2,4-表油菜素内酯和赤霉素互作对羊草种子萌发及幼苗生长的影响
2014-12-24郭慧琴任卫波武自念万东莉
郭慧琴,任卫波,李 平,武自念,万东莉
(1.内蒙古农业大学生命科学学院,内蒙古 呼和浩特010018;2.中国农业科学院草原研究所,内蒙古 呼和浩特010010)
羊草(Leymuschinesis),又名碱草,是禾本科赖草属多年生根茎型禾草,广泛分布于我国东北西部和内蒙古中东部地区,是草甸草原和典型草原的优势种之一。羊草不仅具有营养价值高、适口性好、产草量高等优点,还具有耐寒、耐盐碱等特性,具有重要的生态价值和经济价值[1]。然而,羊草有性繁殖能力低,种子休眠严重,发芽率低,当年采集的种子发芽率仅为6%~10%[2-3],这一“三低”问题严重影响了羊草栽培草地的建设和退化草原的补播恢复。因此,如何打破羊草种子休眠,提高羊草种子发芽率一度成为羊草研究的热点之一。
围绕羊草种子休眠机制及打破休眠的途径和方法,国内外已开展了大量的研究。目前认为,导致羊草种子休眠、发芽率低的主要原因是羊草种子的稃片和胚乳中存在ABA等抑制物[4]。打破羊草种子休眠的方法主要包括去除稃片、变温处理、聚乙二醇处理、外源激素调控等方面[2]。去除内外稃后,羊草种子发芽率可由6%提高到45%;清水浸种1d,羊草种子发芽率可提高到36%,NaOH和水复合处理后,发芽率可达84%[2]。除此之外,外源激素调控是打破羊草种子休眠,提高发芽率的有效途径,如,赤霉素、GA3、IAA、氟啶酮(FL)等激素对羊草种子发芽和种苗生长有促进作用[5-8]。
油菜素内酯(BRs)是继生长素、赤霉素、细胞分裂素、脱落酸、乙烯以后的第六大类植物激素,广泛分布于植物的根、茎、叶、花器和种子中,参与植物种子萌发、株型生长与发育、衰老、干旱、寒冷等逆境响应等重要过程[9]。已有的研究表明,无论是在逆境胁迫下还是在正常环境下,油菜素内酯都能够显著促进植物种子萌发[10-11]。因此,油菜素内酯在打破羊草种子休眠,提高种子发芽能力等方面具有一定的潜力。然而,目前有关油菜素内酯对羊草种子发芽率及幼苗生长影响的研究较少。那么油菜素内酯到底对羊草种子萌发及种苗生长有什么影响?不同浓度间是否存在显著差异?油菜素内酯与赤霉素之间是否存在互作效应?回答这些问题,将有望为解决羊草的“三低”问题提供新的方法和途径。
1 材料与方法
1.1 试验材料
供试野生羊草种子于2012年9月采自内蒙古锡林浩特市毛登牧场。采集后的种子在室温下晾干清选后,装入布袋,放在4℃下保存备用。
1.2 试验设计
