基于ISSR标记的少果毛一球悬铃木遗传稳定性分析
2014-12-27张丽芳涂炳坤丁昭全
姚 军,张丽芳,涂炳坤,丁昭全
(1.武汉市园林科学研究所,湖北 武汉 430081;2. 华中农业大学 园艺林学学院,园艺植物生物学教育部重点实验室,湖北 武汉 430070)
基于ISSR标记的少果毛一球悬铃木遗传稳定性分析
姚 军1,张丽芳2,涂炳坤2,丁昭全1
(1.武汉市园林科学研究所,湖北 武汉 430081;2. 华中农业大学 园艺林学学院,园艺植物生物学教育部重点实验室,湖北 武汉 430070)
本研究应用ISSR分子标记,对3个不同立地条件下的不同嫁接年限的少果毛一球悬铃木优良品种‘园科115’一球悬铃木Platanus occidentalis‘Yuanke 115’的接穗(13 a、14 a与30 a)以及两个相应的砧木(二球悬铃木)进行遗传稳定性分析。16条引物在5个样品中共扩增出75条谱带,其中27条具有多态性,多态性比例为36%。根据ISSR标记数据,基于遗传相似系数的聚类分析显示:5个悬铃木样品间的遗传相似系数在0.680 0~1.000之间,遗传相似系数在0.77时,5个悬铃木样品归为一类;遗传相似系数在0.98时,3个不同嫁接年限的‘园科115’一球悬铃木接穗归为一类。其中,嫁接14 a与30 a的一球悬铃木接穗的遗传相似系数为1,它们与嫁接13 a的一球悬铃木接穗的遗传相似系数也高达0.986 7。以上结果说明:‘园科115’一球悬铃木在不同的立地条件下,经历不同的嫁接时间(最长达30 a),依然具有高度的遗传稳定性。
‘园科115’一球悬铃木;分子标记;遗传稳定性;ISSR分子标记
悬铃木是全世界温带及亚热带广为应用的园林绿化树种,拥有无与伦比的优良特性:生长迅速、冠大荫浓、适应性强以及抗性优良,被誉为“行道树之王”。但是,悬铃木果毛污染让人深受其害。从自然界中选育遗传稳定的少果毛的突变品种,对现有的悬铃木进行换冠嫁接,不啻为一种有效可行的解决手段。少果毛悬铃木品种经历长时间的嫁接后,能否保持遗传稳定是非常关键而迫切需要回答的核心问题,关系到品种能否成功推广。
‘园科115’一球悬铃木Platanus occidentalis‘Yuanke 115’是武汉市园林科学研究所选育出的少果毛悬铃木品种。该品种在武汉地区春季发芽较二球悬铃木P. acerifolia晚7 d左右,秋季落叶较二球悬铃木缩短2~4周,落叶时间短而集中,具有较强的顶端生长优势和明显主枝。该品种在酸、碱性土壤和地下水位高的条件下都能正常生长发育,对高温及辐射热有较好的抗性,扦插与嫁接成活率较高,具有生长快、适应强、萌芽力强、耐修剪等特性,是一个优良的少果毛品种[1]。
ISSR(inter-simple sequence repeat)分子标记是1994年由Zietkiewicz等[2]创建的一种新型DNA分子标记技术,用于检测简单重复序列间DNA序列的多态性。它是基于SSR(simple sequence repeat)基础上,在SSR一段序列的一端上加若干核苷酸碱基作为扩增引物,从而达到扩增SSR间隔序列的目的。与其它分子标记技术相比,ISSR特别适用于基因组信息知之不多的生物,具有引物设计简单、稳定性及重复性强、操作简洁、谱带多、成本低等特点。
ISSR技术在园林植物中也应用广泛,主要应用于:品种亲缘关系研究[3-7]、资源植物遗传多样性[8-11]、品种鉴定与分类[12-13]等。目前对于悬铃木而言,主要采用AFLP、SSR分子标记等方法进行悬铃木遗传变异、遗传多样性及稳定性的研究。其中黄文俊[14]、王艳梅[15]建立了ISSR分子标记方法,筛选出16条ISSR引物,可用于无果悬铃木的遗传分析。
