
表2 室温贮藏不同天数下草玉梅种子的发芽率Table 2 The germination percentage of A.rivularis seeds stored at room temperature for different days %
2.1.3 对发芽指数的影响 草玉梅种子的发芽指数因室温干燥贮藏天数不同、是否去掉果皮和是否光照而异(表3)。
随着室温干燥贮藏天数的增加,去掉果皮草玉梅种子的发芽指数呈不同程度的增加(表3)。贮藏80,150和180d的草玉梅种子去皮光照处理下的发芽指数分别为3.29,3.23和3.42,三者间差异不显著(P>0.05);但皆显著高于贮藏40和0d处理(P<0.05),贮藏40d处理显著高于贮藏0d处理(P<0.05)。贮藏180d的草玉梅种子去果皮自然光、去果皮黑暗、未去果皮光照处理下的发芽指数显著高于贮藏40,80和150d处理(P<0.05),后三者皆显著高于贮藏0d的处理(P<0.05)。贮藏150和180d的草玉梅种子未去果皮自然光照处理下的发芽指数显著高于0,40和80d处理(P<0.05),后三者间的差异不显著。贮藏0,40,80,150和180d的草玉梅种子未去果皮黑暗处理下的发芽指数均为0。
光照不同程度地提高了草玉梅种子的发芽指数(表3)。贮藏0d的草玉梅种子,去掉果皮光照下的发芽指数为0.07,显著高于去皮自然光和黑暗处理(P<0.05),后两者发芽指数为0。贮藏0,40,80,150和180d的草玉梅种子,去掉果皮光照和自然光处理下的发芽指数,皆显著高于去掉果皮黑暗处理(P<0.05),同期的去掉果皮光照和自然光处理下的发芽指数差异不显著(P>0.05)。贮藏40,80,150和180d的草玉梅种子,未去掉果皮光照处理下的发芽指数显著高于自然光和黑暗处理(P<0.05);贮藏0~80d未去果皮的草玉梅种子,在自然光照和黑暗处理下的发芽率为0;贮藏0~180d未去掉果皮黑暗处理下的草玉梅种子发芽指数为0。
除了贮藏0d的自然光照和黑暗处理,其他去掉果皮处理均显著提高了草玉梅种子的发芽指数(P<0.05)(表3)。贮藏40,80,150和180d后去掉果皮草玉梅种子在光照条件下的发芽指数分别为2.84,3.29,3.23和3.42,显著高于贮藏同期的未去掉果皮光照处理(P<0.05)。贮藏40,80,150和180d后去掉果皮草玉梅种子在自然光照条件下的发芽指数分别为2.71,2.74,2.94和3.65,显著高于贮藏同期的未去掉果皮自然光照处理(P<0.05)。同为黑暗条件,贮藏40,80,150和180d后去掉果皮草玉梅种子的发芽指数显著高于贮藏同期未去掉果皮处理(P<0.05)。
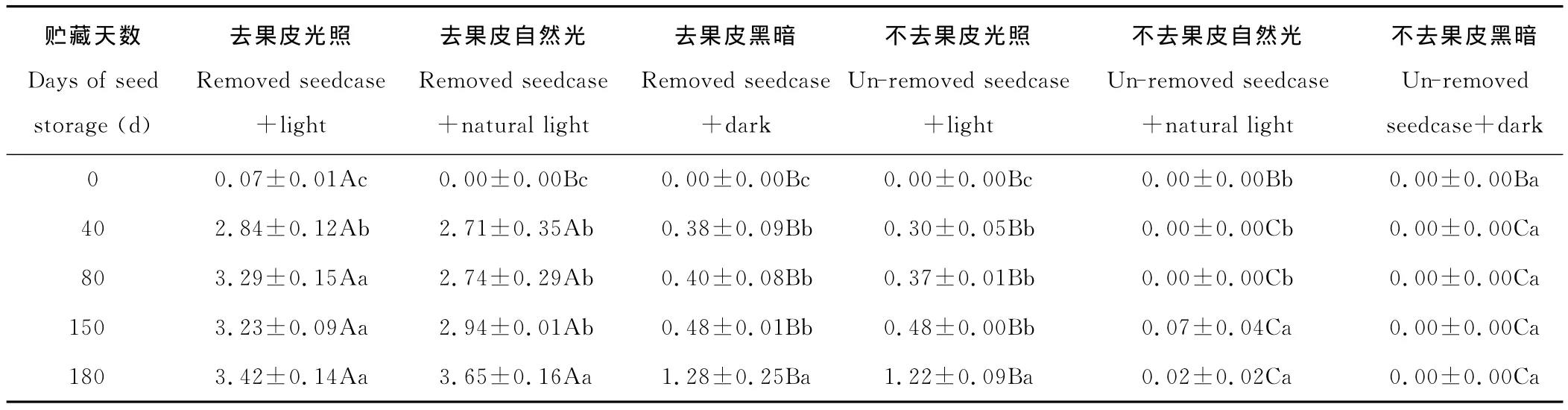
