牧草分子遗传连锁图谱及其应用
2014-12-24谢文刚刘文献张建全王彦荣
谢文刚,刘文献,张建全,王彦荣
(草地农业生态系统国家重点实验室,兰州大学草地农业科技学院,甘肃 兰州730020)
牧草,广义上泛指可用于饲喂家畜的草类植物,包括草本、藤本、小灌木、半灌木和灌木等各类栽培或野生植物;狭义上仅指可供栽培的饲用草本植物,尤指豆科牧草和禾本科牧草[1]。牧草是农业自然资源的重要组成部分,是农业可持续发展的重要生产资料,也是发展草地畜牧业的重要物质基础。培育高产、优质、高抗的牧草新品种,充分发挥其在改良退化农田、恢复草地功能、调整农业结构、固碳减排、建立草地农业系统中不可替代的作用,是保障国家食物安全和生态安全的重要措施之一。
遗传连锁图谱是指通过遗传重组分析得到的基因或专一的多态性遗传标记在染色体上的线性排列顺序 图[2]。目前包括玉米(Zeamays)[3]、水稻(Oryzasativa)[4]、小麦(Triticumaestivum)[5]等主要农作物均已构建了遗传连锁图谱,并被广泛应用于数量性状位点(QTL)定位[6]、比较基因组[7-8]和分子标记辅助育种[9-10]等领域以改良植物的重要农艺性状,大幅度提高植物的育种水平和育种效率。和大多数农作物相似,牧草许多重要的农艺性状如产量、品质、抗性等一般为数量性状,受微效多基因控制。牧草常规育种周期长,效率低,而利用DNA分子标记构建牧草遗传图谱,开展QTL定位研究及分子标记辅助育种,加快培育高产、优质、高抗牧草新品种已成为目前牧草育种工作中的一个重要发展方向。本文就牧草遗传连锁图谱构建及应用的研究现状和最新成果进行了概述,以期为牧草分子育种的进一步发展提供参考。
1 牧草遗传连锁图谱的构建研究现状
1.1 已构建的遗传连锁图谱
与多数一年生模式植物相比,牧草多为多年生、异交的多倍体植物,如紫花苜蓿、白三叶、鸭茅等同源四倍体,而羊草(Leymuschinensis)为异源四倍体,遗传背景相对复杂,异交多倍体牧草存在分离基因型众多、不同DNA片段共分离,杂交后代常表现为四体遗传等特点,造成其在遗传分析上存在较多困难。但随着“拟测交”策略的应用[11]及各种分子标记技术的不断发展,据不完全统计,目前已构建了20余个草种近40张遗传连锁图谱(表1)。所构图谱主要涉及豆科的苜蓿属(特别是紫花苜蓿)[12-18]、三叶草属[19-21]、百脉根[22-23],禾本科的羊茅属[24-26]、黑麦草属[27-34]和鸭茅属[35-37]等常见草种。
1.2 作图群体
构图群体亲本的选择是群体构建成功及图谱质量高低的决定因素之一,最大限度地选择亲本间性状差异与亲和性的统一是亲本选择的基本原则[2]。常见的遗传连锁图谱的群体有:临时性群体如F2及其衍生的F3、F4家系和回交群体(BC)。永久性分离群体,如回交自交系群体(BIL)、重组自交系群体(RIL)、双单倍体群体(DH)、近等基因系群体。对于亲本基因型高度杂合的植物,也可采用“双拟测交”(Double-pseudo-testcross)的方法建立F1分离群体[11]。“双拟测交”是指两个亲本基因型均为杂合,互为测交群体,与测交试验一样,后代基因型的分离比例为1∶1,但两个亲本都非纯合隐性,所以称为拟测交。由于大多数牧草具有多年生、异花授粉和自交不亲和等特性,使培育纯系较为困难,难以构建BC1等群体,因此,目前大部分牧草分子连锁图谱大部分是利用“双拟测交”F1作图群体构建的,如多年生黑麦草[28-30]、多花黑麦草[32-33]、草地羊茅[26]、鸭茅[35-37]和赖草[38]等。对于自交衰退不严重的异交草种,也可仿照一年生农作物创建自交纯系进行杂交以获得F2群体或BC1群体进行遗传作图,如结缕草(Zopsiaspp.)[41]、紫花苜蓿[12,14,16]等。
另外,作图群体的大小也是影响图谱品质的重要因素。一般而言,大的作图群体包含更多的基因重组个体,在遗传作图时能获得更多的分离标记,使所构图谱更饱满,基因组覆盖更全面。如 Mott等[39]利用含387个后代的回交群体,构建了含515个分子标记,图幅长度为2 574cM的遗传图谱,标记间的平均距离为4.9cM。
1.3 作图标记
DNA分子标记直接反映植物分子水平上的遗传多态性,因其简单、快捷、受环境影响小和可重复性强等优点,被广泛应用于牧草遗传图谱构建研究中。目前,用于植物遗传作图的主要分子标记有RFLP、RAPD、AFLP、SRAP和SSR等。
对于大多数牧草物种而言,亲本是高度杂合的个体,从亲本到子代需要传递多个等位基因,RAPD等显性标记往往难以区分纯合基因型或杂合基因型,而SSR等共显性标记能够检测多个等位位点,获得异交亲本中最大量的多态性。早期构建的遗传连锁图谱主要利用的分子标记是RFLP和AFLP等。RFLP标记具有重复性好和共显性的特点,在构建牧草核心或框架图谱,特别是比较作图中有重要作用。如Xu等[24]利用RFLP标记构建了高羊茅的遗传图谱,构图群体中有105个子代,图谱长度达1 274cM,分布在19个连锁群。该标记也被用在紫花苜蓿[12-14,16]、多年生黑麦草[28,30,32]等草种的遗传图谱构建上。AFLP技术耗时量小,信息量大,是构建高精密度分子遗传图谱的重要标记。如Sandal等[23]在日本百脉根的研究中利用AFLP所构建的图谱中图幅的平均距离已小到0.6cM。SSR标记由于具有共显性、基因组中分布广泛、重复性好、多态性高、信息量大等优点,避免了RFLP方法中使用放射性同位素的缺点,又比RAPD重复率和可信度高,在遗传图谱构建方面具有广阔的研究前景,是理想的标记之一,已应用于紫花苜蓿[17]、红三叶[20]、高羊茅[25]、黑麦草[27,29]、鸭茅[35-37]、赖草[38]、结缕草[42,44]等草种的遗传图谱构建。随着高通量测序等技术的发展,单核苷酸多态性标记(Single Nucleotide Polymorphism,SNP)、序列标签位点(Sequence tag Site,STS)标记[39]及多样性微阵列技术(Diversity Arrays Techenology,DArT)[31]等新型标记也将在遗传作图中显示出广泛的应用前景。
