基于特示关系的生态学信息及其认识论意蕴*
2014-12-21范念念
王 义 范念念
(1.东北大学文法学院科技与社会研究中心,沈阳110819;2.四川大学水力学与山区河流开发保护国家重点实验室、水利水电学院,成都610065)
美国生态心理学家Gibson 的生态学方法是在实在论的背景中提出来的。实在论和反实在论争论问题的核心是,被感知的环境是否独立于感知者,如果独立,如何来证明它的独立?显然,这种哲学争论实际上已经超出科学实证的界限。因为实证有效的前提是理论只有指向的是客观事实,实证方法才能有效,而这里由于涉及到客观事实和对客观事实的感知,因而付诸于实证来证明实在论是无效的。对于实在论者来说,如果坚持被感知的世界是实在的这个前提,那么就需要说明,实在的世界和被感知的世界如何是同一个世界。然而对于Gibson 来说,实在论的证明是可能的,它就是能量运动构型和实在之间的特示关系。
一、引言:心理学的生态学方法
特示关系蕴含在心理学的生态学方法之中。在论述特示之前,必须首先阐述生态学方法的实际内涵。到目前为止对生态心理学的理解中,国内学者仍然延循着Lombardo 的观点,即,Gibson 生态心理学的总原理是相互作用(Lombardo,1987),它蕴含着一种整体主义视角。然而,整体主义对还原主义、机械论的批判并不能足够精确地表达出生态学方法的实质内涵。因为符合“方法”的必要条件是,它必须首先达到定量性的标准,才能具有可操作性。
首先需要知道,即使Gibson 本人也没有对生态学方法进行单独论述,它只是贯穿在其对心理学的实际应用中。在The ecological approach to visual perception 一书开篇之处,Gibson 提到“值得牢记却经常被忽略的事实是:动物和环境这两个词是不可分割的一对,一个词暗示着另一个词的存在,没有周围环境动物无法生存……动物-物体被以一种特殊方式环绕着,一个环境围绕一个生命体的方式与一堆物体围绕着一个物理物是不同的”(Gibson,1986,p.8),这可以看作Gibson 对生态学方法最为素朴的表达。
生态学方法分为三个有机研究的维度。一是环境与动物之间的关联或互补,Gibson 将之命名为可供性(affordance)(Gibson,1986,p.127),在生态学语境中,它就是生态学实体,它所作用的对象是动物的行为而不是身体。二是这种关联和互补可以特异化(be specific to)为能量的运动构型、形态、结构等等,也就是说,这里的构型由于是关联和互补关系的特异化(specificity),在生态心理学的语境中是特示(specification),因而能量运动构型对于被能量(而非运动构型)所作用的动物来说就是潜在的可使用、可获得的(available)信息。第一维度和第二维度合在一起意味着环境中存在着一些独立于动物身体(生理)特异性的信息,这些信息是一种客观形成的信息,它在心理学中属于Gibson 生态心理学的独创。三,环境中存在着客观信息,并且它们标识着动物身体和环境之间交互的可能——行为,因而,对它们的拾取(pickup)就是心理学研究或感知觉研究的主要内容(Gibson,1986,p.239)。补充一句,这里信息拾取当然也可以通过认知科学中的“具身认知”来理解。
在Gibson 之后,生态学方法逐渐地融入自然科学之中,相互作用原理也获得了精确的内涵。Warren 的登台阶知觉判断实验最为经典而简洁地将可供性知觉应用于定量研究中(Warren,1984)。Warren 探讨了生态学方法与传统方法之间的差别,即生态学方法使用内在单位进行测量(intrinsic measurement),并提出了身体尺度(body-scaled)和行为尺度(action-scaled)(Warren,1984,p.686),两者之间的差别不在本文内容之内,因而不作进一步论述。Warren 的实验揭示出,当台阶高度除以被试的腿长之后,高和矮两组实验者的知觉判断值出现一致,这说明环境的身体比性质与知觉判断之间存在内在的关联,即知觉判断范畴与行为的临界值即最高值、最优值存在一致关系。
