高寒草甸土壤异养呼吸对气候变化和氮沉降响应的模拟
2015-01-02李东罗旭鹏曹广民吴琴卓玛措李惠梅杨永梅庞炳坤
李东,罗旭鹏,曹广民,吴琴,卓玛措,李惠梅,杨永梅,庞炳坤
(1.青海民族大学青藏高原生态环境研究所,青海 西宁810007;2.南京农业大学资源与环境科学学院,江苏 南京210095;3.西宁市林业科学研究所,青海 西宁810003;4.中国科学院西北高原生物研究所,青海 西宁810006)
气候变暖和大气氮沉降增加作为全球变化中最突出的环境问题而备受关注。IPCC-TAR4指出[1],过去百年(1906-2005年)全球地表温度上升了0.56~0.92℃。温室气体若以当前的或高于当前的速率排放,21世纪全球地表温度将上升1.1~6.4℃。同时,人类活动(化肥的使用、矿物质燃料燃烧和工业排放等)向大气系统排放了大量的氮化物。Galloway等[2]估计,1860-2000年人类活动带来的活性氮生产由15Tg N/a提高到165Tg N/a,增幅高达11倍,约为全球氮素临界负荷(100Tg N/a)的1.6倍[3]。事实证明,这些氮进入大气层后通过大气转化与大气环流60%~80%的氮素又沉降到陆地和海洋生态系统[4-5],对生态系统,尤其是温带生态系统的特征和过程产生影响[6],而北半球高纬度、高海拔地区生态系统对其的响应可能更加敏感和迅速[7]。
青藏高原是地球陆地生态系统的重要组成部分,作为欧亚大陆最高最大的地貌单元,不仅对全球气候变化十分敏感,而且在亚洲气候乃至全球气候变化中扮演重要角色[8]。地面气象观测资料分析结果显示,近47年(1961-2007年)高原气温呈显著暖化趋势,年平均气温以0.37℃/10a的速率上升[9],增温明显高于中国其他地区[10]。同时,高原东部地区大气氮沉降也十分明显,并呈逐年增加趋势,变化范围在8.7~13.8kg N/(hm2·a)之间[11],平均值11.25kg N/(hm2·a)明显高于亚洲7kg N/(hm2·a)[12]及全球5kg N/(hm2·a)[13]氮沉降平均值。受高原气候暖化和氮沉降增加的影响,长期受低温和土壤有效氮限制的高海拔生态系统碳、氮循环过程将不可避免地发生变化,并通过反馈作用对高原周边地区、中国乃至全球尺度的气候系统产生影响。
高寒草甸(面积约51.7×104km2[14])是青藏高原大气与地面之间生物地球化学循环的重要构成部分,在区域碳平衡中起着极为重要的作用。目前,针对高寒草甸碳循环的研究主要集中在净初级生产力[15-18]和土壤有机碳方面[19-24]。土壤呼吸过程作为高寒草甸碳循环研究中的重要环节,虽然也有研究和报道[25-32]。但这些研究大多数仅涉及土壤呼吸的短期流量,季节动态及其影响因素或根系呼吸贡献量等。而从碳平衡的角度出发,系统估算土壤呼吸的年度总量,分析长时间尺度上土壤呼吸对气候变化响应的研究较少,对于异养呼吸的报道也较为鲜见。中国科学院海北高寒草甸生态系统定位研究站建站于1976年,至今已对研究区气候、土壤、植被及主要温室气体通量等进行了长期监测和研究,积累了一些数据。这为模型研究高寒草甸生态系统土壤碳、氮循环过程及其对气候变化的响应提供了基础数据。本研究在分析总结现有研究成果的基础上,对生物地球化学模型CENTURY进行了验证,并进一步利用该模型研究了1960-2005年海北站地区高寒草甸土壤异养呼吸CO2通量年际间变化,并着重分析了研究区主要气候因子(气温和降水量)的波动变化和氮沉降倍增对异养呼吸过程的影响。旨在为全球变化背景下,定量研究青藏高原高寒草甸生态系统源汇特征及其变化提供必要的参考。
1 材料与方法
1.1 研究区概况
研究地位于中国科学院海北高寒草甸生态系统定位站(海北站,图1)。该站地处青藏高原东北隅的青海省海北藏族自治州门源回族自治县境内,祁连山北支冷龙岭东段南麓坡地的大通河河谷西段。地理位置为37°29′-37°45′N,101°12′-101°23′E,山地平均海拔4000m,站内以滩地和丘陵低地为主,平均海拔3200m 左右。属典型的高原大陆性气候特征,冷季漫长而寒冷,暖季短暂而湿润,年平均气温-1.7℃,年降水量618mm,且主要集中于6-8月,占全年降水量的80%左右。植被以莎草科嵩草属(Kobresia)植物为主建群种,如矮嵩草(Kobresiahumilis)、小嵩草(K.pygmaea)、线叶嵩草(K.capillifolia)等,伴有蓼科的珠芽蓼(Polygonum viviparum)以及龙胆属(Gentiana)、虎耳草属(Saxifraga)、银莲花属(Anemone)的高山植物。土壤为草毡寒冻雏形土,草毡表层发育良好,没有发生剥蚀脱落,pH值7.0~7.5。