激素处理试验采用双因素设计,其中双因素为赤霉素GA3和2,4-表油菜素内酯(EBR)两种激素,每种激素设4个水平,其中GA3为0、200、300、500 μg·g-14个水平;2,4-表油菜素内酯为0、0.01、0.10、1.00μg·g-14个水平,两激素组合共16个处理(表1),每处理设4次重复,每个重复选用50粒羊草种子。
1.3 种子激素处理
根据试验设计浓度(表1)配置激素溶液。精选饱满的羊草种子,浸泡在相应的激素溶液中,室温下处理48h后,将种子捞出,并用蒸馏水冲洗3~5次,用0.1%NaClO消毒10min,再用蒸馏水冲洗2~3次,将种子置于培养皿内发芽。(P<0.05)。与对照相比,GA3处理后羊草种子发芽率增加3~4倍,处理与对照间差异显著(P<0.05)(图1)。随着GA3浓度的升高,羊草种子萌发率呈先上升后下降的趋势,其中以300μg·g-1GA3处理种子萌发率最高,可达44%(图1)。
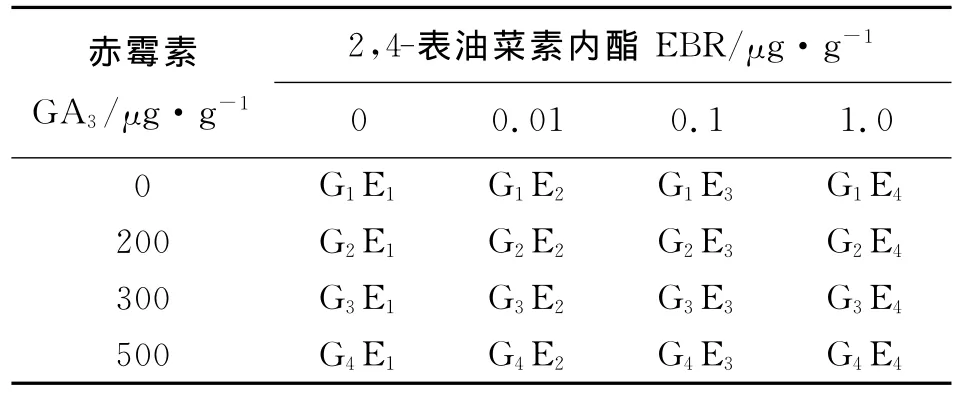
表1 赤霉素与2,4-表油菜素内酯浓度组合Table 1 Combination of different concentrations of gibberellin and brassnoids
油菜素内酯与赤霉素在促进羊草种子萌发方面有显著的互作效应(P<0.00)(表2)。不同浓度组合间存在显著差异,其中以2,4-表油菜素内酯0.10 μg·g-1+GA3200μg·g-1组合最佳,处理后羊草种子萌发率可达76%(图1),比单一2,4-表油菜素
1.4 种子发芽
将浸种过的种子,放入预先备好的培养皿中(蒸馏水充分润湿、装有双层滤纸的直径为12cm的培养皿)进行发芽试验。培养条件为:16℃(16h)/28℃(8h),光照时间8h。每天称重补水,并统计种子萌发数,萌发周期为16d[3]。种子突破种皮即视为萌发。每处理随机选取20株羊草幼苗,用测量纸测量其苗长和根长。
萌发率=发芽结束后发芽种子数/供试种子总粒数×100%。
1.5 数据处理
试验数据用SPSS 14.0对所获取数据进行统计分析,用平均值和标准误表示测定结果。对2,4-表油菜素内酯、GA3及其互作对羊草种子萌发及种苗生长的影响进行二因素方差分析,并用Duncan法进行多重比较;采用Excel 2003制图。
2 结果与分析
2.1 2,4-表油菜素内酯与赤霉素处理对羊草种子(EBR)发芽的影响
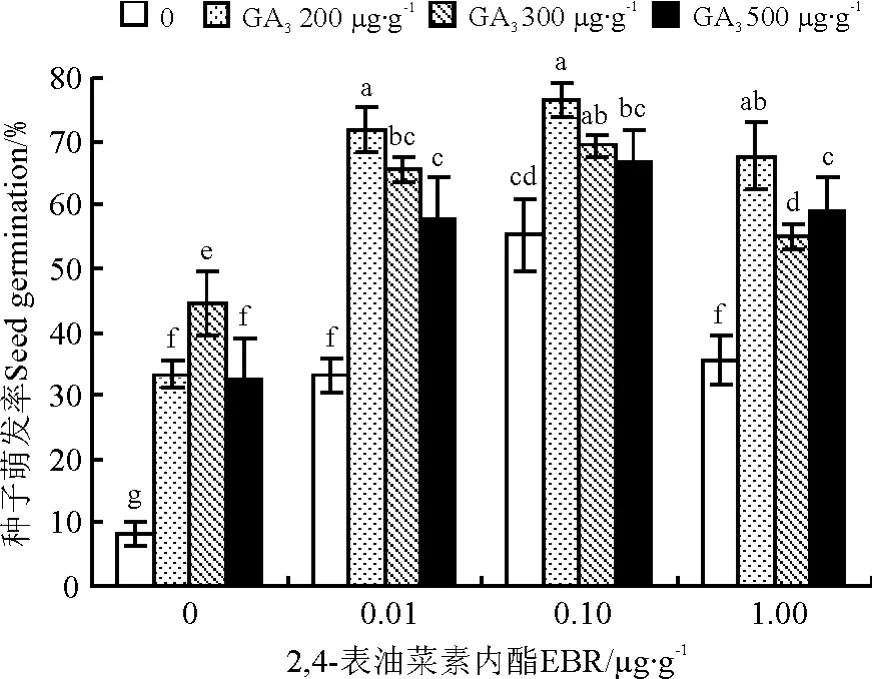