身体尺度或行为尺度是对Gibson 交互作用原理的定量性的表达。它们说明了,动物直接生存的环境是在尺度交互意义上的物理环境。身体比是环境和身体交互作用而形成的本征或内在(intrinsic)尺度,它可以应用到心理学的研究中,是生态学方法的实质内涵。更详细地说,可供性是由环境的动力(重量、应力、摩擦力等)性质和动物的动力(身体的重量、身体的结构、四肢)联结而成,这种交互性质限定了动物行为的界限(行为阈限),展示了动物行为所发生的本征环境——或本征时空。“每个动物——至少在某些程度上——都是感知者和行为者,用老式词汇说,它是有感知能力和有生命力的。它是环境的感知者,并且在环境中行动。但这并不是说它感知物理世界并在物理的空间和时间中行动。”(Gibson,1986,p.8)
测量尺度和本征尺度之间的差别和内在关联,是生态学的重要问题,也是Gibson 生态心理学中生态学方法的本质内涵,它关系到人类认识自然的工具、手段和方法,以及因此而决定的认识界限和认识发展的规律,同时也是解决认识论问题的重中之重,但是这一问题在认识论中还不是特别凸显。“总体来讲,尺度的存在根源于地球表层自然界的等级组织和复杂性。尺度本质上是自然界所固有的特征或规律,而为有机体所感知。因而尺度又可分为测量尺度和本征(Intrinsic)尺度。测量尺度是用来测量过程和格局的,是人类的一种感知尺度,随感知能力的发展而不断发展。本征尺度是自然现象固有而独立于人类控制之外的。测量尺度相当于研究手段,隶属方法论范畴,而本征尺度则是研究的对象。尺度研究的根本目的在于通过适宜的测量尺度来揭示和把握本征尺度中的规律性。”(吕一河,傅伯杰,2001,第2097 页)
二、特示关系与生态信息
生态学第二个研究维度是信息理论,它的核心概念是特示(specify)。生态系统中充斥着多种形式的能量,如光能、机械能、内能、化学能、原子能、生物能等,它们是物质的固有属性,由于物质本身的属性而具有一些确定的量度,是引发物质运动变化的原因。但另一方面,能量所引起的运动变化必然展现为特定的样式、构型或结构,它们反过来特示(specify 或be specific to,下文简称为特示)物质所具有的属性(Gibson,1960,p.701)。也就是说,能量具有两种意义,第一是在它们的转换中遵循量度上的守恒,是物质之间相互作用所具有的量度,第二则作为信息的载体,在这个意义上,它并不关系到物理运动形式上的变化和转换。按Chemero 所说,“信息不是环境中物体或物质以外某种可测量、可计量的东西,相反,信息是环境的某种关系特征。……光中的信息就是光和环境之间的关系。”(Chemero,2009,p.108)
因而,从对特示关系的分析中,可以发现,能量运动构型和物质属性之间的关联,从属于动力学研究范畴,也就是说,信息是动力学的,它的展示是动态的,它与实在之间的关联是无法通过测量尺度还原的,关于这一点,Gibson 以后,Runeson 首先明确指出生态学信息的动力学机制(Runeson,1983,p.587-588)。信息载体的运动既然是动力学的,那么就意味着,信息是物理的、客观的、实在的,“如果某机体接受器中能量流的不变量存在,如果这些不变量对应于环境的不变特性,并且如果它们是机体对环境知觉的基础,而不是感觉信号的基础,那么,认识论中的实在论就有新的证据,同样,心理学中也会出现新的知觉理论”(Gibson,1982,p.374);同时也说明,信息的存在独立于知觉者,或者说它是非感受器和神经特异性的,“信息这个词语在本书中的意义(不是在其他的书中)是观察者的环境的呈现,而不是观察者的接收器或感觉器官的呈现。物体的性质通过信息展现出来;接收器和神经的性质通过感觉展现出来。关于世界的信息与感觉的性质没有什么关系。”(Gibson,1986,p.242)