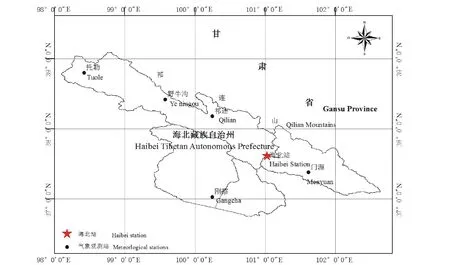
图1 中国科学院海北高寒草甸生态系统定位站地理位置Fig.1 The geographical location of the Research Station of Alpine Meadow Ecosystem,Chinese Academy of Sciences,Qinghai Province
1.2 研究方法
1.2.1 CENTURY模型 CENTURY模型是美国科罗拉多州立大学的Parton等[33-34]建立,起初用于模拟草地生态系统的C、N、P、S等元素的长期演变过程。由于模型将影响土壤有机质分解的4个重要变量(温度、降雨量、土壤质地和植物木质素含量)作为确定一个地点的特征值。同时,在模型改进中增加了栽培、施肥、灌溉、火烧和放牧等人为措施的影响,使CENTURY模型的应用范围从草地生态系统逐步扩大到农田、森林和热带(或亚热带)稀树草原(savanna)生态系统中,并已成为以模拟土壤有机质积累分解为主的比较有影响的模型之一。
模型主要包括土壤有机质、植物产量、土壤水分和温度3个子模型。其中,土壤有机质(SOM)模型采用ROTHC模型[35]的基本思想,将土壤有机质分为活性、缓性和惰性3个组分库。其中活性土壤有机质(active SOM)包括活的微生物及其代谢产物,大小约是活的微生物生物量的2~3倍,周转时间1~5年;缓性土壤有机碳(slow SOM)包括难分解的土壤有机物质和土壤固定的微生物产物,周转时间20~40年。惰性土壤有机碳(passive SOM)是土壤中受物理或化学保护的,极难分解的部分,周转时间长达200~1500年,甚至更长。输入土壤的植物残体库则分为两个库,即不易分解的周转时间在1~5年的结构库(structural pool)和易分解的周转时间在0.1~1年的代谢库(metabolic pool)。由于植物残体地上部分和地下部分的木质素含量相差较大,使得结构库和代谢库具有各自的地上和地下分解速率。土壤有机质各组分库最大分解速率为常数,但受土壤温度、湿度、木质素含量、土壤粘粒含量等的影响。
模型气候参数主要包括月平均最高、最低气温(℃)和月平均降水量(cm)。其他参数包括研究区经纬度、土壤质地[砂粒、粉粒和粘粒含量(%)]、土壤容重(g/cm3)、pH值、凋落物碳氮比、木质素含量(%)、大气干湿沉降和非生物固氮量(g N/m2)及各种草地管理措施等。目前模型有DOS提示符模式和窗口操作模式2个版本,本研究使用的CENTURY模型为DOS 4.0,该版本的详细介绍可参考模型相关网站(http://www.nrel.colostate.edu/projects/century/)。
1.2.2 模型参数化 模型本地化校验与应用的关键在于参数获取。本研究CENTURY模型所需的气象参数取自青海省海北藏族自治州门源县气象观测站(站点编码52765)1960-2005年实测数据(表1)。在输入模型之前,按CENTURY模型所需的格式建立气象资料数据库文件(*.wth),缺失数据的年份用-9.99补齐。土壤和植被参数取自海北站1998年监测数据。植物地上/地下木质素含量因缺乏连续观测数据,不能建立木质素与年降水量的函数关系,其参数值由模型程序根据实际年降水量计算获得。大气干湿沉降和非生物氮固定量[36]每年以固定值输入模型。植被生长参数根据地区牧草生长状况,以5月份牧草返青,8月份为最后生长期,9月份进入枯黄期输入模型。主要气候因子及模型初始化参数如表1和表2所示。
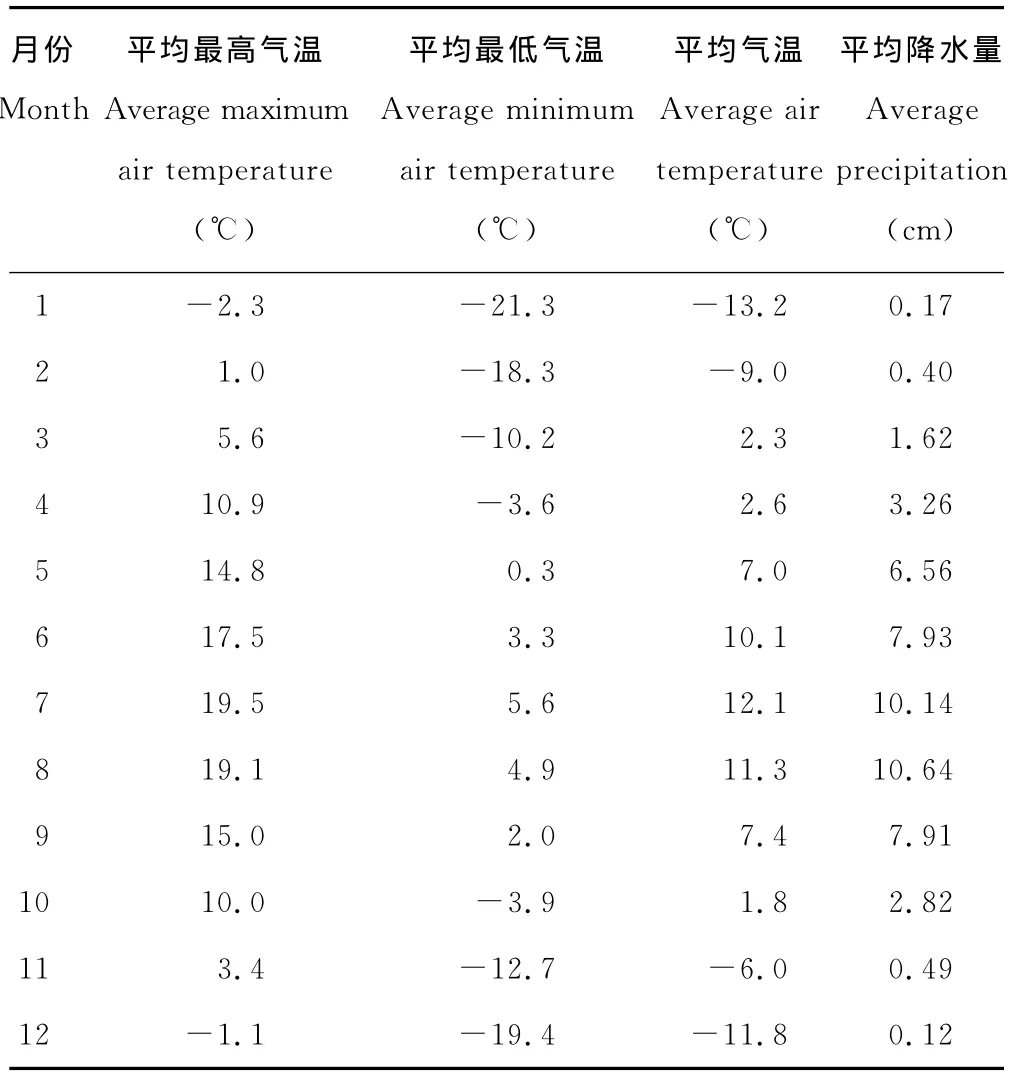
表1 1960-2005年高寒草甸区主要气候因子Table 1 Main meteorological factors of alpine meadow site(1960-2005)
1.2.3 模型验证 模拟结果的可行度只有与实测数据进行比较分析才能确定。尽管CENTURY模型对青藏高原高寒草甸土壤有机碳(SOC)和净初级生产力(NPP)的估算已有研究和报道[22,24,37],模拟结果经野外观测数据验证具有较高可信度。但如果将该模型应用到土壤异养呼吸时,其适用性有待进一步验证。本研究选取的是中国科学院海北高寒草甸生态系统长期定位站土壤异养呼吸CO2通量实测数据(2003-2005年),有关碳通量的研究和报道,可参阅本课题组发表的相关论文[25-28,30,32]。