图1 2,4-表油菜素内酯与赤霉素复合处理对羊草种子发芽率的影响Fig.1 Interaction effect of EBR and GA3on the germination of Leymus chinesis
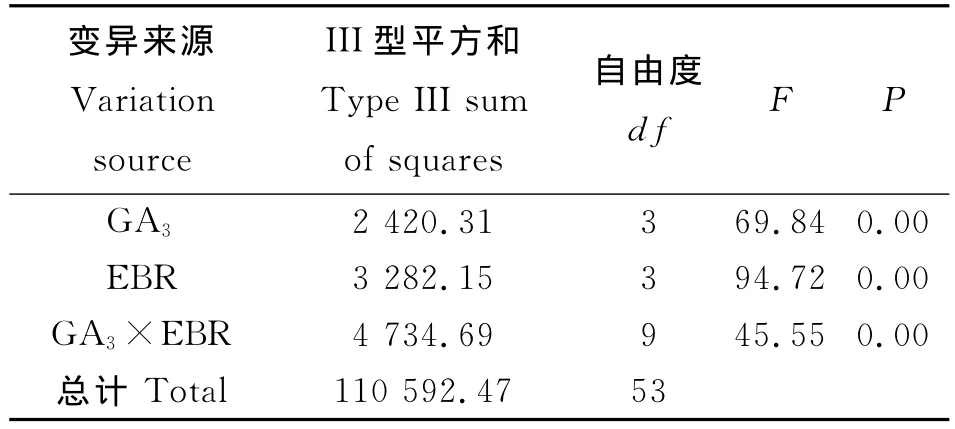
表2 不同激素与浓度互作对羊草种子萌发影响的方差分析Table 2 Two way ANVOA for the interaction effect of hormones and different concentration on germination
2,4-表油菜素内酯对羊草种子萌发率有显著影响(P<0.05)。与对照相比,2,4-表油菜素内酯处理后,羊草种子萌发率增加了2~4倍,差异显著(P<0.05)(图1)。随着浓度的升高,羊草种子萌发率呈先上升后下降的趋势,其中以0.1μg·g-12,4-表油菜素内酯处理后种子萌发率最高,可达54%(图1)。GA3对羊草种子萌发率也有显著影响内酯处理增加21%,比单一GA3处理增加32%。其次为2,4-表油菜素内酯0.01μg·g-1+ GA3200μg·g-1,比上述最佳组合略低6%,两者间差异不显著(P>0.05)(图1)。
2.2 2,4-表油菜素内酯与赤霉素处理对羊草幼苗生长的影响
2,4-表油菜素内酯能同时促进羊草幼苗根和苗的生长。与对照相比,2,4-表油菜素内酯处理后羊草幼苗苗长增加了11%~24%,根长增加了35%~40%,且对根长的影响差异均达到显著水平(P<0.05)(图2、3)。随着2,4-表油菜素内酯浓度的升高,羊草幼苗的根长、苗长均呈现先上升后下降的趋势,但未达到显著水平。
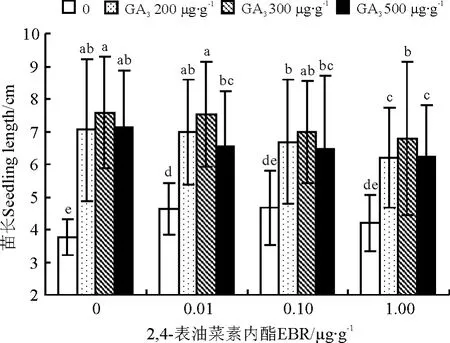
图2 2,4-表油菜素内酯与赤霉素复合处理对羊草种苗苗长的影响Fig.2 Interaction effect of EBR and GA3on the seedling growth of L .chinesis
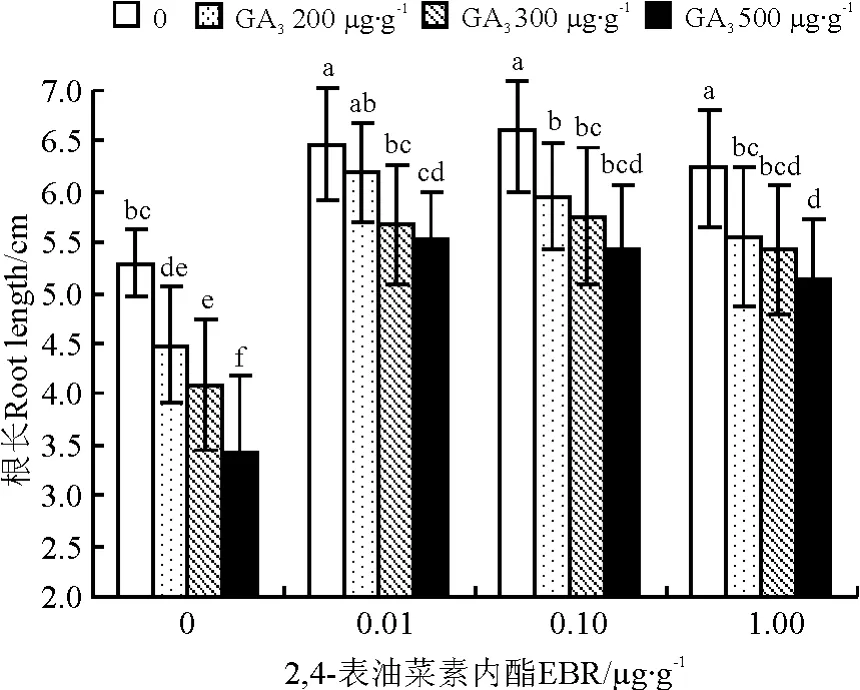
图3 2,4-表油菜素内酯与赤霉素复合处理对羊草种苗根长的影响Fig.3 Interaction effect of EBR and GA3on the root length of L .chinesis
GA3能够促进羊草幼苗生长,但对根生长有一定抑制作用。与对照相比,GA3处理后苗长增加了87%~101%,差异显著(P<0.05)(图2)。随着GA3浓度的升高,苗长呈先上升后下降的趋势,其中以300μg·g-1GA3处理的苗长最长;而根长呈显著下降趋势,降低了15%~35%,且以500 μg·g-1GA3处理根长最短(P<0.05)(图3)。