特异性(specificity)是一个神经生理学概念,它说的是,感觉、神经兴奋和能量刺激作用之间的关系。“在将外部世界的信息传给个体的过程中,感觉系统执行一系列普遍的功能。其中最基础的是,每个系统都与某个刺激的特异性(specificity)发生反应,它们使用特异化的细胞——外周接收器(peripheral receptors)——把刺激转换为所有神经元都能够使用的信号。”(Squire,Berg,& Bloom,etc.,2009,第535 页)在认知科学中,神经特异性概念稳固地树立了外部世界的“表征”对于认知研究的基础性作用。比如说,认知科学家们将认知划分为感受和解释这两个机制。例如他们认为,“感觉是对刺激的探测,并识别出某件事情的发生;而知觉则是解释和理解。”(Squire,Berg,& Bloom,etc.,2009,第536页)
从这两个特异性关系的比较,可以发现,两者在测量方法上存在差异。生态学方法强调身体尺度或行为尺度对于动物行为研究的基础性作用,因而不变量并不是非量化的东西。在Gibson 以后,生态信息理论开始走向量化的发展方向。以生态光学为例,Gibson 认为生态光学的核心假设是光列阵的变量能够承载关于作为光来源的环境的信息。这意味着,光列阵不是复制实在,而是特示实在。Gibson的学生Lee 致力于将基于生态光学的视觉理论数学化,将其应用在位移行为的视觉控制研究上。在《光流域:视觉的基础》一文中,Lee 提出光流域中包含有光学变量τ(t),它能够表示为在某个时刻t,距离与速度之间的比率,即,Z(t)/V(t)。在这里,光学变量τ(t)是一种光学变化率(即扩张率),它能够作为动物为保持与表面特定关系的光学信息量(Lee,1980)。以驾驶行为为例,在时刻t,某辆瞬时速度为V(t)的汽车驶向一个与其距离为Z(t)的表面,为了防止冲撞到这个表面上,驾驶员必须减速(D)刹车,使得该辆汽车在无限接近表面的时候停止,因而,根据位移公式,V(t)2/2D≤Z(t)。通过对Z(t)/V(t)=τ(t)公式进行微分,得到Z(t)D/ V(t)2 =1-dτ(t)/dt,最后得到公式:
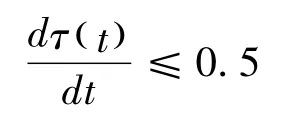
按照这个公式,在与表面接近的过程中,动物与表面之间的接触时间(time-to-contact)就包含在光的运动变量中,它提供了如何与表面保持特定关系的信息,例如,如若避免被碰撞,就需要将τ(t)的时间导数保持为小于等于0.5。对于这个公式的验证,请参看Lee 对蜂鸟飞行的研究(Lee,1991)。光学τ(t)理论提出之后,它被拓展到接触时间的声音变量研究中(Shaw,McGowan,& Turvey,1991)。
三、特示的形式化及生态信息理论的发展
如果能量运动构型是物理事实的特示或特异化,那么就意味着,环绕着动物的能量环境就必然特示动物身体尺度或行为尺度的相关信息,这种信息本质上是行为暗示的,而且它们是准确的清晰的。但另一方面,Gibson 也提出了能量样式对事实特示的局限,这种局限产生了模糊信息。例如光不能特示透明物的结构,前庭系统中耳石的运动样式能够特示启动、停止、推拉、主动和被动加速,即能够特示不稳定状态,但不能够特示静止不动和直线匀速运动状态之间的区别,而半规管的运动样式也不能够特示静止状态和匀速旋转之间的区别,等等。更为明确地说,所谓特示,即能量列阵的样式和事实之间的专属指向关系,但在某些临界条件下会出现一对多,或多对一的关系,这时就产生了模糊信息。模糊信息源于任何能量列阵的运动构型并没有表现出足够的异质性,从而导致动物在拾取信息的时候无法判断事实。