表3 室温贮藏不同天数下草玉梅种子的发芽指数Table 3 The germination index of A.rivularis seeds stored at room temperature for different days
2.2 层积于高寒草甸土壤对草玉梅种子萌发的影响
2.2.1 对初次发芽天数的影响 层积于高寒草甸土壤对草玉梅种子初次发芽天数的影响因层积时践踏与否和萌发温度不同而不同(表4)。践踏层积的草玉梅种子在30/20℃和20/10℃变温条件下的初次发芽天数分别显著低于层积不践踏处理(P<0.05),但25/15℃变温条件下的草玉梅种子初次发芽天数未受到践踏与否的影响。层积的草玉梅种子在30/20℃、25/15℃、20/10℃的初次发芽天数显著低于室温干燥贮藏同期处理(P<0.05)。
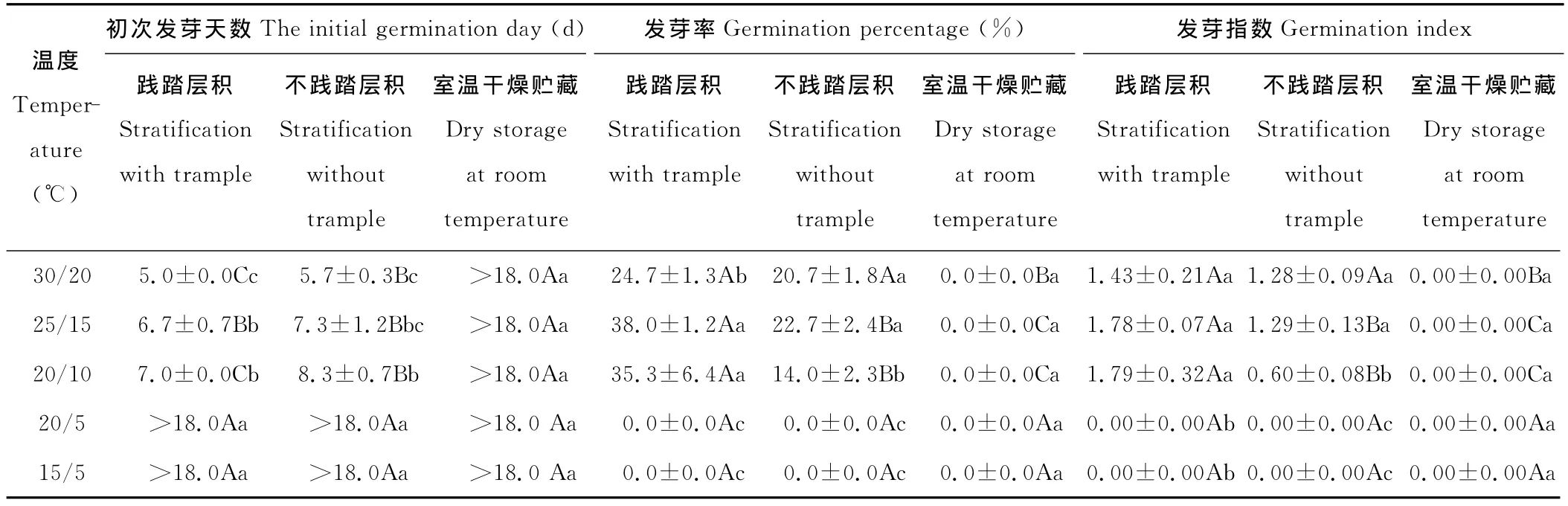
表4 层积(11月-4月)于高寒草甸的草玉梅种子在不同变温条件下的发芽率、发芽指数和初次发芽天数Table 4 The initial germination day,germination percentage and germination index of A.rivularis seeds under different alternative temperature conditions after stratification from November to April at the alpine meadow
2.2.2 对发芽率的影响 层积于高寒草甸土壤对草玉梅种子发芽率的影响因层积时践踏与否和萌发温度不同而异(表4)。践踏层积的草玉梅种子在25/15℃和20/10℃变温条件下的发芽率分别为38.0%和35.3%,显著高于层积不践踏处理(22.7%和14.0%)(P<0.05),但两处理在30/20℃变温条件下的发芽率无显著差异(P>0.05)。层积的草玉梅种子在30/20℃、25/15℃、20/10℃的发芽率显著高于室温干燥贮藏同期处理(P<0.05)。室温干燥贮藏同期(11月-4月)的草玉梅种子在30/20℃、25/15℃、20/10℃、20/5℃和15/5℃变温条件均未能发芽。
2.2.3 对发芽指数的影响 高寒草甸土壤层积对草玉梅种子发芽指数的影响与其对发芽率的影响相似(表4)。践踏层积的草玉梅种子在25/15℃和20/10℃变温条件下的发芽指数分别显著高于层积不践踏处理(P<0.05),但两处理在30/20℃变温条件下的发芽指数无显著差异(P>0.05)。层积的草玉梅种子在30/20℃、25/15℃、20/10℃的发芽指数显著高于室温干燥贮藏同期处理(P<0.05)。