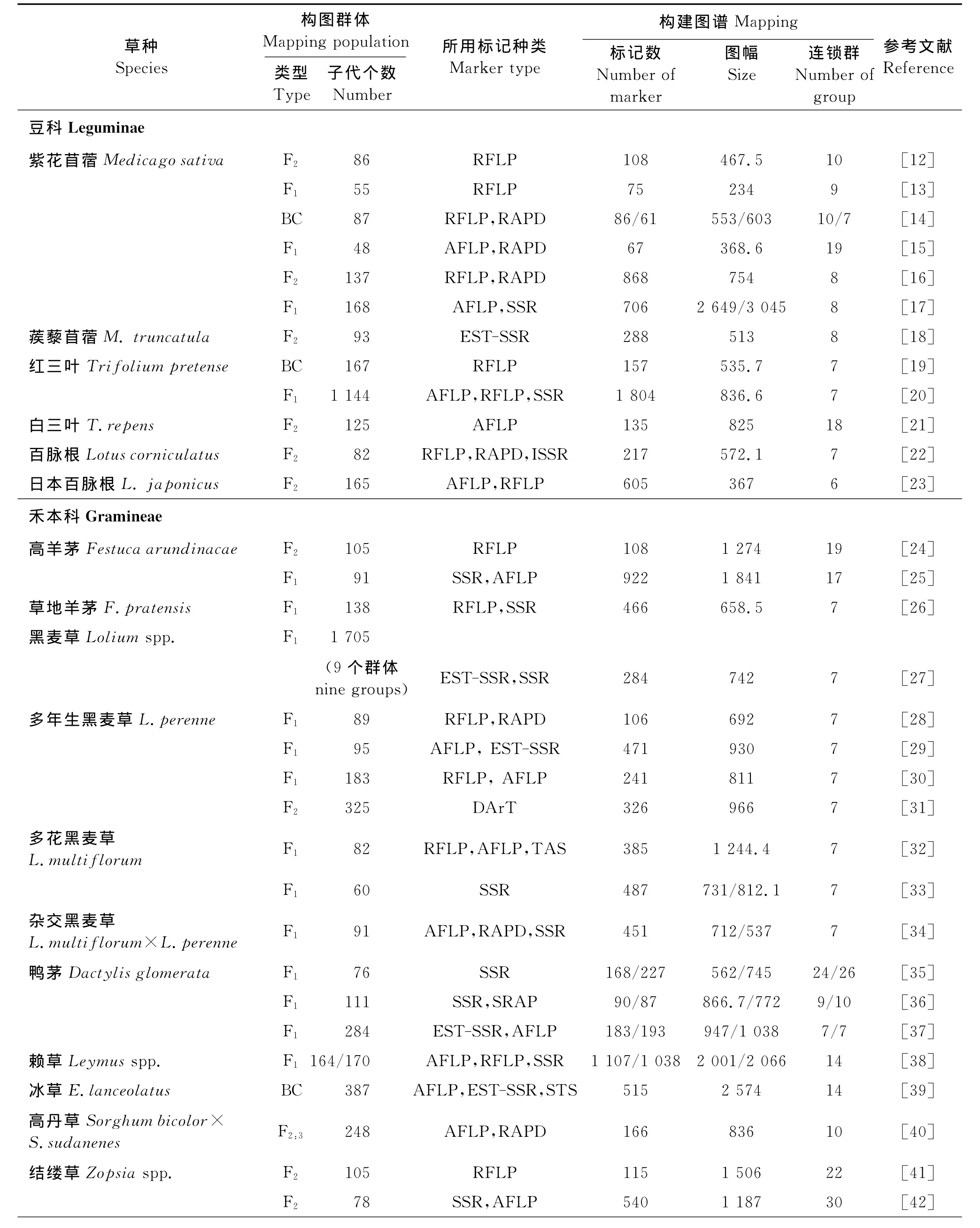
表1 牧草遗传图谱构建研究进展Table 1 Genetic linkage maps in forage grasses

续表1
2 牧草遗传连锁图谱的应用
对于牧草育种者而言,牧草大多数的性状如干物质产量、牧草品质和非生物抗性等常表现出连续的表型变异,并受多个数量性状位点控制。以往育种者主要是通过多年轮回选择对目标性状进行改良,育种周期长,效率低。分子遗传连锁图谱则有助于在分子层面分析与主要农艺性状相关联的基因区域或分子标记,从而定位和克隆重要的农艺性状基因、检测和标记数量性状位点,并应用于分子标记辅助育种,可大幅度提高牧草育种的水平和效率。
2.1 牧草重要农艺性状QTL定位
随着分子标记技术和分子连锁图谱构建的迅猛发展,使得数量性状位点(QTL)定位成为可能。目前国外已有大量的牧草QTL研究的报道(表2),主要集中在黑麦草属[27,50-70]、赖草属[73-77]和苜蓿属[78-84]等。我国牧草QTL研究的报道相对较少,仅见于鸭茅[37]、高丹草[40]、结缕草[90]等少数草种。在牧草QTL研究领域,黑麦草属QTL研究最为广泛和深入,研究内容涉及多个主要的农艺性状,如抗病性[50-56]、开花期[60-63]、种子产量相关性状[58]、牧草品质相关性状[64-65]、牧草产量相关性状[67]等。
2.1.1 抗病QTL定位 牧草抗病QTL研究主要集中在黑麦草属上,包括抗锈病、细菌性萎蔫病、霉粉病和叶斑病等。Studer等[27]用含307单株的“双拟测交”群体为材料对多花黑麦草冠锈病QTL进行了研究,在第1和2连锁群上检测到抗冠锈病相关QTL,它们能解释高达56%的表型变异。在多年生黑麦草第1和2连锁群上也发现了4个冠锈病QTLs,其中LpPc4和LpPc2位于第1连锁群,Lp-Pc3和LpPc1位于第2个连锁群上,它们分别能解释12.5%、24.9%、5.5%和2.6%的表型变异,比较作图分析发现,这些QTL所在连锁群与燕麦A和B染色体具有同源性,在燕麦A和B染色体上已经鉴定了抗冠锈病相关基因[52]。Pfender等[54]也在第1、6和7连锁群上共检测到3个抗杆锈病QTL(qLpPg1、qLpPg2、qLpPg3),其中主效 QTL(qLpPg1)定位在第7连锁群上,可解释30%~38%的表型变异。
2.1.2 产量相关QTL定位 提高牧草单位面积草产量或种子产量是牧草的主要育种目标,其中种子产量对牧草品种的生产、推广和应用有重要影响。Espinoza等[84]利用蒺藜苜蓿(Medicagotruncatula)4个作图群体研究了主枝长度(LPB)、分枝延伸率(BER)、干物质(ADM)、主茎干长(LMS)等产量相关性状,通过对单个和多个群体的分析表明,主要的QTL区域位于第1、2、7和8染色体上,多群体分析揭示了3个与LPB、LMS、BER等性状密切相关的主效的QTL区域。Studer等[58]利用F2作图群体,构建了黑麦草遗传连锁图谱,在连锁群1和2上检测到两个与种子产量相关的QTL,它们分别解释了41%和18%的表型变异。Faville等[67]在多年生黑麦草连锁群1、2、4和6上检测到与草产量有关的QTL。Herrmann等[86]检测到38个与红三叶种子产量性状相关(每株植物的种子数、每个花序上的种子产量、每株植物上的花序数等)的QTL,这为红三叶种子产量性状的改良和分子标记辅助育种奠定基础。Robins和Brammer[80]利用紫花苜蓿F1群体对影响牧草产量、株高、再生性等产量相关性状进行了QTL分析,分别在连锁群3、4、5和7上检测到影响这些性状的QTL位点,它们解释的表型变异在11%~44%。

表2 牧草主要农艺性状QTLTable 2 QTL for important agronomic trait in forage grasses