Stoffregen 等人在姿态平衡的知觉研究领域,对特示关系理论进行了完善,在生态信息理论的发展中作用巨大。就身体姿态的平衡来说,首先需要考虑到身体所处环境的性质,即使在静止的状态下身体也存在振荡性的摆动,因而它是一个动力系统,这意味着动物在感知身体方向的时候,面对的是一个动态的信息环境;其次,对于动态信息环境的感知涉入到自身的动力学性质,因而,这里就涉及非线性动力感知,具体地说,在判断方向的时候,不同的感知系统、感知系统与运动系统、身体系统与环境之间的耦合,方向知觉与这种嵌套系统之间的耦合关系是一种复杂的非线性知觉。
因而,Stoffregen 和Riccio 发现了姿态平衡系统所存在的知觉复杂性问题,对基于前庭系统掌管重力感知的定向理论进行了批评(Stoffregen & Riccio,1988)。根据经典的定向理论,身体的方位信息主要基于重力惯性向量,即公式为,Fs=Fg +Fa。也就是说,当身体方向与重力方向发生偏离的时候,身体会跌落,这时候重力的方向就为身体方向提供了基准。然而,有一些情况会引发重力错觉,即,当视觉、声音、触觉等惯性参照系与重力方向产生冲突的时候,会引发重力错觉;当身体主动运动产生加速度的时候,其惯性力向量则与重力产生偏离,前庭系统也不能区分惯性力和重力。这时候的方向判断就需要在前庭系统感知基础上,进行归纳推理,即与其他方向信息对比处理,完成正确的方向判断。
在Stoffregen 和Riccio 看来,根据生态学法,方向知觉必须考虑到支撑面的性质,因为它影响到支撑面对重力的抵抗以及身体为了保持某种姿态所必须对该支撑面所主动施加的作用。在水中,水的浮力分布全身而抵消了质量中心和支点之间的力臂,身体不会因为和重力方向的偏离而产生加速度,因而前庭系统无法判断垂直方向;在彻底失重情况下,就没有重力惯性向量,因而,也就不需要保持特定的姿势了;在有支撑面的情况下,如果支撑面和重力方向产生偏斜角度,质量中心和身体和支撑面之间的支点(即脚)就会产生旋转运动,而为了控制身姿保持平衡,动物必须对支撑面主动施以驱力来抵抗这种重力惯性向量,这时候,就会产生出一种不断向前的加速度,即身体稳定的方向开始与重力方向发生偏移。
Stoffregen 和Riccio 认为,这种不稳定的平衡状态才是运动中身体垂直方向的信息来源。“定向行为涉及到对一种特定运动学状态的获得,即,不稳定的平衡(unstable equilibrium)。因此我们相信,这种不稳定的平衡方向才是相关于姿势控制的垂直状态”(Stoffregen & Riccio,1988,p.8)。也就是说,身体方位的信息既包含重力惯性向量,也需要考虑到身体的运动状态以及考虑到支撑面和身体之间的关系。这种信息就是“模间不变量(intermodal invariants)”信息,它需要知觉系统的统筹才能获得,即,它不是专属地指向特定模态的感受器的信息(例如前庭系统对垂直方向的感知),然后经神经系统对其他模态感受器接受的信息进行整合处理的产物,它的形成完全是一个身体与环境相互作用的事件,即一个生态学事件。
继提出“模间不变量”概念之后,Stoffregen 和Bardy(2001,p.199)又对特示类型进行了逻辑性地分类。他们认为,从能量列阵和物理事实之间的对应关系上来看,包含有3 种类型的特示:1.“无特示(No Specification)”;2.“单模态特示(Modal Specification)”;3.“独立特示(Independent Specification)”,包括“多模态特示(Multiple Specification)”和“变模态特示(Amodal Specification)”。
