草鱼♀×鳡♂杂交F1及其亲本的形态差异分析
2014-12-28余来宁夏小平刘红艳张繁荣江汉大学生命科学学院湖北武汉430056武汉市四汇水产科技开发有限公司湖北武汉430065
余来宁,夏小平,杨 东,刘红艳,张繁荣(.江汉大学生命科学学院,湖北武汉430056;.武汉市四汇水产科技开发有限公司,湖北武汉430065)
草鱼(Ctenopharyngodon idellus)以草为食,生长快,养殖成本低,是我国重要的淡水养殖对象,但抗病力差。鳡(Elopichthys bambusa)是凶猛性鱼类,以活鱼为食,具有生长快和抗病力强等优点。二者同属于雅罗鱼亚科,是1对较为理想的杂交组合。为了探讨提高草鱼的抗病力。近年来,笔者开展了草鱼♀×鳡♂杂交试验(简称草鳡杂交),初步观察草鳡杂交F1代具有较强的杂交优势[1-2]。草鳡杂交F1形态上与母本草鱼十分相似,仅凭外观难以分辨,因此对其进行系统的形态差异分析十分必要。
传统鱼类形态学的研究方法,主要测定鱼类的可数性状和可量性状,是分类学上种间鉴别的重要手段,但是传统测量法主要集中在鱼体的头、尾部和轴向,这对种内群体及杂交后代的鉴别十分有限。于是,一种能够提供更多信息的框架测量法应运而生[3],并已经在鱼类形态差异研究方面被广泛应用[4-9]。笔者采用传统形态学结合现代框架测量法对草鱼♀×鳡♂杂交F1及其亲本的形态进行了观测,并应用方差分析、聚类分析、主成分分析和判别分析等多元分析方法进行了分析,旨在了解3种鱼间的形态差异,以期为草鳡杂交选育提供依据。
1 材料与方法
1.1 试验材料 试验所用草鱼、鳡及草鳡杂交F1代来源于武汉江夏区鲁湖渔场,每种鱼取样30尾进行测量,3种鱼的体长范围分别为:草鱼232~317 mm;鳡357~436 mm;草鳡杂交F1284~372 mm。