模型运行时,输入上述参数以1960-2005年气象数据运行模型5000年,建立CENTURY模型参数在高寒草甸生态系统中的平衡状态。模型运行稳定后,以2003-2005年逐月平均最高、最低气温(℃)和月平均降水量(mm)为驱动变量,月为时间步长运行模型,得出同期高寒草甸土壤异养呼吸逐月CO2通量(0~20cm)模拟结果。输出结果采用平均绝对误差(the mean absolute deviation,Dabs)和线性回归分析(the slope coefficient)2种方法进行综合评价。具体公式如下:
Ⅰ)平均绝对误差(Dabs)
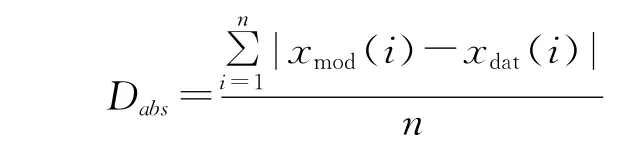
式中,变量xmod和xdat分别为ti(i=1,2,3……n,n=30,31)时的模拟值和观测值。
Ⅱ)线性回归方程
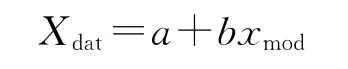
式中,Xdat为观测值,xmod为模拟值,b为斜率,a为截距。模型模拟最理想的结果应该是a=0,b=1。因此,线性回归方程中b与1的接近程度是反映CENTURY模型模拟效果的一个重要指标。
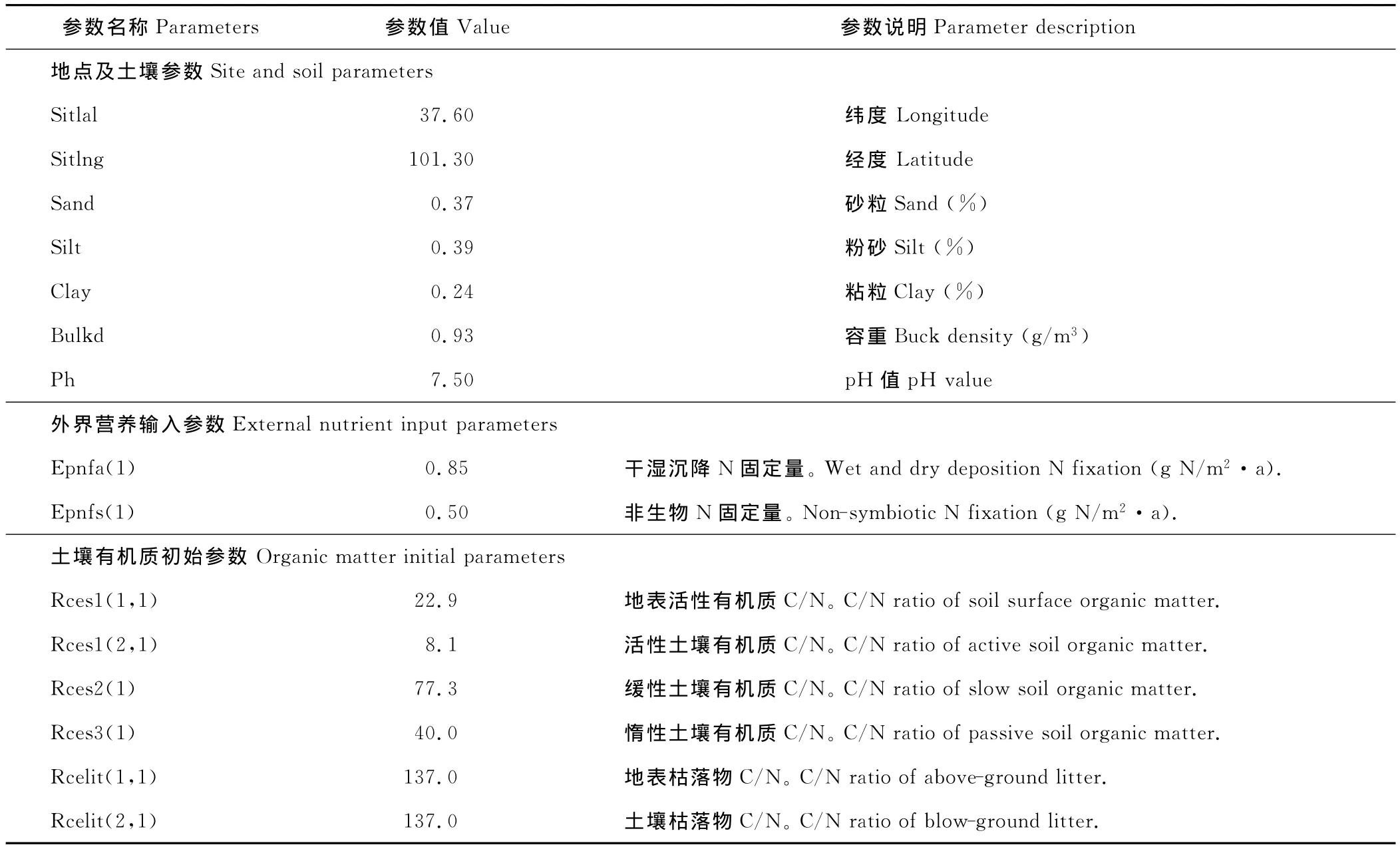
表2 CENTURY模型初始化参数Table 2 Initial parameters of CENTURY model
1.2.4 情景模拟 长期连续的地面气象观测数据能够反映区域气候的地面特征量以及气候本身的变化规律,在影响研究中得到了广泛的应用。气候变化响应模拟时,利用青海省海北藏族自治州门源县气象站历年气侯资料记载的月平均最高、最低气温(℃)和月平均降水量(mm)驱动模型,得出1960-2005年高寒草甸土壤异养呼吸逐年CO2通量模拟结果。提取同期(1960-2005年)地面观测的年平均气温(℃)和年降水量(mm),分析气候因子的波动变化对土壤异养呼吸过程的影响。氮沉降响应模拟时,利用海北站大气干湿氮沉降报道结果7.2~10.0kg N/(hm2·a)[36],平均值8.5kg N/(hm2·a)。分别设置对照(CK,control:8.5kg N/hm2·a)、中氮(MN,medium N:17.0kg N/hm2·a)及高氮(HN,high N:34.0kg N/hm2·a)3种情景。模拟运算时,保持模型基本参数不变,大气干湿氮沉降参数值分别以固定值Epnfa(1)=0.85g N/(m2·a)、Epnfa(1)=1.7g N/(m2·a)和Epnfa(1)=3.4g N/(m2·a)输入并运行模型,得出3种情景下1960-2005年高寒草甸土壤异养呼吸逐年CO2通量模拟结果。模拟值在进行显著性检验后,分析氮沉降倍增对土壤异养呼吸过程的影响。
1.2.5 数据统计与分析 采用 Microsoft Excel 2003软件完成数据处理,相关分析由SPSS 13.0完成。
2 结果与分析
2.1 模型验证
选择中国科学院海北高寒草甸生态系统定位研究站(海北站)马场风匣口南滩(37°29′-37°45′N,101°12′-101°33′E)和干柴滩(37°29′-37°45′N,37°29′-37°45′E)2个观测点数据对CENTURY模型进行验证。图2a,b分别为2003-2005年风匣口、干柴滩观测场定点观测的高寒草甸土壤异养呼吸CO2通量季节变化与模拟结果的比较。结果显示,CENTURY模拟结果与试验点观测结果相吻合,观测值与模拟值的线性回归方程分别为y=0.7776x+23.796(R2=0.6885,n=31)和y=0.9487x-8.6994(R2=0.6062,n=30)。模拟值相对观测值的平均绝对误差(Dabs)分别为18.10和16.97g C/m2。表明,CENTURY模型较好地反映了高寒草甸土壤异养呼吸的季节变化动态,可以模拟不同情景下土壤异养呼吸CO2通量的长期动态变化及其对气候变化的响应。
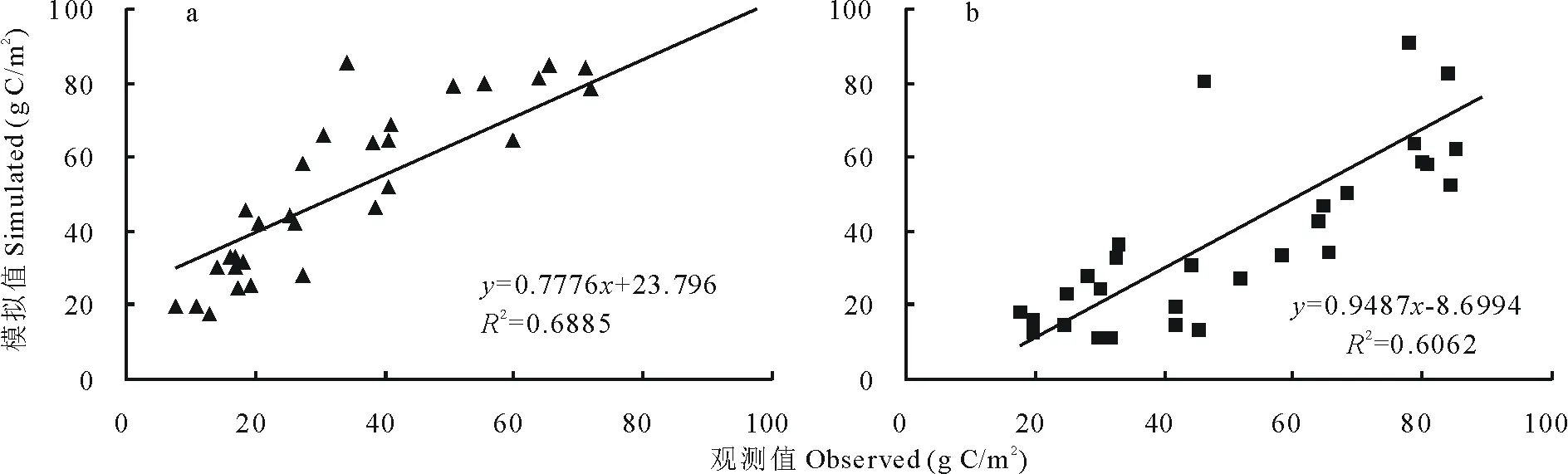
图2 风匣口(a)、干柴滩(b)观测点土壤异养呼吸季节动态模拟结果的检验Fig.2 Test of CENTURY model for simulations the seasonal dynamic of soil heterotrophic respiration at Fengxiakou(a)and Ganchaitan(b)experiment sites
2.2 土壤异养呼吸对气候变化的响应