2.1.3 牧草品质相关QTL定位 Espinoza和Julier[83]利用蒺藜苜蓿作图群体研究了粗蛋白、消化率等品质相关的QTL,主效QTL主要分布于染色体1、3、7和8的4个基因区域,其中牧草品质相关的QTL和形态QTL定位在相同的区域,由此推测品质性状和表型特征具有紧密的关系,该研究为进一步鉴定和挖掘豆科牧草品质相关的基因奠定了基础。Cogan等[64]对黑麦草牧草品质(如粗蛋白、体外消化率、中性洗涤纤维等)的QTL进行了分析,在连锁群3、5、7上共检测到42个影响这些性状的QTL位点,其中LG3包含了影响所观测性状的所有QTL位点,LG7上包含了除粗蛋白外的所有QTL位点。研究指出,与这些位点紧密连锁的分子标记,对于分子标记辅助育种有重要利用价值。碳水化合物含量(WSC)是决定牧草品质的一个重要因素,对该性状进行分子标记辅助选择对于黑麦草分子育种及目标性状的改良意义重大。Turner等[65]检测到影响黑麦草 WSC含量的QTL,同时利用与WSC含量QTL紧密连锁的标记去测试这些QTL的效应,虽然WSC含量和其他因素有复杂的相互影响,但进一步证明了这些QTL区域对黑麦草WSC含量有调控作用。另外,黑麦草脂肪酸成分QTL也被定位于相应的连锁群上,其中棕榈酸含量(C16:0)QTL位于连锁群2和7、硬脂酸(C18:3n-3)位于连锁群3、4和7、亚油酸(C18:2n-6)和亚麻酸(C18:3n-3)分别位于连锁群2和1[69]。Larson和 Mayland[73]对赖草粗蛋白(CP)、中性洗涤纤维(NDF)、酸性洗涤纤维(ADF)和 Al、B、Ca、Cu等14种矿物质含量及 K/(Ca+Mg)比例(KRAT)等进行了分析,发现在作图群体中这些性状存在显著的变异,并检测到所有性状的QTL位点,如 ADF位于 LG1a、LG5Xm、LG7a、NDF位于LG7a,Mn位于 LG2b、LG3b、LG4Xm,S位于 LG3等。研究结果对于筛选和培育新的赖草新品种,鉴定和挖掘影响矿物质含量及纤维互作的功能基因也有重要意义。
2.1.4 抗非生物胁迫QTL定位 水涝通过影响植物根部对水分的吸收从而影响植物正常生长。Pearson等[68]利用NAx和AU6为亲本构建了F1作图群体,在对作图群体进行8d水涝胁迫后,测定了9个相关的数量性状,最终检测到37个QTL位点。其中19个QTL位点位于NAx图谱上,18个位于AU6图谱上。在连锁群3和4上发现了黑麦草耐涝相关的特殊区域。在抗冻性和抗旱性研究方面,Alm等[71]在草地羊茅染色体5F上检测到两个抗冻性QTL,在染色体3F上检测到1个抗旱性QTL。Brouwer等[79]利用紫花苜蓿为材料定位了冬季抗寒、抗冻和秋季生长性QTL,在第5和8连锁群上发现了影响秋季生长,冬季抗寒、抗冻的QTL,但仅对秋季生长和抗冻性有影响的QTL定位在连锁群1和3,这些QTL区域可用于分子标记辅助育种选择。Sledge等[81]对二倍体紫花苜蓿抗铝性进行了QTL研究,发现两个RFLP标记UGAc471和UGAc502与作图F2群体的抗铝性紧密连锁,可用于将检测到的QTL位点渗入到栽培的紫花苜蓿品种以提高其抗铝性。丁成龙等[90]在人工低温胁迫条件下研究结缕草叶片的半致死温度(LT50)、可溶性糖、可溶性蛋白含量及过氧化物歧化酶(SOD)活性的变化,并以447个SSR标记构建的日本结缕草遗传连锁图谱为基础,对抗寒相关的性状可溶性糖、可溶性蛋白含量及SOD活性进行了QTL定位分析,分别定位到与叶片可溶性糖、可溶性蛋白含量和SOD活性相关的QTL各1个,分布于连锁群10、13和17。
2.1.5 繁殖性状相关QTL 了解植物的繁殖特性与种子生产性能是种质资源的保存与更新、野生种的驯化选育和新品种推广利用的重要前提。深入研究草类植物繁殖性状的生物学基础,培育种子产量高、生产性能好的草类新品种,是以往国内外关注不多、但目前亟待研究的重要领域。草类植物的繁殖性状,主要包括开花时间、花部形态、种子育性、种子落粒性、种子形态等。Xie等[37]在鸭茅连锁群2、5和6检测到与开花时间相关的7个QTL。其中LG2上的QTL定位在两亲本同源连锁群相同区域,且都与标记Contig3046紧密连锁,QTL分别解释了8%和22%的表型变异。该研究所定位的QTL位置与近缘物种如黑麦草等所定位的开花QTL位置基本一致,为近一步挖掘鸭茅开花基因奠定了良好基础。Larson和Kellogg[74]在赖草属杂交种第6条染色体检测到控制落粒性的QTL,该位点与水稻第2条染色体具有同源性。Ergon等[72]在草地羊茅染色体7F上检测到一个影响小穗数量变异的基因区域。一个穗长QTL、3个小穗数QTL被鉴定,分别解释了32%~33%的表型变异。并且这些QTL与抽穗期QTL基本一致,表明他们受潜在的相关基因协作调节。在连锁群4的顶端发现一个区域同时包含了抽穗时间、穗长和小穗数QTL,同时这也是穗长主效QTL所在位置。
2.2 重要农艺性状基因定位
从目前牧草研究情况看,在黑麦草、羊茅等少数牧草上有开花基因和抗逆基因的相关报道。开花期是牧草的重要农艺性状,对牧草产量、饲用品质及利用价值有重要影响。抽穗开花后饲草品质往往快速下降,生产上应根据不同需要培育不同生育期的早晚熟品种,尤其在混播草地中培育晚熟的牧草品种能大大提高草地生产能力和利用效率。Jensen等[63]利用多年生黑麦草品种“Veyo”和来自于“Falster”的一个基因型杂交,构建了含184个单株的F2的作图群体,检测到5个影响黑麦草春化的QTL,并在第4连锁群上主效QTL的附近成功定位了一个春花基因VRN1。Xie等[92]利用水稻、小麦、拟南芥(Arabidopsisthaliana)、黑麦草等物种的开花期基因的保守序列,设计基因探针,与鸭茅基因组杂交进行目标基因的捕获,通过测序并设计基因引物最终将Vrn3基因成功定位在鸭茅的第3个连锁群上。Armstead等[60]在多年生黑麦草上鉴定了与水稻Hd1基因和大麦(Hordeumvulgare)HvCOl基因同源的抽穗基因LpHd1,该基因位于黑麦草第7染色体的定位区域。Tamura和Yamada[93]定位了多年生黑麦草的CBF基因簇,4个CBF基因(LpCBFIb,LpCBFII,LpCBFIIIb和LpCBIIIc)定位在连锁群5上,另一个基因LpCBFVb定位于连锁群1上。