1.2 形态测量 使用量鱼板、直尺、游标卡尺及外卡规测量鱼的传统形态学参数和框架结构参数,精确到1 mm。传统形态学指标,包括全长、体长、体高、体厚、头长、吻长、眼径、眼间距、尾柄长、尾柄高(图1)。可数性状包括侧线鳞、鳍棘、鳍条、鳃耙等。框架结构选择10个坐标点,测定坐标点之间的距离。A为胸鳍起点,B为吻端,C为腹鳍起点,D为鳃盖上端,E为臀鳍起点,F为背鳍起点,G为臀鳍基后端,H为背鳍基后端,I为尾鳍基下端,J为尾鳍基上端),测量项目有 AB、AC、AD、BC、BD、CD、CE、CF、DE、DF、EF、EG、EH、FG、FH、GH、GI、GJ、HI、HJ、IJ 21 项,如图2 所示。
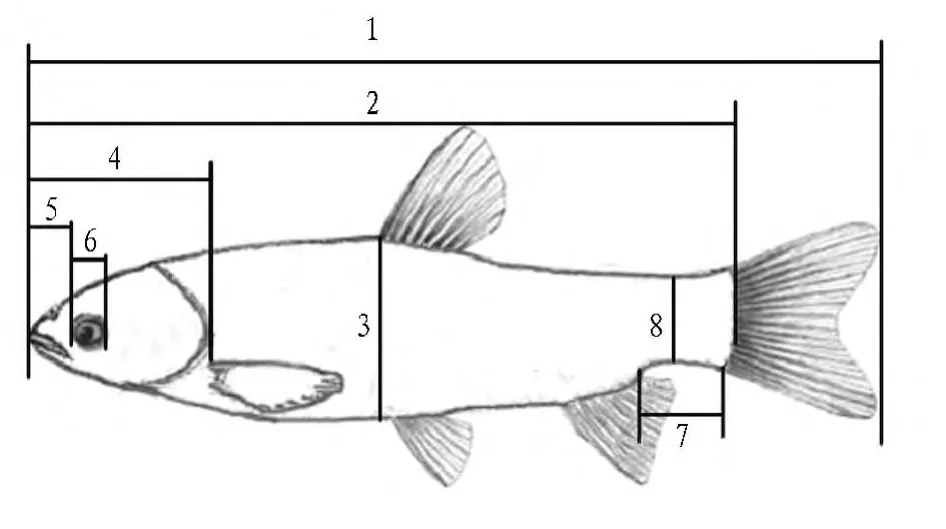
图1 鱼类传统形态测量示意
1.3 数据统计与分析 使用SPSS 17.0软件对试验数据进行统计与分析。对可数性状进行卡方分析,比较种间差异。对传统可量性状进行LSD显著性检验。对传统可量比例性状和框架数据校正后进行聚类分析、主成分分析和判别分析。杂交指数(HI)的计算参照文献[10]的方法。杂交指数(HI)计算公式为:HI=100 ×(Hi-Mi1)/(Mi2- Mi1)。式中,Hi为杂种平均值;Mi1为母本平均值;Mi2为父本平均值;HI介于45~55属于中间性状;若HI<45,为偏母本性状;若HI>55,为偏父本性状;若HI>100或 HI<0,为超亲偏离性状。

图2 鱼类框架测量示意
2 结果与分析
2.1 外观及可数性状 草鳡杂交F1外观上与草鱼十分相似,体形呈长圆筒形,前部圆,后部稍扁。头中等大,口端位,呈弧形,上颌稍突出。鳞片圆大,体色草黄,背部青绿,胸鳍和腹鳍略带褐黄色。背鳍无硬刺,背鳍起点与腹鳍起点相对,尾鳍呈叉形。鳡鱼的外观形态有较大差异,鳡鱼体型侧扁修长,体高和体厚均很小,体长为体高8倍多。头小较长,吻尖,口裂大,上下颌有凹凸结构;鳞片细小,体色背部灰黑,腹部银白,胸鳍、腹鳍和臀鳍呈黄色;背鳍起点在腹鳍起点之后。3种鱼的外形见图3。

图3 草鳡杂交F1和亲本草鱼及鳡鱼形态观察
通过测定可知,3种鱼鳍棘数相同,背、臀鳍棘数为3;胸、腹鳍棘数为1;分枝鳍条数草鱼与鳡鱼差异较大,而草鳡杂交F1代的分枝鳍条除胸鳍条外基本与母本草鱼相同,3种鱼的可数性状测定数据见表1。