本研究将对武汉市园林科学研究所提供的3个处于不同立地条件下的少果毛一球悬铃木嫁接单株,即嫁接13 a、14 a以及30 a的单株,进行ISSR分子标记,以确定‘园科115’一球悬铃木的遗传稳定性。
1 材料与方法
1.1 材料
1.1.1 试验材料
5个悬铃木样品:嫁接14 a的少果毛一球悬铃木接穗(14优,简称a)、嫁接14 a的二球悬铃木砧木(14砧,简称b)、应用于单位绿地;嫁接30 a的少果毛一球悬铃木接穗(30优,简称c)、嫁接30 a的二球悬铃木砧木(30砧,简称d)、作为庭荫树应用于居住区绿地;嫁接13 a的少果一球悬铃木接穗(13优,简称e)作为行道树应用于道路绿地。
本试验采取水培枝条萌发的新鲜叶片提取DNA,现采现提以保证DNA的质量。
1.1.2 仪器与试剂
DNA提取:采用Biomiga 公司植物DNA提取试剂盒、无水乙醇、β-巯基乙醇、无菌超纯水。
ISSR引物(上海桑尼生物科技有限公司,见表1)、dNTPs(北京索莱宝)、10×Buffer(上海博彩)、TaqDNA聚合酶(上海博彩)、D2000 DNA Ladder(TIANGEN, 后 简 称 DL2000)、琼 脂 糖( 西 班 牙 )、2×Taq PCR MasterMix(TIANGEN)等。
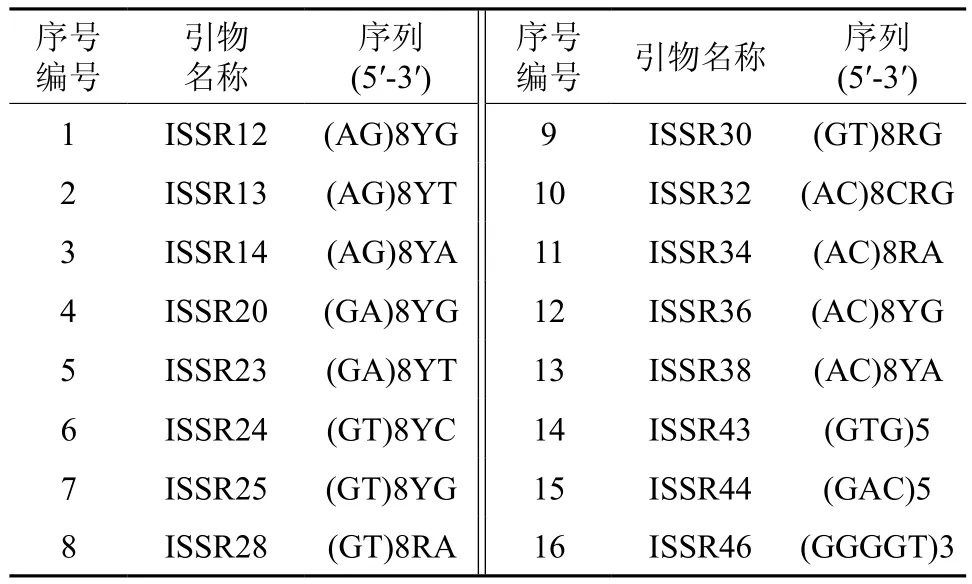
表1 ISSR引物的序号和序列Table 1 Lists of ISSR primers and their sequences in this study
仪器:冷冻离心机、电热恒温水浴锅、紫外分光光度计、DYY-6C型电泳仪、DYCP-33A型电泳槽、PCR-100TM(BIO-RAD)仪、凝胶成像系统(Gel Logic200 Imaging System)等。
1.2 方法
1.2.1 悬铃木DNA的提取
采用Biomiga 公司植物DNA提取试剂盒提取。
1.2.2 悬铃木基因组DNA质量的检测
1.2.2.1 紫外分光光度计分析
以洗脱DNA的超纯水为空白对照,从提取的DNA样品中取出2 µL,在紫外分光光度计下测定DNA在260 nm和280 nm下的吸光值,计算检测的DNA的纯度和浓度。
1.2.2.2 电泳分析
取4 µLDNA溶液点样于1%的琼脂糖凝胶,在100 V恒压条件下,电泳30 min,溴化乙锭染色10 min,在凝胶成像系统中观测、拍照。
1.2.3 ISSR-PCR反应体系筛选
以b和c两个悬铃木样本DNA为材料对3种ISSR-PCR反应体系进行筛选:
体 系 1: 20 µL 的 反 应 体 系, 包 括 5 µL GA TaqMix、0.5 µmol/L ISSR 引 物、10 ng模 板DNA。PCR扩增反应程序为:94℃预变性3 min,94 ℃变性 30 s,58 ℃复性 45 s,72 ℃延伸2 min,梯度降温(1℃/cycle)进行复性,5个循环后稳定在53 ℃复性,再进行36个循环,最后 72 ℃延伸 8 min[14]。
体系2: 20 µL的反应体系,包括终浓度为1×Buffer、1.5 mmol/L MgCl2、0.2 mmol/L dNTP、1 U Taq酶、0.5 µmol/L ISSR引 物、10 ng模 板DNA。PCR扩增反应程序同体系1。