2.3 比较基因组分析
比较作图就是利用共同的遗传标记(主要是分子标记、基因的cDNA克隆以及基因组克隆)对相关物种进行物理或遗传作图,比较这些标记在不同物种基因组的分布情况,揭示染色体片段上的同线性(Synteny)或共线性(Collinearity),从而对不同物种的基因组结构及基因组进化历程进行准确分析[94]。比较作图的分子基础是物种间DNA序列尤其是编码序列的保守性。植物的比较作图最早是在番茄(Lycopersiconesculentum)和马铃薯(Solanum tuberosum)[95]、番茄和胡椒(Pipernigrum)[96]之间进行的。近年来,比较作图在禾本科作物分子作图上迅速发展。通过黑麦(Secalecereale)、玉米、小麦和水稻[97]、玉米和高粱(Sorghumbicolor)等[98]的比较基因组研究表明,虽然这些物种在遗传背景、染色体组成、基因组大小存在差异,但比较作图结果表明:许多标记在不同作物的遗传图谱上的位置和顺序都具有高度的保守性,说明染色体间共线性片段和基因组的同源性在不同物种间广泛存在。开展牧草与作物间的比较基因作图研究,对于在基因组研究较少的牧草物种中发现重要的农艺性状基因,开展功能分析具有重要意义。
比较基因作图和微线性分析表明,黑麦草抽穗基因LpHd1和草地羊茅抽穗基因FpHd1是水稻Hd1基因的同源基因。表明LpHd1和FpHd1基因预测的蛋白质序列是类似CONSTANS基因锌指蛋白,它们与水稻Hd1的序列同源性分别为61%~62%,而与大麦HvCOl的序列同源性为72%。同时LpHd1基因在黑麦草第7染色体的定位区域,与水稻Hd1基因所在的第6染色体和大麦HvCOl基因所在的第7H染色体的基因组间具有一定的共线性[60]。此外,多花黑麦草抽穗期的主效QTL与水稻抽穗期基因Hd3位点具有共线性关系[62]。而黑麦草上已发现3个与春化反应相关的候选基因,DNA序列比对表明它们与二倍体小麦的TmVRN2基因及水稻的Hd1基因具有高度的同源性[99]。Alm等[71]对草地羊茅抗冻性和抗旱性 QTL进行了定位,通过将草地羊茅两个抗冻QTL小麦相关QTL进行比较分析发现,羊茅染色体5F上的两个抗冻QTL与小麦同源连锁群5A上的Fr-A1和Fr-A2具有较高同源性,草地羊茅3F上的抗旱性QTL与水稻染色体上相应的抗旱位点具有线性关系。对鸭茅EST序列和黑麦草EST序列的同源比对发现:鸭茅连锁图谱上的8个SSR标记与黑麦草连锁图谱上的标记具有同源性。其中,鸭茅连锁群1、2、3、4的部分SSR标记与黑麦草相同连锁群上的标记序列具有同源性。而鸭茅连锁群5的SSR标记与黑麦草第7连锁群的EST序列具有同源性[37]。
3 牧草遗传连锁图谱构建存在问题及展望
遗传图谱的构建开创了牧草分子遗传育种的新局面,但现阶段直接用于牧草的育种实践还有很大的距离。主要表现在以下方面:
(1)作图标记以随机标记为主,共显性标记利用有限。目前,一半以上牧草连锁图谱是用RAPD和AFLP等显性标记构建而成的。虽然利用这些随机标记可获得大量分离位点,快速构建连锁图谱,但它们无法准确区分纯合子和杂合子,且这些标记扩增的片段通常是非编码区域,难以直接用于分离控制重要农艺性状的基因,造成在牧草分子育种及目标性状改良上有很大的局限性,极大地降低了遗传图谱的应用价值。因此,在遗传标记选择上,应多选用SSR、EST-SSR等共显性标记,同时利用近缘物种上成功构图且与重要农艺性状基因或QTL连锁的标记联合作图,以大大提高图谱的饱和度,同时利于更准确地进行QTL分析及开展不同物种间比较作图分析。
(2)牧草遗传图谱构图群体选择和群体大小。为保证连锁图谱的准确性,每个群体应保证有足够的个体,而构建骨架连锁图可基于大群体中的一个随机小群体(如150个单株或家系)[100]。在已构建的牧草连锁图谱中构图群体大小从几十到几百不等,仅有红三叶[19]、日本百脉根[23]、多年生黑麦草[30]、鸭茅[37]、赖草[38]和冰草[39]等草种的部分作图群体大于150个单株。但几乎所有的构图群体是F1、F2暂时性分离群体,虽然这些群体构建时间短、成本低、信息量大,但难以较长期保存,研究结果的可比性不强,不利于信息的共享。在今后的工作中应加强多子代永久群体的构建和保存研究,发展基于更大的作图群体下建立起来的植物遗传连锁图谱。
(3)构建的牧草连锁图谱大多密度偏低,且均匀性较差,难以满足基因定位、克隆和分子辅助选择育种的要求。已构建的牧草连锁图谱大多是框架图谱,连锁群数与研究对象实际染色体数或多或少存在差异,不同构图群体和不同亲本间的连锁群有时也不同;大多数已构建的牧草图谱的图幅较小,标记间的间隙较大,还不能满足QTL分析、基因克隆和分子标记辅助选择对遗传图谱的要求。加强准确、完整、实用和高密度遗传图谱的构建研究十分必要。
总之,植物遗传连锁图谱的发展趋势是高饱和化、实用化和通用化。随着人们对牧草重要性认识的不断提高及分子生物学技术的持续发展,牧草遗传连锁图谱构建的研究必将迈上一个新的台阶。在此基础上结合牧草育种的主要目标开展高产、优质、抗逆等重要农艺性状QTL定位研究,挖掘和克隆一批重要的功能基因,以改良牧草重要的农艺性状,加快牧草育种进程,是牧草育种的必由之路。
[1]董宽虎,沈益新.饲草生产学[M].北京:中国农业出版社,2003.