由图3可以看出,1960-2005年高寒草甸区年平均气温趋于暖化,平均线性增温率为0.35℃/10a,尤其是80年代后期增温趋势更为明显,平均线性增温率达0.45℃/10a。降水量变化呈振幅较为稳定的波动变化,最小降水量出现在1962年(380.8mm),最大降水量出现在1989年(730.0mm),年平均降水量为522.5mm。同期,CENTURY模拟的高寒草甸0~20cm土壤异养呼吸CO2通量呈波动性缓慢上升趋势,变化范围在479.22~624.89g C/(m2·a)之间,平均值为(539.56±34.32)g C/(m2·a),通量增加率为16.5g C/(m2·10a)。
根据通量的上述变化情况,我们将1960-2005年高寒草甸土壤异养呼吸CO2通量模拟结果分3个区间分别与46年平均值进行比较。表3显示,20世纪60年代为通量小幅上升期,平均值为(514.32±20.09)g C/(m2·a),但10年间(1960-1969年)除1968,1969年通量值略高于46年平均值,其他8个年份均低于46年平均值。70-80年代为波动变化期,平均值为(529.83±19.37)g C/(m2·a),20年间(1970-1989年)有7个年份的通量值接近或明显高于46年平均值。1990-2005年为上升高峰期,平均值为(567.49±37.85)g C/(m2·a),16年间除1990和1991年外,其他14个年份的通量值均接近或明显高于46年平均值。对模拟结果与主要气候因子(气温和降水量)进行的相关分析表明,高寒草甸土壤异养呼吸CO2通量与年平均气温(℃)具有显著正相关(r=0.7,P<0.05),降水量(mm)的变化对其影响不显著。
2.3 土壤异养呼吸对氮沉降倍增的响应
氮沉降输入增加显著地促进了高寒草甸土壤异养呼吸CO2释放(图4)。1960-2005年对照(CK)、中氮(MN)和高氮(HN)情景下,CENTURY模拟的高寒草甸0~20cm土壤异养呼吸CO2通量变化范围分别为479.22~624.89g C/(m2·a),582.16~828.54g C/(m2·a)和581.08~837.64g C/(m2·a),平均值为(539.56±34.32),(657.01±51.08)和(658.58±54.05)g C/(m2·a)。与对照(CK)相比,中氮(MN)、高氮(HN)通量分别增加了21.76%和22.06%。方差分析结果表明,中氮(MN)、高氮(HN)情景下通量模拟结果与对照(CK)差异极显著(P<0.01),但中氮(MN)与高氮(HN)之间差异不显著。表明,受土壤有效氮、磷限制的高寒草甸土壤呼吸过程对氮沉降响应敏感,大气氮沉降倍增将显著提高土壤异养呼吸CO2释放量,但随着氮沉降输入量的倍增其促进效应降低。
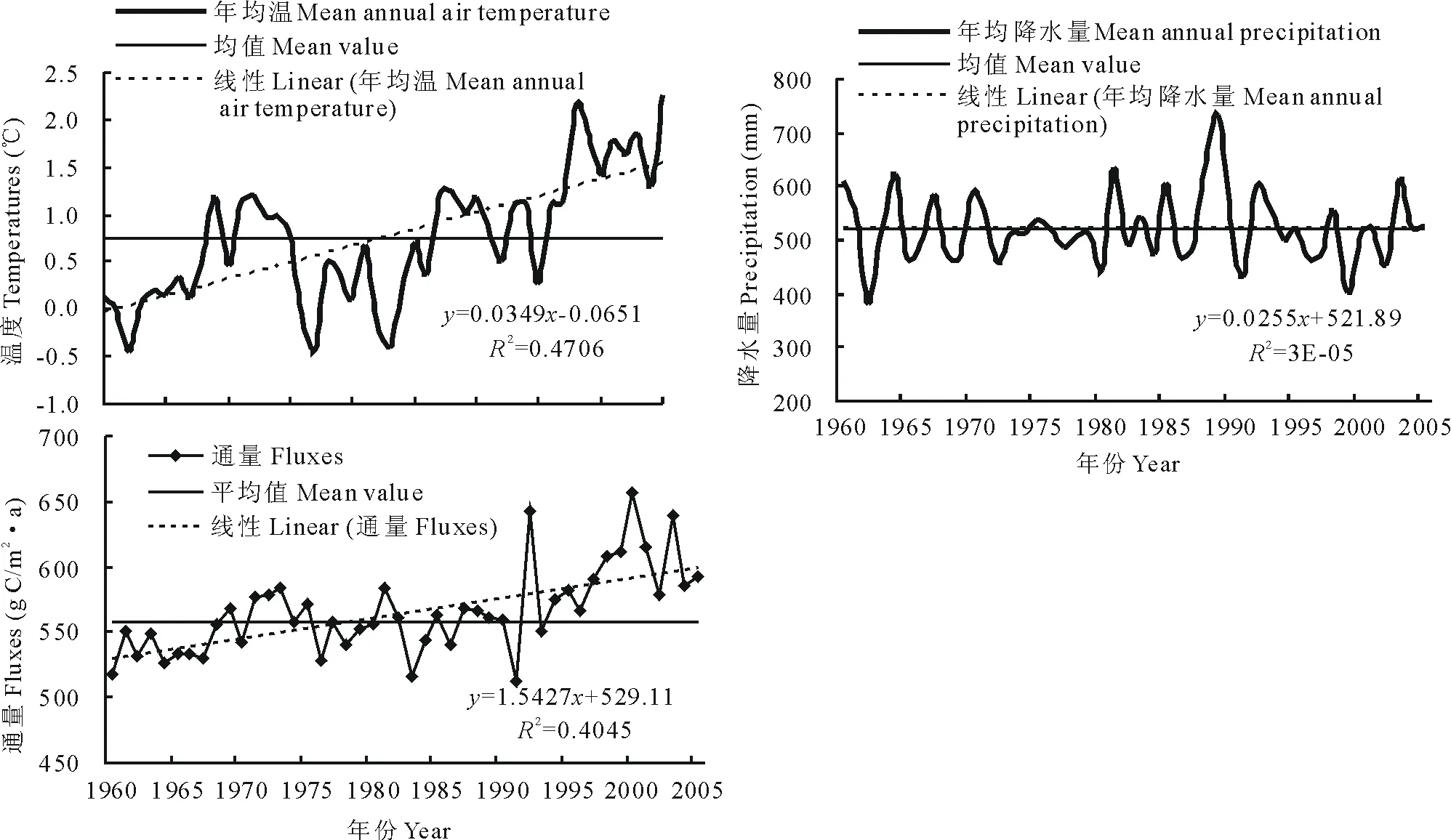
图3 1960-2005年研究区年平均气温、年平均降水量和年CO2通量动态变化Fig.3 Dynamic of mean annual air temperature,mean annual precipitation and annual CO2 fluxes during 1960to 2005

表3 1960-2005年高寒草甸土壤异养呼吸CO2通量变化趋势Table 3 Simulated trend of soil heterotrophic respiration fluxes in alpine meadow site(1960-2005)
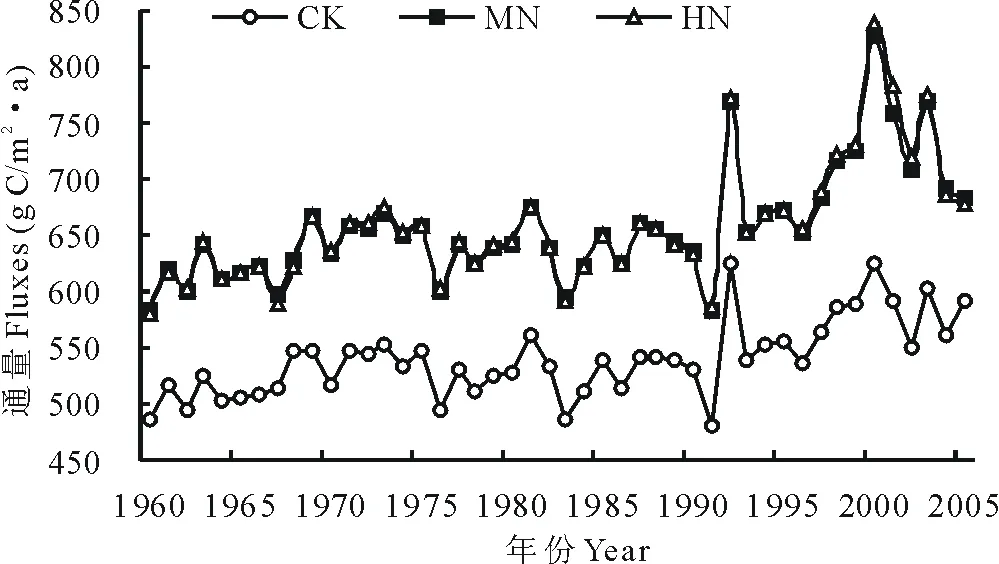
图4 3种氮沉降情景下CENTURY模拟的高寒草甸土壤异养呼吸CO2通量比较分析Fig.4 The analysis of annual CO2fluxes from soil microbial respiration in alpine meadow under different nitrogen deposition scenarios
3 讨论