表1 草鱼♀×鳡♂杂交F1及其亲本的可数性状
通过卡方分析发现,杂交F1和草鱼与鳡鱼的4种分枝鳍条、侧线鳞和鳃耙数均存在显著差异(P<0.05),而杂交F1代与草鱼仅在胸鳍条和侧线鳞上存在显著差异(P<0.05)。通过对杂交指数(HI)进行计算可知,F1代除了胸鳍条为偏父本性状外,其他可数性状均为偏母本性状。可数性状杂交F1的平均杂交指数为17.16,为偏母本遗传。
2.2 可量性状的方差分析 由表2可知,经LSD显著性检验,杂交F1和草鱼的所有可量性状与鳡鱼均存在显著差异;而杂交F1与草鱼除头长/眼径、头长/眼间距、尾柄长/尾柄高无显著差异外,其他可量性状均有显著差异。从杂交指数(HI)可以看出,草鳡杂交F1代的体长/体高、体长/头长、头长/眼径等3项性状的杂交指数(HI)值分别为3.9、37.5和4.0,均小于45为,偏母本性状。其他性状HI>100或HI<0,均为超亲偏离性状[10],可量性状的平均杂交指数为40.27,草鳡杂交F1代可数可量性状总杂交指数平均值为28.71,为偏母本遗传。
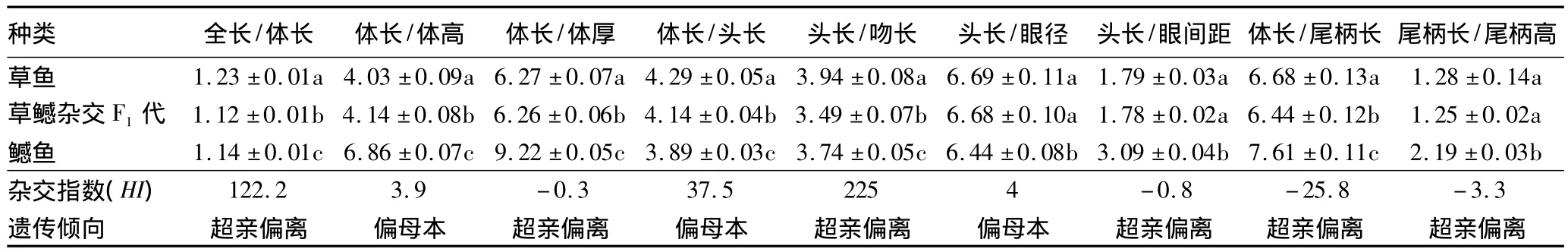
表2 草鳡杂交F1鱼及亲本的可量性状的LSD多重比较
2.3 可量比例性状和框架数据分析
2.3.1 聚类分析。采用欧氏距离对9项传统可量比例性状和21项校正后的框架数据进行聚类分析,结果发现草鱼与草鳡杂交F1代的平均形态距离为0.559,与鳡鱼的距离为4.50,杂交F1与鳡鱼的距离为4.494。从图4可以看出,草鳡杂交F1代与草鱼聚在一支,而鳡鱼为另一支,说明杂交F1形态距离与母本草鱼接近。

图4 草鳡杂交F1与亲本的聚类分析结果