[2]方程,韩建国.牧草遗传连锁图谱构建研究概述[J].草地学报,2006,14(3):287-291.
[3]Castiglioni P,Ajmone-Marsan P,Van Wijk R,Motto M.AFLP markers in a molecular linkage map of maize:Codominant scoring and linkage group distribution[J].Theoretical and Applied Genetics,1999,99(3-4):425-431.
[4]Price A H,Steele K A,Moore B S,Barraclough P P,Clark L J.A combined RFLP and AFLP linkage map of upland rice(OryzasativaL.)used to identify QTLs for root-penetration ability[J].Theoretical and Applied Genetics,2000,100(1):49-56.
[5]Paillard S,Schnurbusch T,Winzeler M,Messmer M,Sourdille P,Abderhalden O,Keller B,Schachermayr G.An integrative genetic linkage map of winter wheat(TriticumaestivumL.)[J].Theoretical and Applied Genetics,2003,107:1235-1242.
[6]Song X J,Huang W,Shi M,Zhu M Z,Lin H X.A QTL for rice grain width and weight encodes a previously unknown RING-type E3ubiquitin ligase[J].Nature Genetics,2007,39(5):623-630.
[7]Kato K,Nakamura W,Tabiki T,Miura H,Sawada S.Detection of loci controlling seed dormancy on group chromosomes of wheat and comparative mapping with rice and barley genomes[J].Theoretical and Applied Genetics,2001,102:980-985.
[8]Varshney RV,Sigmund R,Börner A,Korzun V,Stein N,Sorrells M E,Langridge P,Graner A.Interspecific transferability and comparative mapping of barley EST-SSR markers in wheat,rye and rice[J].Plant Science,2005,168,195-202.
[9]Zhou W C,Kolb F L,Bai G H,Domier L L,Boze L K,Smith N J.Validation of a major QTL for scab resistance with SSR markers and used of marker-assisted selection in wheat[J].Plant Breeding,2008,122:40-46.
[10]Jena K K,Mackill D J.Molecular markers and their use in marker-assisted selection in rice[J].Crop Science,2008,48(4):1266-1276.
[11]Grattapaglia D,Sederoff R.Genetic linkage maps ofEucalyptusgrandisandEucalyptusurophyllausing apseudo-testcross:mapping strategy and RAPD markers[J].Genetics,1994,137(4):1121-1137.
[12]Brummer E C,Bouton J H,Kochert G.Development of an RFLP map in diploid alfalfa[J].Theoretical and Applied Genetics,1993,86:329-332.
[13]Tavoletti S,Veronesi F,Osborn T C.RFLP linkage map of an alfalfa meiotic mutant based on an F1population[J].Journal of Heredity,1996,87:167-170.
[14]Echt C S,Kidwell K K,Knapp S J,Osborn T C,McCoy T J.Linkage mapping in diploid alfalfa(Medicagosativa)[J].Genome,1994,37:61-71.
[15]Barcaccia G,Albertini E,Tavoletti S,Falcinelli M,Veronesi F.AFLP fingerprinting inMedicagospp:Its development and application in linkage mapping[J].Plant Breeding,1999,118:335-340.
[16]KalóP,Endre G,Zimányi,Csanádi G,Kiss G B.Construction of an improved linkage map of diploid alfalfa(Medicago sativa)[J].Theoretical and Applied Genetics,2000,100:641-657.
[17]Julier B,Flajoulot S,Barre P,Cardinet G,Santoni S,Huguet T,Huyghe C.Construction of two genetic linkage maps in cultivated tetraploid alfalfa(Medicagosativa)using microsatellite and AFLP markers[J].BMC Plant Biology,2003,3:9.
[18]Choi H K,Kim D,Uhm T,Limpens E,Lim H,Mun J H,Kalo P,Penmetsa R V,Seres A,Kulikova O,Roe B A,Bisseling T,Kiss G B,Cook D R.A sequence-based genetic map ofMedicagotruncatulaand comparison of marker colinearity withM.sativa[J].Genetics,2004,166:1463-1502.
[19]Isobe S,Klimenko I,Ivashuta S,Gau M,Kozlov N N.First RFLP linkage map of red clover(TrifoliumpratenseL.)based on cDNA probes and its transferability to other red clover germplasm[J].Theoretical and Applied Genetics,2003,108:105-112.
[20]Isobe S,Kölliker R,Hisano H,Sasamoto S,Wada T,Klimenko I,Okumura K,Tabata S.Construction of a consensuslinkage map for red clover(TrifoliumpratenseL.)[J].BMC Plant Biology,2009,9:57.
[21]Jones E S,Hughes L J,Drayton M C,Abberton M T,Michaelson-Yeates,T P T,Bowen C,Forster J W.An SSR and AFLP molecular marker-based genetic map of white clover(TrifoliumrepensL.)[J].Plant Science,2003,165:531-539.
[22]Fjellstrom R G,Steiner J J,Beuselink P R.Tetrasomic linkage mapping of RFLP,PCR,and isozyme loci inLotuscorniculatusL[J].Crop Science,2003,43:1006-1020.
[23]Sandal N,Krusell L,Radutoiu S,Olbryt M,Pedrosa A,Stracke S,Sato S,Kato T,Tebata S,Parniske M,Bachmair A,Ketelsen T,Stougaard J.A genetic linkage map of the model legumeLotusjaponicusand strategies for fast mapping of new loci[J].Genetics,2002,161(4),1673-1683.