2.2.4 践踏层积的草玉梅种子在高寒草甸的发芽情况 层积于天祝高寒草甸放牧地的草玉梅种子,截至8月底,有40.2%的发芽。未发芽的草玉梅种子在培养箱30/20℃到20/5℃的温度范围均能发芽,但发芽率都较低,在30/20℃最高,仅为15.3%,绝大多数种子处于休眠(表5)。8月底取回的未发芽种子在培养箱内30/20℃的初次发芽天数为第2.0天,显著早于层积于1 0月到4月的种子(表4、表5);但在20/10℃变温条件下为第11.0天,显著迟于层积于10月到4月的种子(表4、表5)。8月底取回的未发芽种子发芽指数和发芽率的结果相似。
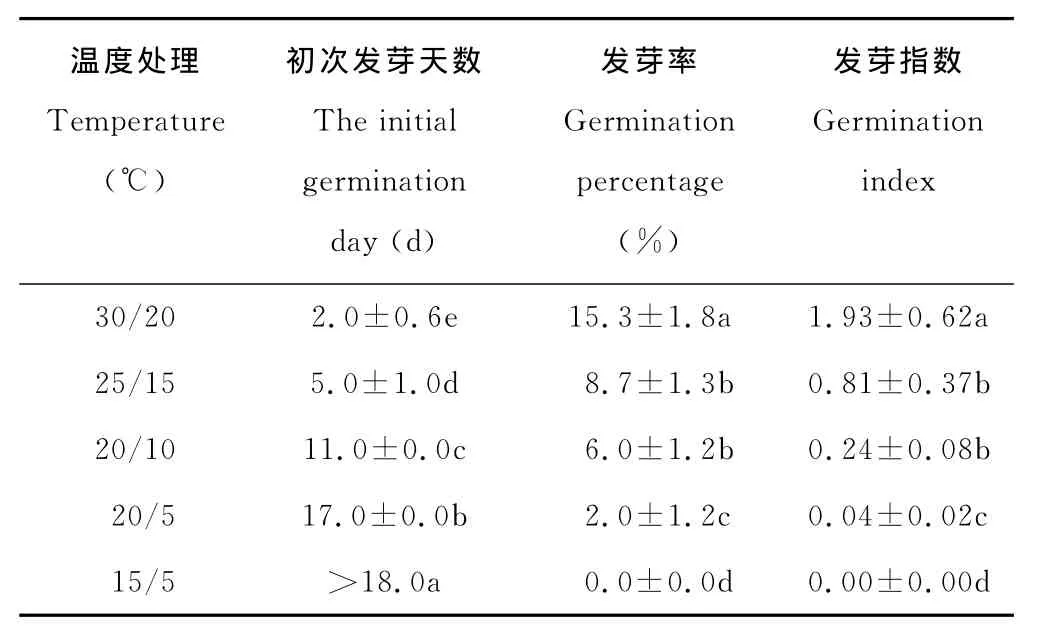
表5 层积(11-8月)于高寒草甸未发芽草玉梅种子在不同变温条件下的初次发芽天数、发芽率和发芽指数Table 5 The initial germination day,germination percentage and germination index of un-geminated A.rivularis seeds under different alternative temperature conditions after stratification from November to August at the alpine meadow
3 讨论
在多变的环境中,植物种群的成功建立取决于种子在适宜的条件下萌发和幼苗形态建成[11]。种子在逆境中休眠,这是植物适应不利环境和保护物种延续的一种生态策略[33]。许多植物的种子在成熟时是休眠的,只有当环境条件与种子的萌发需求完全或部分重叠时,种子才能萌发[32]。室温干燥贮藏能有效降低某些植物种子的休眠,如骆驼蒿种子经室温干燥贮藏6个月,其发芽率由贮藏前的8%提高到98%[11];少花蒺藜草(Cenchrus pauciflorus)种子经室温干燥贮藏6个月后,发芽率由0%提高到81%[34];盐生肉质沙漠植物Salsolaimbricata和Haloxylonsalicornicum种子经室温干燥贮藏3个月后发芽率显著提高[35]。本研究首次进行了室温干燥贮藏和天然高寒草甸土壤中层积处理对草玉梅种子萌发的影响,研究结果显示,随着室温贮藏天数的增加,去掉果皮草玉梅种子的初始发芽天数呈不同程度的减小,发芽率和发芽指数呈不同程度的增大,且减小或增大的趋势与是否去掉果皮和是否光照有关。室温干燥贮藏处理40~180d、去掉果皮后光照和自然光处理的草玉梅种子发芽率为53.0%~72.7%;贮藏40,80,150和180d未去掉果皮的草玉梅种子在光照下的发芽率仍然较低,分别为8.7%,11.0%,14.7和39.3%,在黑暗条件下更低(分别为0.0%,0.0%,0.0%和0.7%)。这说明草玉梅种子萌发为需光类型,室温干燥贮藏能有效降低草玉梅种子的休眠,但其发芽需要去掉果皮。未去掉果皮的草玉梅种子发芽率较低,尤其是在黑暗条件下的发芽率更低,达不到农业生产对种子发芽率的要求[36]。