2.3.2 主成分分析。对30项形态参数进行了主成分分析,从中提取了3个主成分,其对总变异的贡献率依次为59.525%、25.08% 和 3.31%,对总变异的累计贡献率为87.915%,已包含了3种鱼形态总变异的大部分,这说明可以用几个相互独立的因子来概括种间的形态差异。由表3可知,主成分1载荷较大的参数有体长/体厚、头长/眼间距、体长/体高、EH/体长、尾柄长/尾柄高、EF/体长、体长/尾柄长,主要反映鱼体形沿体长纵向和横向的变化。主成分2中载荷较大的参数有DE/体长、AC/体长、头长/吻长、全长/体长、CE/体长。主成分2主要反映了鱼头尾的变化以及腹鳍和臀鳍的位置信息。这与上述外形比较结果基本一致。
从图5可以看出,由主成分1对主成分2的散布图可以看出3种鱼能较好区分,而主成分1对主成分3的散布图中杂交F1与草鱼有部分重叠。由此可见,草鱼、鳡鱼及草鳡杂交F1在形态上的差异很大程度上是由体长、体高、体厚、头长以及腹鳍、背鳍和臀鳍位置等的变化所导致的。
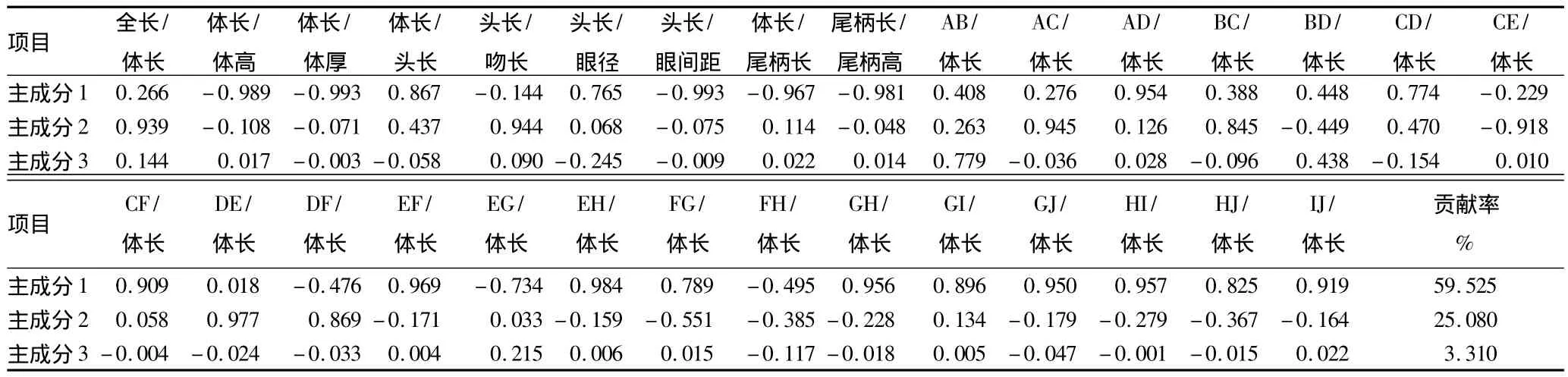
表3 形态参数前3个主成分载荷及对其总变异的贡献率
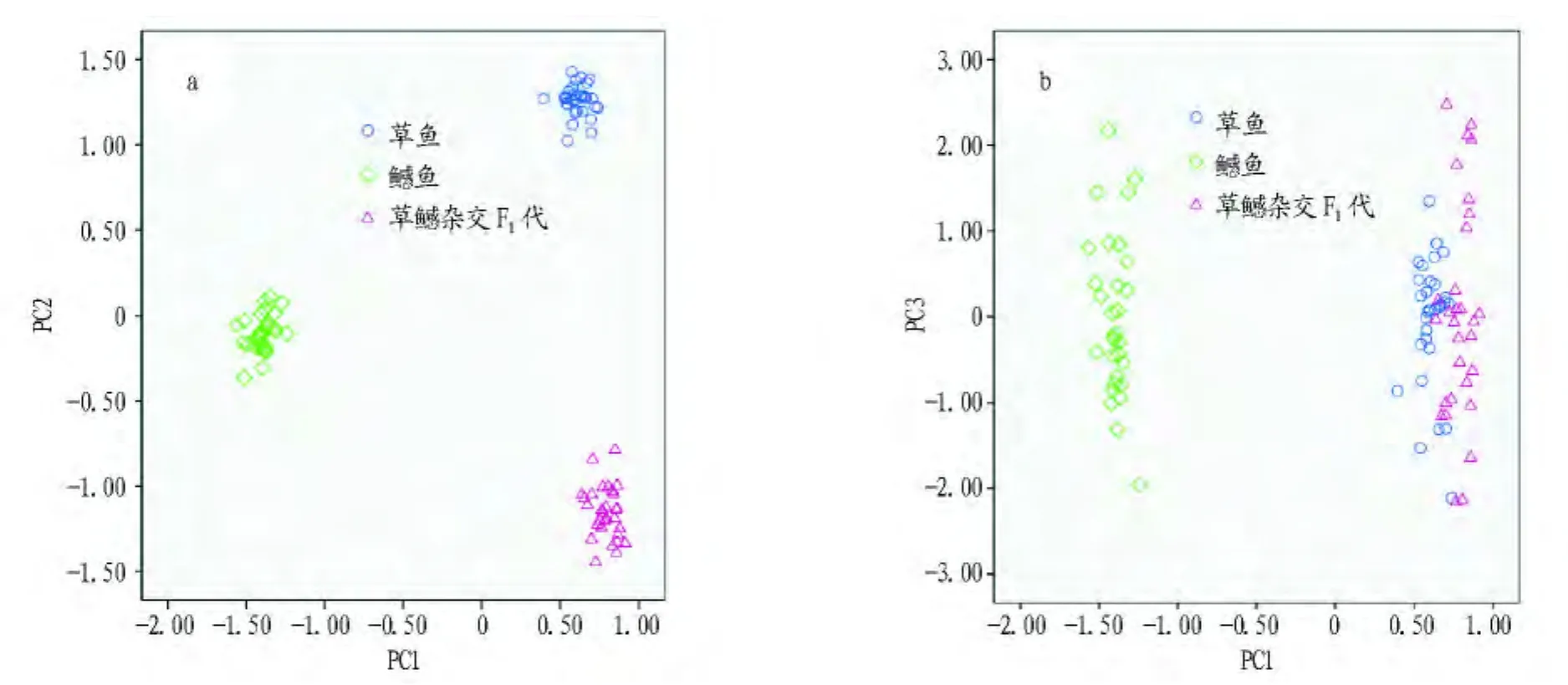
图5 草鳡杂交F1及亲本主成分散布图示意
2.3.3 判别分析。采用逐步判别法对草鳡杂种F1及父母本的30个形态比例参数进行分析,选取贡献最大的2个变量参数EH/体长(X1)和DF/体长(X2),构建了3种鱼的判别函数(Y1、Y2、Y3 分别代表草鱼、杂交 F1、鳡鱼),判别公式为:

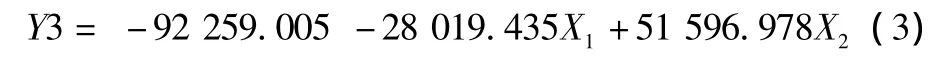
由表4可知,用以上判别函数对观测样本进行预测分类,3种鱼的判别准确率均达到100%,没有出现误判,清楚地显示出它们的形态差异。
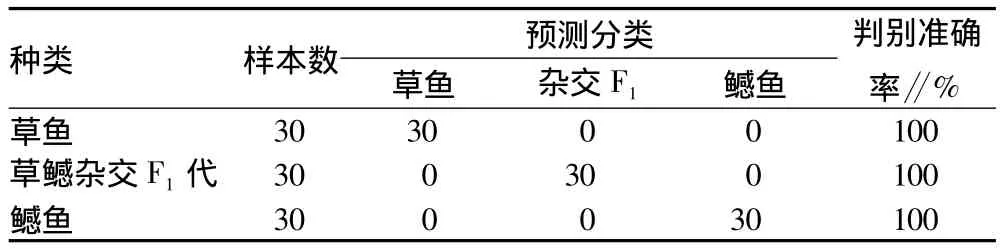
表4 草鳡杂交F1及亲本草鱼和鳡的判别分析结果
3 讨论