[24]Xu W W,Sleper D A,Chao S.Genome mapping of polyploidy tall fescue(FestucaarundinaceaSchreb.)with RFLP markers[J].Theoretical and Applied Genetics,1995,91:947-955.
[25]Saha M C,Mian R,Zwonitzer J C,Chekhovskiy K,Hopkins A A.An SSR-and AFLP-based genetic linkage map of tall fescue(FestucaarundinaceaSchreb.)[J].Theoretical and Applied Genetics,2005,110:323-336.
[26]Alm V,Busso C S,Ergon A,Rudi H,Larsen A,Humphreys M W,Rognli O A.QTL analyses and comparative genetic mapping of frost tolerance,winter survival and drought tolerance in meadow fescue(FestucapratensisHuds.)[J].Theoretical and Applied Genetics,2011,123:369-382.
[27]Studer B,Boller B,Bauer E,Posselt U K,Widmer F,Kolliker R.Consistent detection of QTLs for crown rust resistance in Italian ryegrass(LoliummultiflorumLam.)across environments and phenotyping methods[J].Theoretical and Applied Genetics,2007,115:9-17.
[28]Hayward M D,Forster J W,Jones J G,Dolstra O,Evans C,McAdam N J,Hossain K G,Stammers M,Will J,Humphreys M O,Evans G M.Genetic analysis ofLolium.I Identification of linkage groups and the establishment of a genetic map[J].Plant Breeding,1998,117:451-455.
[29]Bert P F,Charmet G,Sourdille P,Hayward M D,Balfourier F.A high-density molecular map for ryegrass(Loliumperenne)using AFLP markers[J].Theoretical and Applied Genetics,1999,99:445-452.
[30]Jones E S,Mahoney N L,Hayward M D,Armstead I P,Jones J G,Humphreys M O,King I P,Kishida T,Yamada T,Balfourier F,Charmet G,Forster J W.An enhanced molecular marker based genetic map of perennial ryegrass(Lolium perenne)reveals comparative relationships with other Poaceae genomes[J].Genome,2002,45:282-295.
[31]Tomaszewski C,Byrne S L,Foito A,Kildea S,Kopecky D,Dolezel J,Heslop-Harrison J S,Stewart D,Barth S.Genetic linkage mapping in an F2perennial ryegrass population using DArT markers[J].Plant Breeding,2012,131(2):345-349.
[32]Inoue M,Gao Z,Hirata M,Fujimori M,Cai H W.Construction of a high-density linkage map of Italian ryegrass(Lolium multiflorumLam.)using restriction fragment length polymorphism,amplified fragment length polymorphism,and telomeric repeat associated sequence markers[J].Genome,2004,47:57-65.
[33]Hirata M,Cai H W,Inoue M,Yuyama N,Miura Y,Komatsu T,Takamizo T and Fujimori M.Development of simple sequence repeat(SSR)markers and construction of an SSR-based linkage map in Italian ryegrass(LoliummultiflorumLam.)[J].Theoretical and Applied Genetics,2006,113:270-279.
[34]Warnke S E,Barker R E,Jung G,Sim S C,RoufMian M A,Saha M C,Brilman L A,Dupal M P,Forster J W.Genetic linkage mapping of an annual×perennial ryegrass population[J].Theoretical and Applied Genetics,2004,109:294-304.
[35]Song Y,Liu F,Zhu Z,Tan L,Fu Y,Sun C,Cai H.Construction of a simple sequence repeat marker-based genetic linkage map in the autotetraploid forage grassDactylisglomerataL[J].Grassland Science,2011,57:158-167.
[36]Xie W,Zhang X,Cai H,Huang L,Peng Y,Ma X.Genetic maps of SSR and SRAP markers in diploid orchardgrass(DactylisglomerataL.)using the pseudo-testcross strategy[J].Genome,2011,54:212-221
[37]Xie W G,Robins J G,Bushman B S.A genetic linkage map of tetraploid orchardgrass(DactylisglomerataL.)and QTL for heading date[J].Genome,2012,55:360-369
[38]Wu X L,Larson S R,Hu Z,Palazzo A J,Jones T A,Wang R R C,Jensen K B,Chatterton N J.Molecular genetic linkage maps for alloteraploidLeymuswildryes(Gramineae:Triticeae)[J].Genome,2003,46:627-646.
[39]Mott I W,Larson S R,Jones T A,Robins J G,Jensen K B,Peel M D.A molecular genetic linkage map identifying the St and H subgenomes ofElymus(Poaceae:Triticeae)wheatgrass[J].Genome,2011,54:819-828.
[40]Lu X P,Yu J P,Gao C P,Surya A.Quantitative trait loci analysis of economically important traits inSorghumbicolor×Sorghumsudaneneshybrid[J].Canadian Journal of Plant Science,2011,91(1):81-90.
[41]Yaneshita M S,Kaneko S,Sasakuma T.Allotetraploidy ofZoysiaspecies with 2n=40based on a RFLP genetic map[J].Theoretical and Applied Genetics,1999,98:751-756.
[42]Cai H W,Inoue M,Yuyama N,Takahasha W,Hirata H,Sasaki T.Isolation,characterization and mapping of simple sequence repeat markers in Zoysiagrass(Zoysiaspp.)[J].Theoretical and Applied Genetics,2005,112:158-166.
[43]Cai W H,Inoue M,Yuyama N,Nakayama S.An AFLP-based linkage map of Zoysiagrass(Zoysiajaponica)[J].Plant Breeding,2004,123:543-548.
[44]Li M L,Yuyama N,Hirata M,Han J G,Wang Y W,Cai H W.Construction of a high-density SSR marker-based linkage map of zoysiagrass(ZoysiajaponicaSteud.)[J].Euphytica,2009,170:327-338.
[45]Harris-Shultz K R,Schwartz B M,Hanna W W.Development,linkage mapping,and use of microsatellites in bermudagrass[J].Journal of the American Society for Horticultural Science,2010,135(6):511-520.
[46]Bethel C M,Sciara E B,Estill J C,Bowers J E,Hanna W,Paterso A H.A framework linkage map of bermudagrass(Cynodondactylon×transvaalensis)based on single-dose restriction fragments[J].Theoretical and Applied Genetics,2006,112:727-737.
[47]Rotter D,Amundsen K,Bonos S A,Meyer W A,Warnke S E,Belanger F C.Molecular genetic linkage map for allotetraploid colonial bentgrass[J].Crop Science,2009,49:1609-1618.
[48]Porceddu A,Albertini E,Barcaccia G,Falistocco E,Falcinelli M.Linkage mapping in apomictic and sexual Kentucky bluegrass(PoapratensisL.)genotypes using a two way pseudo-testcross strategy based on AFLP and SAMPL markers[J].Theoretical and Applied Genetics,2002,104:273-280.