土壤呼吸速率的变化受温度与水分共同调控[38-39]。温度升高一般会促进土壤 CO2的排放[40-41]。但由于温度和水分作用性质的不同,土壤呼吸过程对二者的响应程度在地区间存在一定的差异。通常受低温限制的高纬度或高海拔地区,气温升高可显著提高土壤中微生物或根系的代谢活性,从而导致土壤呼吸作用的增加[42],但随着增温时间的延长,土壤呼吸对温度变化表现出了一定的“适应现象”[43-46]。相对温度的影响,降水对土壤呼吸的影响相对较为复杂[47],降水强度、过程、历时长短都会对土壤呼吸产生影响,特别是在干旱和半干旱地区尤为如此。研究表明[48-49],降水后土壤呼吸明显增加,特别是在土壤处于长期干旱时的土壤呼吸再降水后的增加更为明显。但在内蒙古锡林河流域,4种草地群落土壤呼吸沿着降水梯度呈递减趋势[50],尤其是在强降雨后土壤温度变低或者强降雨使得土壤的空隙被雨水填满,减少了CO2排放的通路,从而使得土壤呼吸被显著抑制[28,51]。不难看出,目前针对土壤呼吸与温度、降水量变化的研究存在着诸多不确定性,而且在自然生态系统中,温度和水分往往交互作用影响土壤呼吸过程。因此,在较大的空间和时间尺度上综合考虑气温和降水量的变化,定量研究土壤呼吸各组分与气候因子之间的相关性更能反映出整个土壤呼吸过程对气候变化响应程度。土壤异养呼吸(HR)是土壤呼吸的重要组成部分,是土壤中微生物分解有机质释放CO2的过程,约占土壤呼吸CO2释放量的60%~90%[28,52-55],受温度、湿度、植被类型、土壤性质等多种因素影响,土壤异氧呼吸作用表现出了强烈的时空变异性。空间尺度上,谢薇等[56]通过文献调研研究了土壤异养呼吸与年降水量和年平均气温的关系后指出,中国陆地生态系统的土壤异养呼吸与年平均气温和年降水量均呈显著正相关,且与年降水量的相关性高于年平均气温,而Cao等[41]通过模型对中国区域的研究表明,土壤异氧呼吸的年际变化总体上同温度正相关,与降水量则相关性不显著。本研究在较长的时间尺度上(46a)研究了高寒草甸土壤异养呼吸与气温和降水量的关系,得出了与Cao等[41]相似的结论,土壤异养呼吸与年平均气温呈显著正相关关系(P<0.05),而与降水量相关性不显著。说明,受低温限制的高寒草甸生态系统土壤异养呼吸与气候变暖之间将构成正反馈环,高原气候系统的暖化将引起土壤异养呼吸作用的显著上升。
土壤中微生物分解有机质释放CO2的过程是一个受温度、土壤含水量、有机质含量以及氮可利用性等诸多因子影响的生物化学过程[57-59]。当大量的氮素持续进入生态系统后,可以通过改变土壤中微生物生物量、群落结构、组成以及微生物功能和活性,从而引起土壤异养呼吸作用的改变。Bowden等[60]对温带森林土壤进行培育实验后发现,施氮对土壤中的白腐菌产生了抑制作用,降低了酚氧化酶(一种木质素降解酶)的活性,从而引起异养呼吸的降低。同样,Olsson等[61]在瑞典北部的一个生长40年的挪威云杉(Piceaabies)林中进行施氮实验后指出,施氮降低了异养呼吸CO2释放,单位面积施氮样地异养呼吸减少了20%~30%。但这一结论并不是一个普遍现象。也有研究者认为,氮素增加可以引起植物地上部分对土壤有机碳输入的增加,最终表现为土壤中的有机质的增加,而这些增加的有机质为微生物提供了更多可利用的底物,从而有利于微生物的活动,促进土壤异养呼吸CO2释放[62]。如Emmett[63]研究后指出,在受氮素限制的森林生态系统中,增加氮输入量可增加土壤中微生物量,并增强其活性,加速土壤有机物分解,促进土壤CO2的排放。本研究得出了与Emmett[63]相似的结论,中氮、高氮情景下,CENTURY模型模拟的高寒草甸土壤异养呼吸CO2释放量相比对照均有显著提高。初步分析原因,这可能是由于高寒草甸植物生长和微生物活动长期受氮素的限制,氮素增加解除这一限制,增加了土壤中微生物分解底物(有机质)的输入量,同时微生物活性的增强又加速了底物的分解过程,进而引起异养呼吸作用的激增。但由于高寒草甸可能存在着一定的“氮饱和”现象,当氮素输入量超过土壤中生物(可能包括非生物)对氮的需求时,这种促进效应降低,这可能也是本研究中高氮(HN)与中氮(MN)处理间差异不显著的主要原因。
4 结论
CENTURY模型较好地反映了高寒草甸土壤异养呼吸季节变化动态,模拟与观测结果相吻合。2个实验点观测值与模拟值的线性回归方程分别为y=0.7776x+23.796(R2=0.6885,n=31)和y=0.9487x-8.6994(R2=0.6062,n=30),模拟值相对观测值的平均绝对误差(Dabs)分别为18.10和16.97g C/m2。
1960-2005年CENTURY模拟的高寒草甸土壤异养呼吸CO2通量呈波动性缓慢上升趋势,变化范围在479.22~624.89g C/(m2·a)之间,平均值为(539.56±34.32)g C/(m2·a),通量增加率为16.5g C/(m2·10a)。模拟结果与主要气候因子进行的相关分析表明,高寒草甸土壤异养呼吸与气温呈显著正相关(r=0.7,P<0.05),与降水量相关性不显著。