层积处理能使种子中的抑制物质含量下降、种皮软化、提高透性、种胚发育后熟,为种子发芽提供了有利条件,已在打破多种植物种子休眠上取得较好效果[3,11-12,15]。本研究结果显示,层积践踏、层积不践踏处理的草玉梅种子在30/20℃、25/15℃和20/10℃下有部分种子发芽,其发芽率均显著高于室温干燥贮藏的种子。由于草玉梅种子(瘦果)的休眠为果皮遮光引起种子休眠的物理休眠和胚休眠类型组成的综合休眠类型[28]。这说明在层积和践踏过程中,草玉梅果皮对光线到达胚部的遮光作用减弱,草玉梅种子(瘦果)萌发对光照的依赖性降低,其种子的休眠在逐步释放。
许多植物的种子都具有休眠特性,野生植物尤其是逆境下生活的野生植物和某些杂草种子的休眠率更高[3]。世界各植被区的7351种植物的种子中,休眠物种占70.1%,不休眠物种仅为29.9%[1]。休眠可防止种子在适宜萌发但不利于幼苗建植的环境条件下萌发,土壤中始终保留着一部分休眠的活种子,如遇到突发性的自然灾害后可继续产生新的植株,从而确保种群在风险环境中的延续[1,3,33,37]。在环境相对恶劣区域,种子长时间处于休眠状态直至环境适宜萌发与幼苗形态建成,被视为是物种重要的适应环境策略[5,38]。本研究结果显示,当年成熟的草玉梅种子层积于天祝高寒草甸放牧地至次年8月底有40.2%的发芽,未发芽的草玉梅种子在30/20℃到20/5℃的温度范围均能发芽,但发芽率都较低,在30/20℃最高,仅为15.3%,绝大多数的种子处于休眠。草食动物放牧过程中,动物的践踏、采食和风吹雨淋使得天然草地植物种子的果皮逐渐分解或部分果皮脱落、抑制物质含量下降、种皮软化、透性提高[25,39]。在青藏高原高寒草甸,草玉梅瘦果秋季成熟后脱落母株掉落在土壤表层,由于受制于休眠和温度,成熟的草玉梅种子在当年不发芽,其休眠性为其避开冬季寒冷条件保持种子活力创造条件。这些被埋进土壤中含有一定量水分的草玉梅瘦果或真种子在天然高寒草甸土壤中经历层积践踏和不践踏处理,等到春、夏季气温上升后,部分草玉梅种子发芽生长,部分种子因果皮的完整性较好,继续休眠,直到果皮脱落或透光性变得满足其种子发芽条件,遇到适宜的水分和温度后陆续发芽,进而生长、繁殖下一代。
种子室内干燥贮藏和天然草地层积代表了种子贮藏的两种主要方式,本研究进行了室内干燥贮藏和天然草地层积对草玉梅种子休眠和萌发的影响。室温干燥贮藏和高寒草甸土壤中层积可影响种子的生理后熟和形态后熟,以及影响草玉梅果皮的透光性[28],关于草玉梅种子的休眠类型和环境条件对草玉梅种子休眠的调控机理需要进一步研究。
[1]Baskin C C,Baskin J M.Seeds-ecology,Biogeography,and Evolution of Dormancy and Germination[M].San Diego:Academic Press,1998.