体系3: 25 µL的反应体系,包括10 mM Tris-Hcl(pH 8.3)、1.5 mmol/L MgCl2、0.2 mmol/L dNTP、1 U Taq酶、0.2 µmol/L ISSR引 物、50~100 ng模板DNA。PCR扩增反应程序为:94℃预变性30 s,然后进行44个循环为94℃30 s,58℃ 45 s,72℃ 45 s,最后 72℃延伸 8 min[15]。
1.2.4 ISSR扩增产物检测
取 5 µLPCR 扩增产物与 1 µL Lording buffer混合,于1%的琼脂糖凝胶上,在100 V电压下,电泳150 min左右,溴化乙锭染色后检测。
1.2.5 统计分析
将5个样品重复试验中扩增清晰、重复性好、分辨度高的条带用于数据分析。以Marker为对比,按凝胶同一位置上的DNA条带有无统计,有带记为“1”,无带记为“0”,得到原始的“0”、“1”数据,构成ISSR表型数据矩阵。采用Nei等[16]的方法计算相似系数(GS) :GS=2Nij/(Ni+Nj),其中Ni和Nj分别为i和j两材料的总等位基因数,Nij为i和j两材料的共有等位基因数。遗传差异(GD)=1-GS。应用NTSYS-PC(version2.10)软件进行UPGMA聚类分析。
2 结果与分析
2.1 悬铃木基因组DNA质量的检测
2.1.1 紫外分光光度计分析
通过紫外分光光度计检测悬铃木DNA的质量,从表2中看出,5个悬铃木样品DNA的OD260/OD280值都在 1.8~2.0 之间,纯度及质量均能满足 ISSR 反应。
2.1.2 电泳分析
从图1可以看出,5个悬铃木样品的DNA质量较好,胶孔干净,无明显拖尾,且带型亮度高而整齐,这说明DNA纯度高,完整性好,适合下一步的ISSR扩增。

表2 悬铃木DNA基因组紫外分析†Table 2 Ultraviolet analysis of genomic DNA of Platanus

图1 悬铃木DNA基因组电泳检测Fig. 1 Electrophoresis detection of genomic DNA of Platanus
2.2 ISSR-PCR反应体系筛选
2.2.1 体系 1
以b(嫁接14 a的悬铃木砧木)和c(嫁接30 a的少果悬铃木接穗)为模板,对体系1进行筛选,凝胶电泳检测结果如图2和图3,结果表明:采用PCR MasterMix进行扩增,有12条引物可以扩出条带,但清晰度差、多态带少。
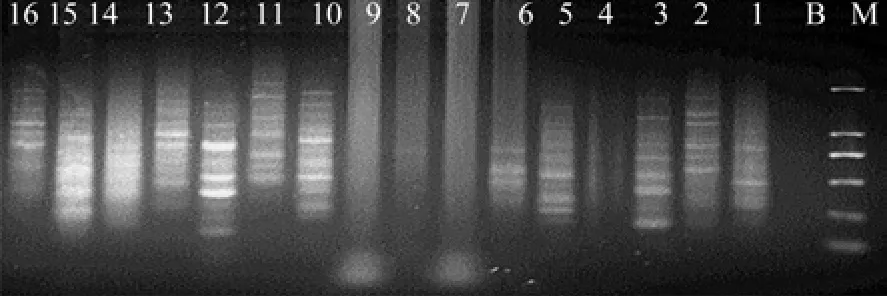
注:1-16表示引物编号,引物名称见表1,M表示DL2000分子标记

图3 14砧对反应体系1的筛选Fig.3 Screening of reaction system No.1 by rootstock grafted for 14 years
2.2.2 体系 2
以材料b和c为模板,对体系2进行筛选,凝胶电泳检测如图4和图5,结果表明,大多数引物可以扩出清晰的多态条带,因此拟用该体系进行下步PCR分析。

图4 30优对反应体系2的筛选Fig. 4 Screening of reaction system No. 2 by scion graftedfor 30 years

图5 14砧对反应体系2的筛选Fig. 5 Screening of reaction system No.2 by rootstock grafted for 14 years
2.2.3 体系3
反应体系3无条带扩出。
2.3 ‘园科115’一球悬铃木接穗的遗传稳定性