氮沉降倍增显著促进了高寒草甸土壤异养呼吸CO2释放。对照(CK)、中氮(MN)和高氮(HN)情景下,土壤异养呼吸CO2通量变化范围分别为479.22~624.89g C/(m2·a),582.16~828.57g C/(m2·a)和581.08~837.64g C/(m2·a),平均值分别为(539.56±34.32)g C/(m2·a),(657.01±51.08)g C/(m2·a)和(658.58±54.05)g C/(m2·a)。中氮(MN)、高氮(HN)情景下CO2通量与对照相比分别增加了21.76%和22.06%,但由于呼吸作用对氮沉降可能存在着一定的“氮饱和”现象,随着氮沉降的倍增,其促进效应降低。
[1]IPCC.Summary for Policy makers of Climate Change 2007:The Physical Science Basis.Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change[M].Cambridge:Cambridge University Press,2007.
[2]Galloway J N,Aber J D,Erisman J W,etal.The nitrogen cascade.Bio-Science,2003,53:341-356.
[3]Kaiser J.The other global pollutant:nitrogen proves tough to curb.Science,2001,294:1268-1269.
[4]Moffat A S.Global nitrogen overload problem grows critical.Science,1989,244:776-783.
[5]Galloway J N,Levy H,Kasibhatla P S.Year 2020:consequence of population growth and development on deposition of oxidized nitrogen.Ambio,1994,23:120-123.
[6]Pamela M,Kathleen A L,Sharon J H.The globalization of nitrogen deposition:Consequences for terrestrial ecosystem.Ambio,2002,31(2):113-121.
[7]Bai W.The Effects of Global Climate Changes of Alpine Grassland Ecosystem and its Carbon Emission in Head-water Region of the Yongtze River[D].Lanzhou:Lanzhou University,2010.
[8]Cheng G D,Li P J,Zhang X S,etal.Impacts of Climate Change on Glaciers,Permafrost and Snow Cover in China[M].Lanzhou:Gansu Culture Press,1997:22-56.
[9]Li L,Chen X G,Wang Z Y,etal.Climate change and its regional differences over the Tibetan plateau.Advances in Climate Change Research,2010,6(3):181-186.
[10]Ding Y H,Ren G Y,Shi G Y,etal.National assessment report of climate change(I):Climate change in China and its future trend.Advances in Climate Change Research,2006,2(1):3-8.
[11]Lv C,Tian H.Spatial and temporal patterns of nitrogen deposition in China:Synthesis of observational data.Journal of Geophysical Research:Atmospheres(1984-2012),2007,112:D22S05,doi:10.1029/2006JD007990.
[12]Zheng X H,Fu C B,Xu X K,etal.The Asian nitrogen cycle case study.Ambio,2002,31:79-87.