[49]Ortiz J A ,Pessino S C,Bhat V,Hayward M D,Quarín C L.A genetic linkage map of diploidPaspalumnotatum[J].Crop Science,2001,41:823-830.
[50]Studer B,Boller B,Herrmann D,Bauer E,Posselt U K,Widmer F,Kölliker R.Genetic mapping reveals a single major QTL for bacterial wilt resistance in Italian ryegrass(LoliummultiflorumLam.)[J].Theoretical and Applied Genetics,2006,113:661-671.
[51]Inoue M,Gao Z S,Cai H W.QTL analysis of lodging resistance and related traits in Italian ryegrass(LoliummultiflorumLam.)[J].Theoretical and Applied Genetics,2004,109:1576-1585.
[52]Muylle H,Baert J,Bockstaele E V,Pertijs J,Roldán-Ruiz I.Four QTLs determine crown rust(Pucciniacoronataf.sp.lolii)resistance in a perennial ryegrass(Loliumperenne)population[J].Heredity,2005,95,348-357.
[53]Schejbel B,Jensen L B,Asp T,Xing Y,Lübberstedt T.QTL analysis of crown rust resistance in perennial ryegrass under conditions of natural and artificial infection[J].Plant breeding,2007,126,347-352
[54]Pfender W F,Saha M C,Johnson E A,Slabaugh M B.Mapping with RAD (restriction-site associated DNA)markers to rapidly identify QTL for stem rust resistance inLoliumperenne[J].Theoretical and Applied Genetics,2011,122:1467-1480.
[55]Schejbel B,Jensen L B,Asp T,Xing Y,Lübberstedt T.Mapping of QTL for resistance to powdery mildew and resistance gene analogues in perennial ryegrass[J].Plant breeding,2008,127,368-375
[56]Curley J,Sim S C,Warnke S,Leong S,Barker R,Jung G.QTL mapping of resistance to gray leaf spot in ryegrass[J].Theoretical and Applied Genetics,2005,111:1107-1117.
[57]Dracatos P M,Cogan N O I,Sawbridge T I,Gendall A R,Smith K F,Spangenberg G C,Forster J W.Molecular characterisation and genetic mapping of candidate genes for qualitative disease resistance in perennial ryegrass(LoliumperenneL.)[J].BMC Plant Biology,2009,9:62.
[58]Studer B,Jensen L B,Hentrup S,Brazauskas G,Kölliker R,Lübberstedt T.Genetic characterisation of seed yield and fertility traits in perennial ryegrass(LoliumperenneL.)[J].Theoretical and Applied Genetics,2008,117:781-791.
[59]Armstead I P,Turner L B,Marshall A H,Humphreys M O,King I P,Thorogood D.Identifying genetic components controlling fertility in the outcrossing grass species perennial ryegrass(Loliumperenne)by quantitative trait loci analysis and comparative genetics[J].New Phytologist,2008,178:559-571.
[60]Armstead I P,Skøt L,Turner L B,Skøt K,Donnison I S,Humphreys M O,King I P.Identification of perennial ryegrass(Loliumperenne(L.))and meadow fescue(Festucapratensis(Huds.))candidate orthologous sequences to the rice Hd1(Se1)and barley HvCO1CONSTANS-like genes through comparative mapping and microsynteny[J].New Phytologist,2005,167:239-247.
[61]Byrne S,Guiney E,Barth S,Donnison I,Mur L A J,Milbourne D.Identification of coincident QTL for days to heading,spike length and spikelets per spike inLoliumperenneL[J].Euphytica,2009,166:61-70.
[62]Armstead I P,Turner L B,Farrell M,Skøt L,Gomes P,Montoya T,Donnison I S,King I P,Humphreys M O.Synteny between a major heading-date QTL in perennial ryegrass(LoliumperenneL.)and the Hd3heading-date locus in rice[J].Theoretical and Applied Genetics,2004,108:822-828.
[63]Jensen L B,Andersen J R,Frei U,Xing Y Z,Taylor C,Holm P B,Lübberstedt T.QTL mapping of vernalization response in perennial ryegrass(LoliumperenneL.)reveals co-location with an orthologue of wheat VRN1[J].Theoretical and Applied Genetics,2005,110:527-536.
[64]Cogan N O I,Smith K F,Yamada T,Francki M G,Vecchies A C,Jones E S,Spangenberg G C,Forster J W.QTL analysis and comparative genomics of herbage quality traits in perennial ryegrass(LoliumperenneL.)[J].Theoretical and Applied Genetics,2005,110:363-380.
[65]Turner L B,Farrell M,Humphreys M O,Dolstra O.Testing water-soluble carbohydrate QTL effects in perennial ryegrass(LoliumperenneL.)by marker selection[J].Theoretical and Applied Genetics,2010,121:1405-1417.
[66]Barre P,Moreau L,Mi F,Turner L,Gastal F,Julier B,Ghesquière M.Quantitative trait loci for leaf length in perennial ryegrass(LoliumperenneL.)[J].Grass and Forage Science,2009,64,310-321.
[67]Faville M J,Jahufer M Z Z,Hume D E,Cooper B M,Pennell C G L,Ryan D L,Easton H S.Quantitative trait locus mapping of genomic regions controlling herbage yield in perennial ryegrass[J].New Zealand Journal of Agricultural Reaearch,2012,55(3):263-281.
[68]Pearson A,Cogan N O I,Baillie R C,Hand M L,Bandaranayake C K,Erb S,Wang J P,Kearney G A,Gendall A R,Smith K F,Forster J W.Identification of QTLs for morphological traits influencing waterlogging tolerance in perennial ryegrass(LoliumperenneL.)[J].Theoretical and Applied Genetics,2011,122:609-622.
[69]Hegarty M,Yadav R,Lee M,Armstead I,Sanderson R,Scollan N,Powell W,Skøt L.Genotyping by RAD sequencing enables mapping of fatty acid composition traits in perennial ryegrass(Loliumperenne(L.))[J].Plant Biotechnology Journal,2013.11(5)572-581.
[70]Brown R N,Barker R E,Warnke S E,Cooper L D,Brilman L A,Mian M A R,Jung G,Sim S C.Identification of quantitative trait loci for seed traits and floral morphology in a field-grownLoliumperenne×Loliummultiflorummapping population[J].Plant Breeding,2010,129,29-34.
[71]Alm V,Busso C S,Ergon A,Rudi H,Larsen A,Humphreys M W,Rognli O A.QTL analyses and comparative genetic mapping of frost tolerance,winter survival and drought tolerance in meadow fescue(FestucapratensisHuds.)[J].Theoretical and Applied Genetics,2011,123:369-382.