经典的鱼类分类学主要是以鱼的形态为分类依据。可数性状是鱼类分类学上鉴别物种的重要指标[11]。草鱼与鳡鱼是不同属的鱼类,外部形态差异明显。草鱼与鳡鱼的可数性状中有一部分是相同的,如背鳍、胸鳍、腹鳍、臀鳍的鳍棘数均分别为3、1、1、3,也有一部分性状是不同的,如分枝鳍条、侧线鳞、鳃耙数存在差异。草鳡杂交F1代大部分可数性状却并不是中间类型。从可数性状的卡方检验结果可以看出,草鱼与鳡鱼在所检测的6项指标上均存在显著差异,容易将二者区分开来。但是,杂交F1在体色和外观以及可数性状上与草鱼十分相似,背鳍、腹鳍、臀鳍的分枝鳍条数是完全相同的,仅胸鳍条数有差异,这种情况在其他属间杂交鱼中也出现过。例如,江黄颡和乌苏里拟鲿杂交后代[12]属间杂交在胸鳍条数产生差异是否为普遍现象,仍值得进一步研究。虽然草鱼与杂交F1胸鳍条和侧线鳞在30个样本检测上存在显著差异,而作为单个个体测定时,由于2种鱼指标有重叠,是不能完全准确将其区分开的。多元分析方法能够较好地判别种类间的差异,已被广泛用于鱼类杂交后代与亲本的关系研究[13-15]。这说明用多元分析方法研究草鳡杂交F1与其亲本间的形态差异确有必要。
从杂交指数可以看出,草鳡杂交F1代大部分可数可量性状都是偏母本的。这说明杂交F1在外形上有较强偏母性遗传倾向。聚类分析结果表明,杂交F1与草鱼聚为一支,进一步证实了以上结论。母性遗传的现象在鱼类杂交中较多,如荷包红鲤♀×元江鲤♂[16]、乌苏里拟鲿♀×江黄颡♂、团头鲂♀×三角鲂♂[17]、奥利亚罗非鱼♀×尼罗罗非鱼♂[18]、黑龙江野鲤♀×建鲤♂[19]等,这可能与杂交中母本提供卵子以及受线粒体母性遗传的影响有关,但也有一些杂交后代性状呈中间型或偏父本的情况[20-21],显然杂交后代的遗传倾向性是一个相当复杂的问题,尚有待进一步研究。
从判别分析结果来看,对其影响最大的2个变量是臀鳍起点至背鳍基后端的距离和鳃盖前端上侧至背鳍起点的距离,这2个性状均属于框架参数,仅用这2个变量建立的判别公式,就可以将3种鱼鉴别出来,准确率达100%。这说明框架参数中包涵了形态差异的重要信息,由此可见选用框架参数进行鱼类形态差异分析是非常必要的。
[1]余来宁,夏小平,杨东,等.草鱼♀×鳡♂杂交F1代胚胎发育及胚后发育研究[J].安徽农业科学,2014,42(20):6639 -6645.