油菜素内酯与赤霉素组合在促进羊草种苗生长方面有显著的互作效应(P<0.05)(表3),不同浓度组合间存在差异。对于苗长,以2,4-表油菜素内酯0.01μg·g-1+ GA3300μg·g-1组合最佳(图2);其次为2,4-表油菜素内酯0.01μg·g-1+GA3200 μg·g-1和2,4-表油菜素内酯0.1μg·g-1+GA3300μg·g-1组合,均比最佳组合处理低7.61%(P>0.05);再 次 为 2,4-表 油 菜 素 内 酯 0.1 μg·g-1+GA3200μg·g-1组合,比最佳组合降低了11.42%(P<0.05)。
对于根长,以2,4-表油菜素内酯0.01μg·g-1+GA3200μg·g-1组合最佳(图3),该组合处理后根长仅比单一2,4-表油菜素内酯最佳浓度处理略低6.49%(P<0.05);其次为2,4-表油菜素内酯0.10 μg·g-1+GA3200μg·g-1组合(图3),比最佳组合低3.87%,两者间差异也未达到显著水平(P>0.05)。
3 讨论
3.1 油菜素内酯对植物种子萌发及幼苗生长的影响
作为六大植物激素之一,油菜素内酯对植物种子萌发及幼苗生长有重要的调控作用。适宜浓度的外源油菜素内酯如2,4-表油菜素内酯处理可促进植物种子萌发和幼苗生长,尤其是干旱和盐胁迫等逆境条件下,效果更加明显。如,0.05μg·g-12,4-表油菜素内酯处理后,茄子(Solanummelongema)在150mol·L-1NaCl盐胁迫下的种子发芽率提高8%,幼苗株高和根长分别增加了56.67%和23.83%[11]。经2,4-表油菜素内酯和硝普钠复合处理后,玉米(Zeamays)种子的发芽率可提高15%~16%,主根长和芽长也有增加[12]。油菜素内酯对黄豆(Glycinemax)种子萌发及下胚轴生长也有显著的促进作用,0.05μg·g-1油菜素内酯处理后,黄豆种子发芽率可提高14%~19%,下胚轴长度增加79%[13]。李凯荣和韩刚[14]也发现,油菜素内酯对柠条(Caraganakorshinskii)和紫穗槐(Amorpha fruticosa)种子萌发及下胚轴生长有显著影响,且随着浓度的增加,种子发芽率呈先升高后降低的趋势,其中以0.05μg·g-1油菜素内酯处理效果最佳,与对照相比,种子发芽率可提高20%~23%。另外,外源BR可有效缓解拟南芥(Arabidopsisthaliana)GA3和ABA突变体的低发芽率问题[15]。