[13]Galloway J N,Dentener J F,Capone G D,etal.Nitrogen cycles:past,resent and future.Biogeochemistry,2004,70:153-226.
[14]Yang Y H,Fang J Y,Tang Y H,etal.Storage,patterns and controls of soil organic carbon in the Tibetan grasslands.Global Change Biology,2008,14:1592-1599.
[15]Li Y N,Wang Q J,Zhao X Q,etal.The influence of climatic warming on the climatic potential productivity of alpine meadow.Acta Agrestia Sinica,2000,18(1):23-29.
[16]Zhang J H,Li Y N.The research on effect of climate change on vegetation productivity in Qinghai province.Journal of Arid Land Resources and Environment,2008,22(2):97-102.
[17]Li Y N,Wang X Q,Gu S,etal.Integrated monitoring of alpine vegetation types and its primary production.Acta Geographica Sinica,2004,59(1):40-48.
[18]Yang K,Lin E D,Gao Q Z,etal.Simulation of climate change impacts on grassland productivity in Northern Tibet.Chinese Journal of Ecology,2010,29(7):1469-1476.
[19]Wang G X,Qian J,Cheng G D,etal.Soil organic carbon pool of grassland soils on the Qinghai Tibetan Plateau and its global implication.The Science of the Total Environment,2002,291:207-217.
[20]Zeng Y N,Feng D,Cao G C,etal.The soil organic carbon storage and its spatial distribution of alpine grassland in the source region of the Yellow river.Acta Geographica Sinica,2004,59(4):497-504.
[21]Tao Z,Shen C D,Gao Q Z,etal.Soil organic carbon storage and vertical distribution of alpine meadow on the Tibetan plateau.Acta Geographica Sinica,2006,61(7):720-728.
[22]Zhang Y Q,Tang Y H,Jiang J.The dynamics characteristics of soil organic carbon of grassland on Qinghai Tibet Plateau,Science in China Ser.D Earth Sciences,2006,36(12):1140-1147.
[23]Toshiyuki O,Mitsuru H,Zhan X Z,etal.Soil organic carbon pools in alpine to nival zones along an altitudinal(4400-5300 m)on the Tibetan Plateau.Polar Science,2008,2:277-285.
[24]Li D,Huang Y,Wu Q,etal.Modeling dynamics of soil organic carbon in an alpine meadow ecosystem on the Qinghai-Tibetan plateau using CENTURY model.Acta Prataculturae Sinica,2010,19(2):160-168.
[25]Hu Q W,Cao G M,Wu Q,etal.Comparative study on CO2emissions from different types of alpine meadow during grass exuberance period.Journal of Georaphical Science,2004,14(2):167-176.
[26]Li D,Cao G M,Hu Q W,etal.A primary study on CO2emission from alpinePotentillafruticosascrub meadow ecosystem.Acta Agrestia Sinica,2005,13(2):144-148.
[27]Li D,Cao G M,Wu Q,etal.The seasonal change rule of CO2release rate in alpinePotentillafruticosashrub meadow ecosystem.Pratacultural Science,2005,22(5):4-10.
[28]Wu Q,Hu Q W,Cao G M,etal.A primary study on CO2emission from soil-plant systems ofKobresiahumilismeadow.Resources Science,2005,27(2):96-101.
[29]Zhao L,Xu S X,Li Y N,etal.Relations between carbon dioxide fluxes and environmental factors ofKobresiahumilismeadows andPotentillafruticosameadows.Acta Botanica Boreali Occidentalia Sinica,2006,26(1):133-142.
[30]Hu Q W,Wu Q,Cao G M,etal.Growing season ecosystem respirations and associated component fluxes in two alpine meadows on the Tibetan Plateau.Journal of Integrative Plant Biology,2008,50(3):271-279.
[31]Zhu T H,Cheng S L,Fang H J,etal.Early responses of soil CO2emission to simulating atmospheric nitrogen deposition in an alpine meadow on the Qinghai Tibetan Plateau.Acta Ecologica Sinica,2011,31(10):2687-2696.
[32]Wu Q,Hu Q W,Cao G M,etal.CO2emission from an alpineKobresiahumilismeadow in winters.Acta Ecologica Sinica,2012,31(18):5107-5122.
[33]Parton W J,Schimel,D S,Cole C V,etal.Analysis of factors controlling soil organic matter levels in Great Plains grasslands.Soil Science Society America Journal,1987,51:1173-1179.
[34]Smith P,Smith J U,Powlson D S,etal.A comparison of the performance of nine soil organic matter models using datasets from seven longterm experiments.Geoderma,1997,(81):153-225.
[35]Jenkinson D S,Rayner J H.The turnover of organic matter in some of the Rothamsted classical experiments.Soil Science,1977,123:298-305.
[36]Zhou X M.KobresiaMeadow in China[M].Beijing:Science Press,2001.
[37]Lv X M,Zheng D.Impacts of global change on the alpine meadow ecosystem in the source region of the Yangtze River.Resources and Environment in the Yangtze Basin,2006,15(5):603-607.
[38]Raich J W,Tufekciogul A.Vegetation and soil respiration:correlations and controls.Biogeochemistry,2000,48(1):71-90.
[39]Qi Y,Xu M,Wu J G.Temperature sensitivity of soil respiration and its effects on ecosystem carbon budget:nonlinearity begets surprises.Ecological Modelling,2002,153(1/2):131-142.
[40]Bond-Lamberty B,Thomson A.Temperature-associated increases in the global soil respiration record.Nature,2010,464:579-582.
[41]Cao M K,Prince S D,Li K R,etal.Response of terrestrial carbon uptake to climate interannual variability in China.Global Change Biology,2003,9(4):536-546.
[42]Rustad L E,Campbell J L,Marion G M,etal.A meta-analysis of the response of soil respiration,net nitrogen mineralization,and aboveground plant growth to experimental ecosystem warming.Oecologia,2001,126:543-562.
[43]Luo Y Q,Wan S Q,Hui D F,etal.Acclimatization of soil respiration to warming in a tall grass prairie.Nature,2001,413:622-624.
[44]Lisji J,Nissinen A,Erhard M,etal.Climatic effects on litter decomposition from arctic tundra to tropic rain forest.Global Change Biology,2003,9(4):575-584.
[45]Melillo J M,Steudler P A,Aber J D,etal.Soil warming and carbon-cycle feedbacks to the climate system.Science,2002,298(13):2173-2175.
[46]Tjoelker M G,Oleksyn J,Reich P B.Acclimation of respiration to temperature and CO2in seedlings of boreal tree species in relation to plant size and relative growth rate.Global Change Biology,1999,5(4):679-691.
[47]Li H J.Studies on Soil Respiration and its Relations to Environmental Factors in Different Ecosystems[D].Taiyuan:Shanxi University,2008.
[48]Flanagan L B,Johnson B G.Interacting effects of temperature,soil moisture and plant biomass production on ecosystems respiration in a northern temperate grassland.Agricultural and Forest Meteorology,2005,130(3-4):237-253.