以3个‘园科115’一球悬铃木接穗(即a、c、e)为材料,用16条ISSR引物进行凝胶电泳分析,如图6所示,16条引物将3个样品一共扩增出63条清晰条带。其中,有62条条带在3个样品中呈现一致性,只有第16条引物(ISSR44)在3个样品中出现1条多态性条带,材料e(嫁接13 a的接穗)出现一条特异的亮带。由上可知,嫁接14 a与嫁接30 a的‘园科115’一球悬铃木样品对应的16条引物扩增的电泳条带完全一致,拥有极高的遗传稳定性;而材料e在引物ISSR46电泳下,在750 bp片段大小处出现多态性条带,说明嫁接13 a的‘园科115’一球悬铃木较其它两株嫁接14 a和嫁接30 a的接穗有差异,Nei's遗传相似系数为99.47%;对于3个‘园科115’一球悬铃木嫁接单株而言,群体遗传差异为0.53%。根据对枣椰[13]遗传变异的研究结果,遗传差异低于2.6%被认为遗传稳定。‘园科115’一球悬铃木嫁接年限分别为13 a、14 a和30 a的3个接穗,虽然处于不同的立地环境条件,但是其遗传差异仅仅为0.53%,遗传非常稳定,能保持亲本的优良遗传性状。

图6 3个少果毛悬铃木样品凝胶电泳图像(箭头示多态性条态)Fig. 6 Gel electrophoresis image of three scion samples(arrows indicate polymorphic bands)
2.4 ‘园科115’一球悬铃木接穗与对应的砧木间亲缘关系
2.4.1 5个悬铃木样品ISSR遗传多样性分析
由图7和8可知,以16条引物进行扩增,5个悬铃木样品之间存在多态性条带。如表3,以16条引物共扩出75条条带,平均每条引物扩出4.687 5条,其中多态性条带27条,多态条带百分率为36%。扩增条带片断大小在250~2 000 bp之间。由此可知,5个样品间存在丰富的遗传多样性。其中12、13、16号引物多态性条带比率较高,都在60%以上。

图7 嫁接14 a‘园科115’一球悬铃木与对应砧木凝胶图像Fig. 7 Gel electrophoresis image of grafted 14 years plus tree and rootstock

图8 嫁接30a‘园科115’一球悬铃木与对应砧木凝胶图像Fig.8 Gel electrophoresis image of grafted 30 years plus tree and root stock

表3 检测5个悬铃木样品的遗传相关性ISSR引物列表Table 3 ISSR primers list of 5 Platanu tested samples’Genetic eovariance
2.4.2 5个悬铃木样品遗传相似度与聚类分析
从表4可以看出,5个悬铃木样品间的遗传相似系数在0.680 0~1.00 0之间,其中14优与30优之间无差异,而这两个悬铃木样品与13优的遗传相似系数均为0.986 7,相似度很高;14优与14砧、30优与30砧遗传相似系数均为0.800 0。
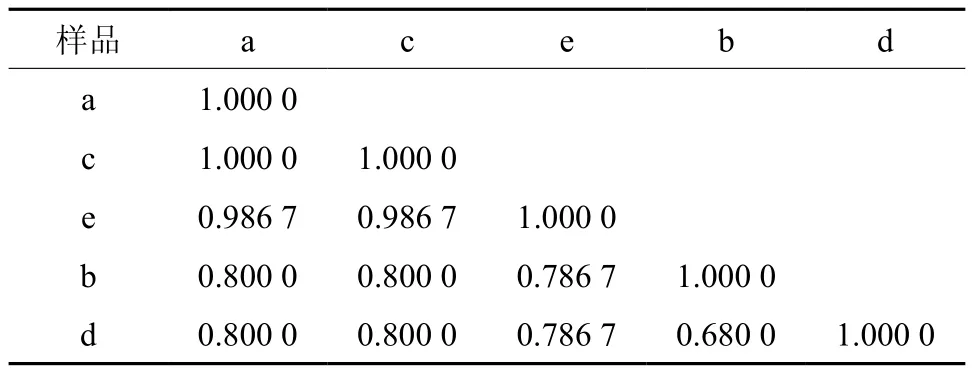
表4 5个悬铃木样品种间相似系数†Table 4 Genetic similarity coefficients among 5 Platanus samples
基于遗传相似系数的聚类分析显示(图9),14优与30优间无差异;而遗传相似系数在0.98~1.0以上可以把14优、30优与13优3个单株归为一类,说明3个‘园科115’一球悬铃木嫁接单株遗传相似度极高,结论与上述(2.3)相同。遗传相似系数在0.80左右时,可以将3个‘园科115’一球悬铃木接穗与14砧划分为一大类;而在遗传相似系数0.77左右将5个悬铃木样品划分为一大类。