[72]Ergon A,Fang C,JørgensenØ,Aamlid T S,Rognli O A.Quantitative trait loci controlling vernalization requirement,heading time and number of panicles in meadow fescue(FestucapratensisHuds.)[J].Theoretical and Applied Genetics,2006,112:232-242.
[73]Larson S R,Mayland H F.Comparative mapping of fiber,protein,and mineral content QTLs in two interspecific Leymus wildrye full-sib families[J].Molecular Breeding,2007,20:331-347.
[74]Larson S R,Kellogg E A.Genetic dissection of seed production traits and identification of a major-effect seed retention QTL in hybridLeymus(Triticeae)wildryes[J].Crop Science,2009,49:29-40.
[75]Hu Z M,Wu X L,Larson S R,Wang R R C,Jones T A,Chatterton N J,Palazzo A J.Detection of linkage disequilibrium QTLs controlling low-temperature growth and metabolite accumulations in an admixed breeding population ofLeymuswildryes[J].Euphytica,2005,141:263-280.
[76]Kaur P,Larson S R,Bushman B S,Wang R R C,Mott I W,Hole D,Thimmapuram J,Gong G,Liu L.Genes controlling plant growth habit inLeymus(Triticeae):Maize barren stalk1(ba1),rice lax panicle,and wheat tiller inhibition(tin3)genes as possible candidates[J].Functional &Integrative Genomics,2008,8:375-386.
[77]Larson S R,Wu X L,Jones T A,Jensen K B,Chatterton N J,Waldron B L,Robins J G,Bushman B S,Palazzo A J.Comparative mapping of growth habit,plant height,and flowering QTLs in two interspecific families of Leymus[J].Crop Science,2006,46:2526-2539.
[78]Robins J G,Luth D,Campbell T A,Bauchan G R,He C,Viands D R,Hansen J L,Brummer E C.Genetic mapping of biomass production in tetraploid alfalfa[J].Crop Science,2007,47:1-10.
[79]Brouwer D J,Duke S H,Osborn T C.Mapping genetic factors associated with winter hardiness,fall growth,and freezing injury in autotetraploid alfalfa[J].Crop Science,2000,40:1387-1396.
[80]Robins J G,Brummer E C.QTL underlying self-fertility in tetraploid alfalfa[J].Crop Science,2010,50:143-149.
[81]Sledge M K,Bouton J H,Dall’Agnoll M,Parrott W A,Kochert G.Identification and confirmation of Aluminum tolerance QTL in diploidMedicagosativasubsp.coerulea[J].Crop Science,2002,42:1121-1128.
[82]Narasimhamoorthy B,Bouton J H,Olsen K M,Sledge M K.Quantitative trait loci and candidate gene mapping of aluminum tolerance in diploid alfalfa[J].Theoretical and Applied Genetics,2007,114:901-913.
[83]Espinoza L C L,Julier B.QTL detection for forage quality and stem histology in four connected mapping populations of the model legumeMedicagotruncatula[J].Theoretical and Applied Genetics,2013,126(2):497-509.
[84]Espinoza L C L,Huguet T,Julier B.Multi-population QTL detection for aerial morphologenetic traits in the model legumeMedicagotruncatula[J].Theoretical and Applied Genetics,2012,124(4):739-754.
[85]Barrett B A,Baird I J,Woodfield D R.A QTL analysis of white clover seed production[J].Crop Science,2005,45:1844-1850.
[86]Herrmann D,Boller B,Studer B,Widmer F,Kölliker R.QTL analysis of seed yield components in red clover(Trifolium pratenseL.)[J].Theoretical and Applied Genetics,2006,112:536-545.
[87]Herrmann D,Boller B,Studer B,Widmer F,Kölliker R.Improving persistence in red clover insights from QTL analysis and comparative phenotypic evaluation[J].Crop Science,2008,48:269-277.
[88]Gondo T,Sato S,Okumura K,Tabata S,Akashi R,Isobe S.Quantitative trait locus analysis of multiple agronomic traits in the model legumeLotusjaponicas[J].Genome,2007,50:627-637.
[89]Jessup R W,Renganayaki K,Reinert J A,Genovesi A D,Engelke M C,Paterson A H,Kamps T L,Schulze S,Howard A N,Giliberto B,Burson B L.Genetic mapping of fall armyworm resistance in Zoysiagrass[J].Crop Science,2011,51:1774-1783.
[90]丁成龙,刘颖,许能祥,顾洪如.日本结缕草抗寒相关性状的 QTL分析[J].草地学报,2010,18(5):703-707.
[91]Chakraborty N,Curley J,Warnke S,Casler M D,Jung G.Mapping QTL for dollar spot resistance in creeping bentgrass(AgrostisstoloniferaL.)[J].Theoretical and Applied Genetics,2006,113:1421-1435.
[92]Xie W G,Robins J,Bushman B S.A search for candidate genes affecting late heading in orchardgrass/cocksfoot(DactylisglomerataL.)[A].Bushman S,Spangenberg G C.Proceedings of the 7th International Symposium on the Molecular Breeding of Forage and Turf[C].Salt Lake City:USDA-FRRL,2013:14-16.
[93]Tamura K,Yamada T.A perennial ryegrass CBF gene cluster is located in a region predicted by conserved synteny between Poaceae species[J].Theoretical and Applied Genetics,2007,114 :273-283.
[94]沈立爽,朱立煌.植物的比较基因组研究和大遗传系统[J].生物工程进展,1995,15:23-28.
[95]Bonierable M W,Plaisted P L,Tanksley S D.RFLP maps based on a common set of clones reveal modes of chromosomal evolution in potato and tomato[J].Genetics,1998,120:1095-1103.
[96]Tanksley S D,Bernatzky R,Lapitan N L,Prince J P.Conservation of gene repertoire but not gene order in pepper and tomato[J].Proceedings of the National Academy of Sciences of the United States of America,1988,85(17):6419.
[97]Devos K M,Gale M D.Genome relationship:the grass model in current research[J].The Plant Cell,2000,12:637-646.
[98]Grivet L,D’Hont A,Dufour P,Hamon P,Roques D,Glaszmann J C.Comparative genome mapping of sugar cane with other species within the Andropogoneae tribe[J].Heredity,1994,73(5):500-508.
[99]Andersen J R,Jensen L B,Asp T,Lübberstedt T.Vernalization response in perennial ryegrass(LoliumperenneL.)involves orthologues of diploid wheat(Triticummonococcum)VRN1and rice(Oryzasativa)Hd1[J].Plant Molecular Biology,2006,60:481-494
[100]徐云碧,朱立煌.分子数量遗传学[M].北京:中国农业出版社,1994:81-83.