在拟南芥中,过量表达dwf4基因(BR合成关键基因之一),能够有效解除因ABA引起的种子休眠、恢复和提高拟南芥种子的发芽率[16]。过量表达HSD1基因(该基因参与BR的生物合成)后可有效降低拟南芥种子休眠[17]。本研究也发现,2,4-表油菜素内酯对羊草种子萌发有显著影响,适宜浓度下羊草种子萌发率显著提高,这与其他植物已有的研究结果是一致的。BR对种子发芽的促进作用可能是通过乙醇胺磷酸甘油酯(Phosphatidylethanolamine)结合蛋白(MFT)调控来实现的[18]。
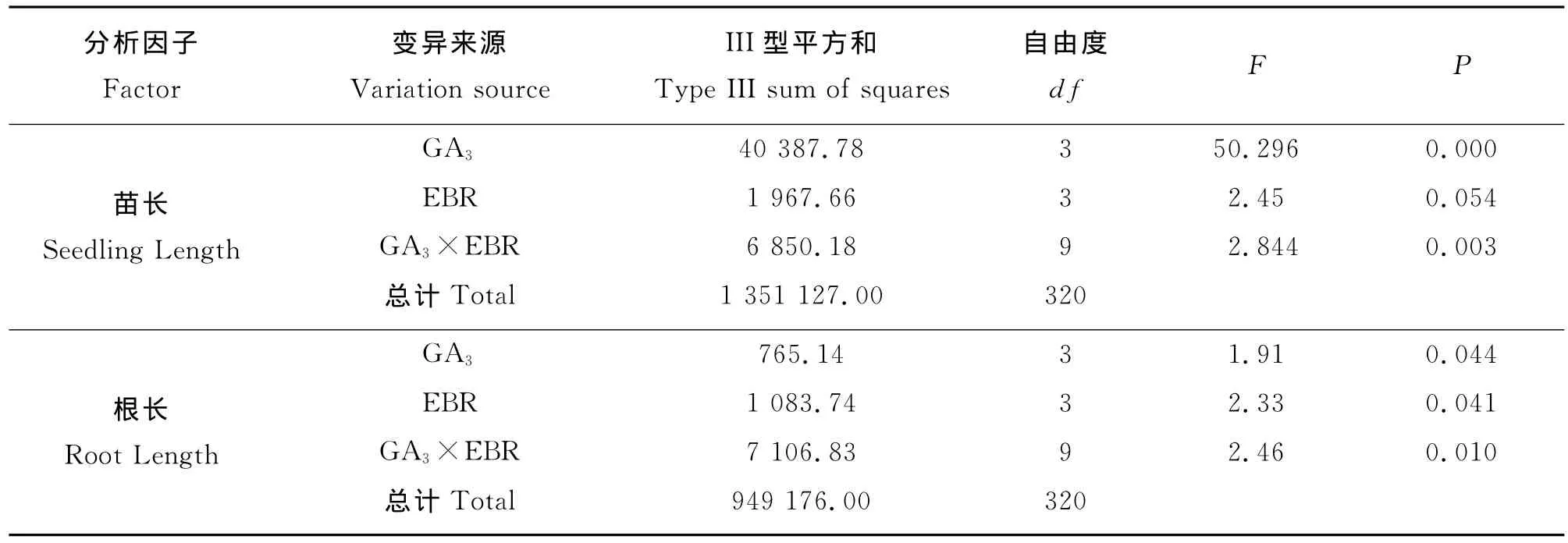
表3 不同激素与浓度互作对羊草种苗生长影响的方差分析Table 3 Two way ANVOA for the interaction effect of hormones and different concentrations on seedling growths
此外,本研究发现,适宜浓度的2,4-表油菜素内酯能够显著促进羊草幼苗根的伸长,这与已有的研究结果一致。如,低浓度BR能够同时提高拟南芥野生型和BR缺失突变体幼苗的根长[19],在玉米野生型上也有类似的效果[20],而且BR引发的根伸长作用途径可能是与生长素调控途径不同的独立途径,因为这种促进效应不受生长素转运抑制因子的影响[20]。
3.2 赤霉素对羊草种子萌发及幼苗生长的影响
本研究发现,赤霉素对羊草种子萌发率及种苗生长有显著影响,而且随着赤霉素浓度的增加,种子发芽率、苗长呈先升高后降低的趋势,但根长受到不同程度的抑制。这一结果与多数已有的研究结果是一致的。不同赤霉素浓度下羊草种子萌发率呈先增加后降低的单峰曲线变化,当赤霉素浓度为300 μg·g-1时,羊草种子萌发率比对照提高9%,浓度超过400μg·g-1时,羊草种子萌发率显著降低[6]。这说明低浓度的赤霉素可促进羊草种子萌发,但高浓度则对羊草种子萌发有明显的抑制作用。300 μg·g-1赤霉素处理后,羊草的苗长显著高于对照,根长和根冠比则呈逐步减少[6]。100~300μg·g-1GA3处理对羊草种子发芽表现为促进作用,当GA3浓度超过400μg·g-1时,对羊草种子萌发率表现出不同程度的抑制[8]。与此同时,GA3处理显著促进羊草幼苗伸长,但对根有一定的抑制作用,根冠比降低[8]。然而,也有研究发现,不同浓度赤霉素处理对羊草种子发芽率无显著影响[2]。造成差异的原因可能在于:1)处理方法的不同。