[2]Huang Z Y.Adaptation strategies of seed dormancy and germination ofPsammochloavillosa,a sand dune grass inhabiting Ordos Plateau,China.Acta Botanica Boreali-occidentalia Sinica,2003,23(7):72-77.
[3]Wang Y R,Yang L,Hu X W.Seed dormancy release and soil seed bank of three arid desert plants under burial conditions.Chinese Journal of Plant Ecology,2012,36(8):774-780.
[4]Thompson K,Ooi M K J.To germinate or not to germinate:more than just a question of dormancy.Seed Science Research,2010,20:209-211.
[5]Donohue K,Dorn L,Griffith C,etal.The evolutionary ecology of seed germination ofArabidopsisthaliana:Variable natural selection on germination timing.Evolution,2005,59:758-770.
[6]Zhang J N,Liu K.Mechanism for plants detecting the optimum time and place to germinate.Acta Prataculturae Sinica,2014,23(1):328-338.
[7]Wang J H,Chen W,Ma R J.Effects of storage condition on the seed germination of four Solanaceae species in Hexi Corridor,China.Chinese Journal of Ecology,2013,32(7):1807-1812.
[8]Song Y G,Li L,Zeng X H,etal.Response of the germination on dimorphic seeds ofSuaedaaralocaspicato salt stress.Acta Prataculturae Sinica,2014,23(1):192-198.
[9]Wang L D,Yao T,He F L,etal.Natural vegetation restoration and change of soil enzyme activity on secondary grassland of abandoned land area in the downstream of Shiyang River.Acta Prataculturae Sinica,2014,23(4):253-261.
[10]Li X M,Zhao X Y,Yu R D.Effects of seed maturation time and dry storage on germination of twoAtraphaxisspecies.Acta Ecologica Sinica,2010,30(14):3727-3732.
[11]Zhen X F,Chen W,Wang J H,etal.Effect of cold stratification and dry storage at room temperature on seed germination of four herbaceous species of Zygophyllaceae.Agricultural Research in the Arid Areas,2011,29(4):53-59.
[12]Luo X,Pan Y Z,Yang X J,etal.Effect of cold stratification on dormancy and germination of fourCarexspecies seeds.Acta Prataculturae Sinica,2010,19(3):117-123.
[13]Ma X J,Wei C H,Zhang Y R,etal.Effects of cold-wet stratification,light and temperature on the seed germination of alpine meadow plants.Journal of Anhui Agricultural Sciences,2008,36(6):2221-2223.
[14]Probert R J.The role of temperature in the regulation of seed dormancy and germination.In:Fenner M.Seed:The Ecology of Regeneration in Plant Communities(2ndedn)[M].Wallingford:CABI Publishing,2000:261-291.
[15]Yu X J,Wang F,Long R J.Advance of the methods to relieve seed dormancy.Seed,2005,24(7):46-49.
[16]Wang J H,Chen W,Zhang Y,etal.Effects of storage condition on seed germination of four xeromorphic shrub species from Hexi Corridor.Chinese Journal of Ecology,2011,30(3):477-482.
[17]Zhao Y Z.Distribution pattern and migration route ofAnemonerivularisandA.rivalarisvar.flore-minore.Acta Scientiarum Naturalium Universitatis Neimongol,2002,33(4):443-445.
[18]Liu S W.Flora of Qinghai(First Volumes)[M].Xining:Qinghai People’s Publishing Press,1997.
[19]Cao P,Wu F E,Ding L S.Advances in the studies on the chemical constituents and biologic activities forAnemonespecies.Natural Product Research and Development,2004,16(6):581-584.