图9 5个悬铃木样品的ISSR聚类分析树Fig.9 Dendrogram for 5 Platanus by cluster analysis(UPGMA) based on ISSR marker
综上所述,经过长达14 a及30 a的嫁接时间,‘园科115’一球悬铃木接穗与砧木之间遗传差异显著,而3个‘园科115’一球悬铃木嫁接单株之间遗传差异极小,由此可说明该少果毛品种——‘园科115’一球悬铃木遗传稳定性高,多年嫁接后仍与母本性状保持一致。3个‘园科115’一球悬铃木材料分别嫁接于二球悬铃木应用于单位绿地、居住区绿地和道路绿地,因此也可以说明不同的立地条件对该少果毛悬铃木品种的遗传特性影响较小。
3 讨 论
本试验通过对比3种PCR体系,确定了‘园科115’一球悬铃木最佳ISSR反应体系,即:选用20 µL的反应体系,包括终浓度为1×Buffer、1.5 mmol/L MgCl2、0.2 mmol/L dNTP、1 U Taq 酶、0.5 µmol/L ISSR引物、10 ng模板DNA。PCR扩增反应程序为:94℃预变性3 min,94 ℃变性30 s,58 ℃复性45 s,72 ℃延伸2 min,梯度降温(1℃/cycle)进行复性,5个循环后稳定在53 ℃复性,再进行36个循环,最后72 ℃延伸8 min。
嫁接14 a及30 a后,2个‘园科115’一球悬铃木接穗与对应砧木仍存在着较大的遗传差异,其中嫁接14 a‘园科115’一球悬铃木接穗与砧木、嫁接30 a‘园科115’一球悬铃木接穗与砧木遗传相似系数均为0.800 0,说明嫁接砧木与‘园科115’一球悬铃木的间遗传差异显著。以嫁接13 a、14 a及30 a的3个‘园科115’一球悬铃木为材料,通过16条ISSR引物凝胶电泳分析,认为3个嫁接单株遗传相似度极高。以上结果说明嫁接多年后,该少果毛品种与母本仍保持一致,遗传性状稳定,可以进行大面积推广。
本研究采用可靠的ISSR分子标记,发现嫁接30 a与14 a的‘园科115’一球悬铃木无任何遗传变异。嫁接13 a的‘园科115’一球悬铃木与嫁接14 a和30 a的单株也仅仅有1个位点的变异,还有待于进一步验证。可以说:各种立地环境条件和嫁接时间不改变‘园科115’一球悬铃木的遗传稳定性。砧木也对接穗的遗传组成无任何影响。
[1] 曾幼佛,黄正华,祁德胜,等. 悬铃木少毛栽培管理措施的调查研究[J]. 中国园林,1985,(3):21-23.
[2] Zietkiewicz E, Rafalski A, Labuda D. Genome fingerprinting by simple sequence repeat (SSR)-anchored polymerase chain reaction amplif i cation[J]. Genomics, 1994, (20):176-183.
[3] 刘小莉,刘飞虎,李宗菊,等. 10种报春花亲缘关系的ISSR分析[J].云南大学学报:自然科学版,2004,26(5):454-458.
[4] 李 萍,石金磊,胡永红,等. 凤梨亚科光萼荷属与其近源属亲缘关系的ISSR分子鉴定[J].种子,2007,26(11):35-40.
[5] Rout G R, Senapati S K, Aparajita S. Studies on the genetic relationship among 13 cultivars of Calathea (Marantaceae) using RAPD and ISSR markers[J]. Advances in Horticultural Science,2007, 21(3):147-155.