[49]Li Q,Xue H X,Wang Y L,etal.The preliminary study on the impact of soil temperature and moisture on carbon flux overStipakryloviiecosystem,Journal of Agro-Environment Science,2011,30(3):605-610.
[50]Dong Y S,Qi Y C,Liu J Y,etal.The soil respiration flux variation in different rainfall intensity of four kinds of grassland.Chinese Science Bulletin,2005,50(5):473-480.
[51]Bouma T J,Bryla D R.On the assessment of root and soil respiration for soils of different textures:Interactions with soil moisture contents and soil CO2concentrations.Plant and Soil,2000,227(1-2):215-221.
[52]Silvola J,Alm J,Ahlholm U.The effect of plant roots on CO2release from peat soil.Suo,1992,43:259-262.
[53]Kucera C L,Kirkham D R.Soil respiration studies in tall-grass prairie in Missouri.Ecology,1971,52(5):912-915.
[54]Upadhyaya S D,Singh V P.Microbial turnover of organic matter in a tropical grassland soil.Pedobiologia,1981,21(2):100-109.
[55]Coleman D C.Soil carbon balance in a successional grassland.Oikos,1973,24:195-199.
[56]Xie W,Chen S T,Hu Z H.Factors influencing variability in soil heterotrophic respiration from terrestrial ecosystem in China.Environment Science,2014,35(1):334-340.
[57]Leirós M C,Trasar-Cepeda C,Gil-Sotres F.Dependence of mineralization of soil organic matter on temperature and mois-ture.Soil Biology and Biochemistry,1999,31(3):327-335.
[58]Yu W C,Song X L,Xiu W M,etal.Effects of additional nitrogen on litter decomposition inStipabaicalensisgrassland.Acta Prataculturae Sinica,2014,23(5):49-60.
[59]Cao C C,Qi Y C,Dong Y S,etal.Effects of nitrogen deposition on critical fractions of soil organic carbon in terrestrial ecosystems.Acta Prataculturae Sinica,2014,23(2):323-332.
[60]Bowden R D,Davidson E,Savage K,etal.Chronic nitrogen additions reduce total soil respiration and microbial respiration in temperate forest soils at the Harvard Forest.Forest Ecology and Management,2004,196:43-56.
[61]Olsson P,Linder S,Giesler R,etal.Fertilization of boreal forest reduces both autotrophic and heterotrophic soil respiration.Global Change Biology,2005,11(10):1745-1763.
[62]Jia B R,Zhou G S,Wang F Y,etal.Affecting factors of soil microorganism and root respiration.Chinese Journal of Applied Ecology,2005,16(8):1547-1552.
[63]Emmett B A.The impact of nitrogen on forest soils and feedbacks on tree growth.Water,Air and Soil Pollution,1999,116:65-74.
参考文献:
[7]白炜.长江源区高寒草地生态系统变化及其碳排放对气候变化的响应[D].兰州:兰州大学,2010.
[8]程国栋,李培基,张祥松,等.气候变化对中国积雪、冰川和冻土的影响评价[M].兰州:甘肃文化出版社,1997:22-56.
[9]李林,陈晓光,王振宇,等.青藏高原区域气候变化及其差异性研究.气候变化研究进展,2010,6(3):181-186.
[10]丁一汇,任国玉,石广玉,等.气候变化国家评估报告(Ⅰ):中国气候变化的历史和未来趋势.气候变化研究进展,2006,2(1):3-8.
[15]李英年,王启基,赵新全.气候变暖对高寒草甸气候生产潜力的影响.草地学报,2000,18(1):23-29.
[16]张景华,李英年.青海气候变化趋势及对植被生产力影响的研究.干旱区资源与环境,2008,22(2):97-102.
[17]李英年,王学勤,古松,等.高寒植被类型及其植被生产力的监测.地理学报,2004,59(1):40-48.
[18]杨凯,林而达,高清竹,等.气候变化对藏北地区草地生产力的影响模拟.生态学杂志,2010,29(7):1469-1476.
[20]曾永年,冯东,曹广超,等.黄河源区高寒草地土壤有机碳储量及分布特征.地理学报,2004,59(4):497-504.
[21]陶贞,沈承德,高全洲,等.高寒草甸土壤有机碳储量及其垂直分布特征.地理学报,2006,61(7):720-728.
[22]张永强,唐艳鸿,姜杰.青藏高原草地生态系统土壤有机碳动态特征.中国科学D辑:地球科学,2006,36(12):1140-1147.
[24]李东,黄耀,吴琴,等.青藏高原高寒草甸生态系统土壤有机碳动态模拟研究.草业学报,2010,19(2):160-168.
[26]李东,曹广民,胡启武,等.高寒灌丛草甸生态系统CO2释放的初步研究.草地学报,2005,13(2):144-148.
[27]李东,曹广民,吴琴,等.海北高寒灌丛草甸生态系统CO2释放速率的季节变化规律.草业科学,2005,22(5):4-10.
[28]吴琴,胡启武,曹广民,等.矮嵩草草甸植被-土壤系统CO2的释放特征.资源科学,2005,27(2):96-101.
[29]赵亮,徐世晓,李英年,等.青藏高原矮嵩草草甸和金露梅灌丛草甸CO2通量变化与环境因子的关系.西北植物学报,2006,26(1):0133-0142.
[31]朱天鸿,程淑兰,方华军,等.青藏高原高寒草甸土壤CO2排放对模拟氮沉降的早期响应.生态学报,2011,31(10):2687-2696.
[32]吴琴,胡启武,曹广民,等.矮嵩草草甸冬季CO2的释放特征.生态学报,2012,31(18):5107-5122.
[36]周兴民.中国嵩草草甸[M].北京:科学出版社:2001.
[37]吕新苗,郑度.气候变化对长江源地区高寒草甸生态系统的影响.长江流域资源与环境,2006,15(5):603-607.
[47]李洪建.不同生态系统土壤呼吸与环境因子的关系研究[D].太原:山西大学,2008.
[49]李琪,薛红喜,王云龙,等.土壤温度和水分对克氏针茅草原生态系统碳通量的影响初探.农业环境科学学报,2011,30(3):605-610.
[50]董云社,齐玉春,刘纪远,等.不同降水强度4种草地群落土壤呼吸通量变化特征.科学通报,2005,50(5):473-480.
[56]谢薇,陈书涛,胡正华.中国陆地生态系统土壤异养呼吸变异的影响因素.环境科学,2014,35(1):334-340.
[58]于雯超,宋晓龙,修伟明,等.氮素添加对贝加尔针茅草原凋落物分解的影响.草业学报,2014,23(5):49-60.
[59]曹丛丛,齐玉春,董云社,等.氮沉降对陆地生态系统关键有机碳组分的影响.草业学报,2014,23(2):323-332.
[62]贾丙瑞,周广胜,王风玉,等.土壤微生物与根系呼吸作用影响因子分析.应用生态学报,2005,16(8):1547-1552.