[2]余来宁,夏小平,杨东,等.草鱼♀×鳡♂杂交F1代同工酶和蛋白质的电泳分析[J].安徽农业科学,2014,42(30):10573 -10575.
[3]BOOKSTEIN F L.Foundation of morphometrics[J].Ann Rev Ecol Syst,1982,13:451 -470.
[4]李思发,李晨虹,李家乐.尼罗罗非鱼品系间形态差异分析[J].动物学报,1998,44(4):450 -457.
[5]张健东,马龙,陈刚,等.不同地理群体多鳞鱚的形态差异分析[J].安徽农业科学,2010,38(11):5678 -5681.
[6]王伟,陈立侨,顾志敏,等.7个不同翘嘴红鲌群体的形态差异分析[J].淡水渔业,2007,37(3):40 -44.
[7]姚景龙,陈毅峰,李垫,等.中华鮡与前臀鮡的形态差异和物种有效性[J].动物分类学报,2006,31(1):11 -17
[8]刘楚珠,严利平,李建生,等.基于框架法的东黄海日本鲭产卵群体形态差异分析[J].中国水产科学,2011,18(4):908 -917.
[9]王飞全,李强,易祖盛,等.光倒刺鲃不同种群的形态差异分析[J].安徽农业科学,2013,41(6):2464 -2465.
[10]CRICELLI A,DUPONT F.Biometrical and biological features of Alburnus alburnus×Rutilus rubilio natural hybrids from Lake Mikni Prespa,northern Greece[J].Journal of Fish Biology,1988,31(6):721 -733.
[11]伍献文.中国鲤科鱼类志(上卷)[M].上海:上海科学技术出版社,1964-1982:13-40.
[12]蔡永祥,陈友明,陈校辉,等.江黄颡和乌苏里拟鲿杂交F1形态差异[J].湖泊科学,2011,23(2):264 -270.
[13]王燕,张勇,张海发,等.两种杂交石斑鱼及其亲本的形态差异分析[J].水产学报,2014,38(6):778 -785.
[14]李思发,陈林,李先仁,等.“吉奥”罗非鱼同其亲本及近缘杂交罗非鱼形态差异比较分析[J].上海海洋大学学报,2010,19(6):721 -727.
[15]郭诗照,王荣泉,傅建军,等.草鱼、鳙及其杂交鱼的形态差异分析[J].江苏农业科学,2011,39(5):320 -322.
[16]国家水产总局长江水产研究所育种室鲤鱼组.荷元鲤(荷包红鲤♀×元江鲤♂)杂种优势利用及性状遗传的研究[J].淡水渔业,1981(3):2-9.
[17]杨怀宇,李思法,邹曙明.三角鲂与团头鲂正反杂交F1的遗传性状[J].上海水产大学学报,2002,11(4):305 -309.
[18]李家乐,李思发,李勇,等.尼奥鱼[尼罗罗非鱼♀×奥利亚罗非鱼♂]同其亲本的形态和判别[J].水产学报,1999,23(3):261 -265.
[19]李冰,柴学森,张成锋,等.建鲤和黑龙江野鲤自交和正反交F1形态差异分析[J].中国水产科学,2014,21(1):59 -66.
[20]赵建,朱新平,陈永乐,等.翘嘴鳜、斑鳜及其杂交种形态差异分析[J].华中农业大学学报,2008,27(4):506 -509.
[21]顾志敏,贾永义,叶金云,等.翘嘴红鲌♀×团头鲂♂杂种F1的形态特征及遗传分析[J].水产学报,32(4):533 -544.