得到显著性差异结果的研究多是采用浸泡法即在发芽前将羊草种子完全浸泡在激素溶液中;而非显著性的研究则是在种子发芽过程中,将赤霉素溶液添加进培养皿中,羊草种子未能与激素溶液充分接触,这可能是导致处理效果截然不同的原因之一。采用添加法对羊草种子进行赤霉素处理后,也发现赤霉素对羊草种子萌发无显著影响[4]。最近的研究为这一推测提供了进一步的证据。闫小红等[21]研究发现,2,4-表油菜素内酯浸泡处理辣椒(Capsicumannuum)种子后,其种子的发芽率显著提高,但同样浓度的2,4-表油菜素内酯采用添加处理后,辣椒种子发芽率与对照相比无显著变化。2)羊草种子来源不同。不同生态环境导致了羊草种子的品质和发芽率存在显著差异。已有的研究表明,不同来源的羊草种子经过相同的处理后,种子发芽率表现出一定差异。同样在300 μg·g-1赤霉素处理下,吉林长岭地区采集的野生羊草种子发芽率为16.7%,黑龙江安达草原采集的野生羊草发芽率可达34.45%[8]。
3.3 2,4-表油菜素内酯与GA3组合对植物种子萌发及幼苗生长的影响
已有的研究表明,2,4-表油菜素内酯是种子萌发过程中GA3生物学功能的促进因子,两者在促进植物种子萌发和幼苗生长方面具有显著的互作效应。如,2,4-表油菜素内酯与 GA3在促进绿豆(Vignaradiata)上胚轴和黄瓜(Cucumissativus)下胚轴生长有叠加效应[22],在促进拟南芥幼苗下胚轴伸长生长方面具有协同作用,同时使用两种激素的下胚轴长度要比单独使用一种激素的高出3~4倍[23]。本研究结果与已有的研究基本一致。同时发现,使用2,4-表油菜素内酯和GA3处理后的羊草种子发芽率要比单一使用GA3或2,4-表油菜素内酯高出20%~30%,这说明2,4-表油菜素内酯和GA3在促进羊草种子萌发方面具有叠加效应。在促进羊草种苗生长方面,双激素处理后,羊草种苗的苗长和根长均可显著增加,可实现在促进幼苗生长的同时,弥补GA3对根长的抑制作用,两者在促进种苗生长方面表现出了显著的协同效应。
4 结论
1)2,4-表油菜素内酯浓度在0.01~0.1 μg·g-1内时,能够促进羊草种子萌发和种苗生长,尤其是对根的促进效果更加明显。
2)对于羊草种子萌发及种苗生长,2,4-表油菜素内酯和赤霉素存在显著的互作效应,综合考虑羊草种子发芽率、根长和苗长,以2,4-表油菜素内酯0.1μg·g-1+GA3200μg·g-1、2,4-表油菜素内酯0.01μg·g-1+ GA3200μg·g-1效果最好。
[1]杨丽娜,宝音陶格涛.不同改良措施下羊草群落生物量的研究[J].中国草地学报,2010,32(1):86-91.