[20]Zhang S,Wang H.Reproductive biology and ecological adaptation ofAnemonerivularisin Himalaya,China[C].Kunming:Proceeding of National Systematic and Evolutionary Botany in 2011and the Tenth Session of the Youth Academic Seminar,2011.
[21]Zhang H Y,Zhang Z Y.Traditional Chinese Medicine Resources[M].Beijing:Science Press,1994.
[22]Liao X,Li B G,Wang M K,etal.The chemical constituents fromAnemonerivularis.Chemical Research in Chinese Universities,2001,22(8):1338-1441.
[23]Zhang L F,Wei B,Ge Q Z,etal.Relationship between productivity and plant functional traits along successive recovery stages in an alpine meadow.Acta Prataculturae Sinica,2012,21(6):235-241.
[24]Shi F X,Zhao C Z,Ren H,etal.Plant functional types and their impact environmental factors in a naturally recovered poplar-birch woodland on the north slope of Qilian Mountains,China.Chinese Journal of Applied and Environmental Biology,2012,18(4):546-552.
[25]Yu X J,Xu C L,Wang F,etal.Recovery and germinability of seeds ingested by yaks and Tibetan sheep could have important effects on the population dynamics of alpine meadow plants on the Qinghai-Tibetan Plateau.The Rangeland Journal,2012,34:249-255.
[26]Duan C H,Yu X J,Xu C L,etal.Effects of soaking with Tibetan sheep rumen fluid on the seed germination of 11plant species on alpine meadow of Qinghai-Tibetan Plateau,China.Chinese Journal of Ecology,2013,32(6):1483-1489.
[27]Duan C H.Effect of Different Treatments on Seed Germination Characteristics of 12Alpine Meadow Species[D].Lanzhou:Gansu Agricultural University,2013.
[28]Yu X J,Xu C L,Wang F,etal.A study on the causes ofAnemonerivularisdormancy and the methods for breaking dormancy.Chinese Journal of Ecology,2014,33(1):65-70.
[29]Yu X J,Pu X P,Huang S J,etal.Effects of ants(Tetramoriumsp.)on eastern Qilian Mountains alpine grassland ecosystem.Acta Prataculturae Sinica,2010,19(2):140-145.
[30]Ren J Z.Research on the Method of Pratacultural Science[M].Beijing:China Agriculture Press,1998.
[31]Ministry of Agriculture of the People’s Republic of China.NY/T 635-2002,Calculation of Stocking Rate in Natural Rangeland[S].Beijing:Chinese Standards Press,2002.
[32]Wang J H,Ma R J,Chen W.Effects of cold stratification and dry storage at room temperature on seed germination of eight desert species from the Hexi Corridor of China.Chinese Journal of Plant Ecology,2012,36(8):791-801.
[33]Yu X J.Seed dormancy of forage plants.In:Shi S L.Seed Science of Forage Plants[M].Beijing:Science Press,2011.
[34]Zhou L Y,Li J H,Ma F,etal.Study on seed germination characteristics ofCenchruspauciflorusBenth.Journal of Inner Mongolia University for Nationalities(Natural Sciences),2013,28(2):203-205.
[35]Ali E K.Effects of seed storage on germination of two succulent desert halophytes with little dormancy and transient seed bank.Acta Ecologica Sinica,2013,33(6):338-343.
[36]Han J G.Forage Seed Science[M].Beijing:China Agricultural University Press,2000.
[37]Yang J N,Wang R Y.Effects of drought stress simulated by PEG on seed germination of four desert plant species.Acta Prataculturae Sinica,2012,21(6):23-29.
[38]Thompson K,Ceriani R M,Bakker J P,etal.Are seed dormancy and persistence in soil related.Seed Science Research,2003,13:97-100.
[39]Hou F J,Chang S H,Yu Y W,etal.A review on trampling by grazed livestock.Acta Ecologica Sinica,2004,24(4):784-789.
参考文献:
[2]黄振英.鄂尔多斯高原固沙禾草沙鞭种子休眠和萌发与环境的关系.西北植物学报,2003,23(7):72-77.
[3]王彦荣,杨磊,胡小文.埋藏条件下3种干旱荒漠植物的种子休眠释放和土壤种子库.植物生态学报,2012,36(8):774-780.
[6]张佳宁,刘坤.植物调节萌发时间和萌发地点的机制.草业学报,2014,23(1):328-338.