[6] 刘海龙,马锦林,张日清,等.11份山茶属植物亲缘关系的ISSR分析[J]. 经济林研究, 2012,30(4):87-90.
[7] 王 利,陈 述, 董金伟,等. 古银杏种质亲缘关系及遗传多样性的ISSR分析[J]. 经济林研究, 2010,28(3):20-24.
[8] 杨淑达,施苏华,龚 洵,等.滇牡丹遗传多样性的ISSR分析[J].生物多样性,2005,13(2):105-111.
[9] Rzepka-Plevneš D, Smolik M, Tańska K. Genetic similarity of chosen Syringa species determined by the ISSR-PCR technique[J]. Dendrobiology, 2006,(56):61-67.
[10] 王晓明,谢碧霞,李俊彬,等.金银花ISSR分子标记及遗传多样性分析[J]. 中南林业科技大学学报, 2008, 28(6):14-18.
[11] 李芳东,袁德义,莫文娟,等.白花泡桐种源遗传多样性的ISSR分析[J]. 中南林业科技大学学报, 2011,31(7):1-7.
[12] 邱英雄,傅承新,何云芳.乐昌含笑不同类型鉴定的 ISSRPCR分析[J].林业科学,2002,38(6):49-52.
[13] 沈永宝,施季森,赵洪亮.利用 ISSR DNA 标记鉴定主要银杏栽培品种[J].林业科学,2005,41(1):202-204.
[14] 黄文俊.农杆菌介导不育基因转化悬铃木及长期继代培养植株的遗传稳定性分析的研究[D].武汉:华中农业大学,2007.
[15] 王艳梅,景自景,刘 震,等.悬铃木ISSR反应体系的建立[J].山东农业大学学报:自然科学版, 2011,42(1):30-34.
[16] Nei M, Li W H. Mathematical model for studying genetic variation in terms of restriction endonucleases[J]. Proc Natl Acad Sci USA, 1979,76(10):5269-5273.
[17] Saker M M, Adawy S S, Mohamed A A, et al. Monitoring of cultivar identity in tissue culture-derived date palms using RAPD and AFLP analysis[J]. Biol Plant, 2006,50(2):198-204.
Analysis on less fruit hair Platanus occidentalis genetic stability based on ISSR marker
YAO Jun1, ZHANG Li-fang2, TU Bing-kun2, DING Zhao-quan1
(1. Wuhan Institute of Landscape Gardening, Wuhan 430081, Hubei, China; 2. Key Lab. of Horticultural Plant Biology Supported by China Ministry of Education, College of Horticulture and Forestry, Huazhong Agricultural University, Wuhan 430070, Hubei, China)
The analyses of genetic stability were performed on the 3 scions of Platanus occidentalis‘Yuanke 115’in different site conditions with different grafting time (13, 14 and 30 years respectively) and 2 corresponding rootstocks based on ISSR molecular marker. There were 75 bands in total amplified with 16 ISSR primers in 5 samples, of which 27 bands were polymorphic, and the polymorphic percentage was 36%. According to ISSR markers data, the cluster analysis based on genetic similarity coeff i cients was conducted. The results are as follows: The genetic similarity coeff i cients between 5 samples ranged from 0.6800 to 1.0000; 5 samples were grouped into one category with the genetic similarity coeff i cient of 0.77; The 3 scions were grouped into one category with the genetic similarity coeff i cient of 0.98; The genetic similarity coeff i cient between scions with 14 years and 30 years grafting time was 1.0000; Either of the scions with 14 years and 30 years grafting time had a genetic similarity coeff i cient of 0.9867 with the scion with 13 years grafting time. The results above show that P. occidentalis‘Yuanke 115’confers high level of genetic stability in different site conditions and through different period of grafting time (up to 30 years).
Platanus occidentalis‘Yuanke 115’; molecular makers; genetic stability; ISSR molecular marker
S722.3+1;S687.1
A
1673-923X(2014)02-0006-06
2013-05-15
武汉市园林局科研项目(武园[2009]55号)
姚 军(1977-),男,湖北荆门人,高级工程师,主要从事园林植物繁育与应用研究;E-mail:yaojun77@gmail.com
涂炳坤(1952-),男,湖北罗田人,教授,博士生导师,主要从事林木良种选育、林木生理生态、工业原料林定向栽培、经济林栽培生理生化研究;E-mail:bktu@mail.hzau.edu.cn
[本文编校:文凤鸣]