[2]何学青,胡小文,王彦荣.羊草种子休眠机制及破除方法研究[J].西北植物学报,2010(1):120-125.
[3]马红媛,吕丙盛,杨昊谕,闫超,梁正伟.松嫩平原退化草地羊草种子萌发对环境因子的响应[J].植物生态学报,2012,36(8):812-818.
[4]黄晓辉,胡小文,徐宗海.羊草种子休眠和萌发的激素调控研究[J].草业学报,2013,22(5):183-189.
[5]刘彩红,李成云.GA3浸种对羊草种子发芽和幼苗生长的影响[J].草业科学,2011,28(5):797-801.
[6]刘彩红,李成云,朴光一.2,4-二氯苯氧乙酸对不同羊草种子发芽特性的影响[J].东北农业大学学报,2011,42(12):125-129.
[7]李晓宇,蔺吉祥,穆春生.外源植物激素对羊草种子形成及萌发的影响[J].中国农学通报,2012,28(23):11-14.
[8]马红媛,梁正伟,黄立华,闫超,孔祥军.4种外源激素处理对羊草种子萌发和幼苗生长的影响[J].干旱地区农业研究,2008,26(2):69-73.
[9]Yang C J,Zhang C,Lu Y N.The mechanisms of brassinosteroids’action:From signal transduction to plant development[J].Molecular Plant,2011,4(4):588-600.
[10]Viret C,Eugenia R,Christophe R.Boosting crop yield with plant steroids[J].The Plant Cell,2012,24:842-857.
[11]吴雪霞,查丁石,朱宗文.外源2,4-表油菜素内酯对盐胁迫下茄子种子萌发和幼苗生理特性的影响[J].植物生理学报,2011,47(6):607-612.
[12]张红.硝普钠、2,4-表油菜素内酯\水杨酸浸种对盐胁迫下玉米种子萌发及幼苗生长的影响[J].核农学报,2012,26(1):0164-0169.
[13]程彦伟,柳家婷,韩建民,杨亚会.油菜素内酯对黄豆种子萌发及下胚轴和胚根生长的影响[J].洛阳师范学院学报,2013,32(11):57-59.
[14]李凯荣,韩刚.天然油菜素内酯促进柠条和紫穗槐种子发芽及下胚轴伸长效应[J].西北林学院学报,2003,18(4):17-20.
[15]Steber C M,McCourt P.A role for brassinosteroids in germination inArabidopsis[J].Plant Physiology,2001,125:763-769.
[16]Divi U K,Krishna P.Overexpression of the brassinosteroid biosynthetic geneAtDWF4inArabidopsisseeds overcomes abscisic acid-induced inhibition of germination and increases cold tolerance in transgenic seedlings[J].Journal of Plant Growth Regulation,2010,29:385-393.
[17]Baud S.Regulation ofHSD1in seeds ofArabidopsisthaliana[J].Plant Cell Physiology,2009,50:1463-1478.
[18]Xi W,Yu H.Mother of FT and TEL1regulates seed germination and fertility relevant to the brassinosteroid signaling pathway[J].Plant Signal Behavior,2010,5:1315-1317.
[19]Kim S K.Involvement of brassinosteroids in the gravitropic response of primary root of maize[J].Plant Physiology,2000,123:997-1004.
[20]Mussig C,Shin G H,Altmann T.Brassinosteroids promote root growth inArabidopsis[J].Plant Physiology,2003,133:1261-1271.
[21]闫小红,胡文海,曾受鑫.低温胁迫下2,4-表油菜素内酯对辣椒种子萌发及幼苗生长的影响[J].华中农业大学学报,2012,31(5):563-568.
[22]Gregory L E,Mandava N B.The activity and interaction of brassinolide and gibberellic acid in mung bean epicotyls[J].Plant Physiology,1982,54:239-243.
[23]Tanaka K,Nakamura Y,Asami T.Physiological roles of brassinosteroids in early growth ofArabridopsis:Brassinosteroids have a synergistic relationship with gibberellin as well as auxin in light-grown hypocotyl elongation[J].Plant Growth Regulation,2003,22:259-271.