[7]王桔红,陈文,马瑞君.不同贮藏方式对河西走廊四种茄科植物种子萌发的影响.生态学杂志,2013,32(7):1807-1812.
[8]宋以刚,李利,曾歆花,等.异子蓬二型性种子萌发对盐胁迫的响应.草业学报,2014,23(1):192-198.
[9]王理德,姚拓,何芳兰,等.石羊河下游退耕区次生草地自然恢复过程及土壤酶活性的变化.草业学报,2014,23(4):253-261.
[10]李晓梅,赵晓英,于瑞德.成熟时间和干贮藏对两种木蓼种子萌发的影响.生态学报,2010,30(14):3727-3732.
[11]郑秀芳,陈文,王桔红,等.低温层积和室温干燥贮藏对4种蒺藜科草本植物种子萌发的影响.干旱地区农业研究,2011,29(4):53-59.
[12]罗弦,潘远智,杨学军,等.低温层积处理对4种苔草种子休眠与萌发的影响.草业学报,2010,19(3):117-123.
[13]马晓娟,魏茶花,张摇荣,等.冷湿层化光照和温度对高寒草甸植物种子萌发的影响.安徽农业科学,2008,36(6):2221-2223.
[15]鱼小军,王芳,龙瑞军.破除种子休眠方法研究进展.种子,2005,24(7):46-49.
[16]王桔红,陈文,张勇,等.贮藏条件对河西走廊四种旱生灌木种子萌发的影响.生态学杂志,2011,30(3):477-482.
[17]赵一之.草玉梅与小花草玉梅的分布式样和迁移路线.内蒙古大学学报(自然科学版),2002,33(4):443-445.
[18]刘尚武.青海植物志(第1卷)[M].西宁:青海人民出版社,1997.
[19]曹沛,吴凤锷,丁立生.银莲花属植物的化学及药理研究概况.天然产物研究与开发,2004,16(6):581-584.
[20]张舒,王红.中国—喜马拉雅草玉梅繁殖生物学及生态适应研究[C].昆明:2011年全国系统与进化植物学暨第十届青年学术研讨会论文集,2011.
[21]张惠源,张志英.中国中药资源志要[M].北京:科学出版社,1994.
[22]廖循,李伯刚,王明奎,等.草玉梅中的化学成分.高等学校化学学报,2001,22(8):1338-1441.
[23]张灵菲,魏斌,葛庆征,等.高寒草甸恢复演替过程中生产力与植物功能特征关系的研究.草业学报,2012,21(6):235-241.
[24]石福习,赵成章,任珩,等.祁连山北坡自然恢复杨桦林地植物功能型组成及其影响因素.应用与环境生物学报,2012,18(4):546-552.
[26]段春华,鱼小军,徐长林,等.藏羊瘤胃内容物浸泡对11种高寒草甸植物种子萌发的影响.生态学杂志,2013,32(6):1483-1489.
[27]段春华.不同处理对12种高寒植物种子萌发特性的影响[D].兰州:甘肃农业大学,2013.
[28]鱼小军,徐长林,王芳,等.草玉梅种子休眠机制的研究.生态学杂志,2014,33(1):65-70.
[29]鱼小军,蒲小鹏,黄世杰,等.蚂蚁对东祁连山高寒草地生态系统的影响.草业学报,2010,19(2):140-145.
[30]任继周.草业科学研究方法[M].北京:中国农业出版社,1998.
[31]中华人民共和国农业部.NY/T 635-2002,天然草地合理载畜量的计算[S].北京:中国标准出版社,2002.
[32]王桔红,马瑞君,陈文.冷层积和室温干燥贮藏对河西走廊8种荒漠植物种子萌发的影响.植物生态学报,2012,36(8):791-801.
[33]鱼小军.草类植物种子的休眠.见:师尚礼.草类植物种子学[M].北京:科学出版社,2011.
[34]周立业,李建华,马菲,等.少花蒺藜草种子发芽特性研究.内蒙古民族大学学报(自然科学版),2013,28(2):203-205.
[36]韩建国.牧草种子学[M].北京:中国农业大学出版社,2000.
[37]杨景宁,王彦荣.PEG模拟干旱胁迫对四种荒漠植物种子萌发的影响.草业学报,2012,21(6):23-29.
[39]侯扶江,常生华,于应文,等.放牧家畜的践踏作用研究评述.生态学报,2004,24(4):784-789.