“无特示”,即能量列阵和实在之间的对应关系是多对多,即,对于某个确定的能量列阵,也可以对应多个实在;对于确定的实在,可以对应某能量列阵的不同形式、结构。在无特示关系中,感觉是模糊的,感觉本身不能作为信息,只能作为信息处理的材料;“单模态特示”关系则主要是Gibson 生态学方法中的特示观点,即单个的能量列阵和实在之间是一一对应关系,例如光环境和光列阵之间的确定对应关系,振动事件和声波之间的对应关系,等等;“独立特示”主张,实在和能量列阵之间的对应关系是一对多,即实在引发相互独立的多种模态列阵以及能量列阵的不同方面。其中的“多样特示”主张,实在某方面影响某特定列阵的多重结构,这些多重结构之间互不相关地特示实在的变化。
Stoffregen 和Bardy 认为,在动物-环境的相互关系中,只有多种模态能量列阵之间生成的高阶列阵即“全模态列阵”才能够清晰地指向、特示一个确定的事件(Stoffregen & Bardy,2001,p.206)。“全模态列阵”揭示出生态系统中能量之间的相互关系,以及这种相互关系对生态事件的特示。任何一事件的变化,都会影响到多种列阵的样式变化,这种惟一的指向关系要求在实在和运动构型之间只能存在惟一的运动构型,因而,无论是单模态特示,还是独立特示,都没有考虑到实在特示的唯一性,因而存在逻辑上的欠缺。全模态列阵不是专属于某特定模态感知系统的信息,而是专属于知觉系统的信息,它的提出是为了规避某特定知觉系统得到信息以后再与其他系统信息进行处理的内在机制问题;它也能够更好地说明为什么有机体要主动行为来探索信息,为什么行为能够更好地支持知觉的实施。例如,被试在判断物体是否在接触范围内的时候,在实验室控制头部静止条件下判断的准确性,和在头部自主运动条件下判断的准确性之间存在差异,后者的准确性具有统计学效度(Mantel,Stoffregen,&Bardy,2008)。
四、特示关系的认识论意蕴
Gibson 的生态心理学被视为具身认知的一个理论来源。本文的研究对象则是为具身认知奠定基础的生态学信息,它是具身认知研究的前奏。理由如下:具身认知的理论前提是实在论的,而不能是表征主义的,即,认识对象必须是实在的、客观的,或者说,认识论的前提必须是“物理主义”的,原因是,如果认识的对象是“心理的”,那么认识本身就是无意义的语义循环。然而,物理主义认识论的危险性在于,在认识一种具有“主观的”、“价值的”属性的研究对象的时候,测量尺度是无法派上用场的,它必须取消“心理的”所标识的属性。最后像行为主义一样,将所有的行为都视为是机械的过程。
因而,在这个意义上,Gibson 的生态学方法给认识论指出了一条有价值的方向。其中的特示关系说明,环境中存在着准确的信息,如果将它应用到动物和环境的交互系统研究中,就会发现,环境中存在着的信息并不是刺激式的,也不是解释的,而是身体尺度或行为尺度的。这说明,信息对于认识的作用是行为引导性的,感觉系统与运动系统的特异性是自然演化的,起源于适应环境选择的功能性要求,而不是结构性的或机制性的。更进一步说,特示关系将知觉、经验等内在意向性的认识论研究方法转变为外在的信息探索问题,在本征尺度的时空中来研究知觉,因而具有将心理学引入动力学研究的可能。但Gibson 没有进一步将其形式化和数学化。因而在他之后,这些问题成为生态心理学的奠基性工作。由于特示理论是生态信息理论的根基,因而Stoffregen 等人“全模态列阵”的研究进一步丰富了特示关系的内涵和外延。
Squire,L.,Berg D.,& Bloom,F.,etc.(2009).基础神经学系列3:感觉和运动系统.北京:科学出版社.
吕一河,傅伯杰.(2001).生态学中的尺度及尺度转换方法.生态学报,21(12).
Chemero,A.(2009).Radical embodied cognitive science.Cambridge,MA:the Massachusetts Institute of Technology Press.
Gibson,J.J.(1960).The Concetp of The Stimulus in Psychology.The American Psychologist.
Gibson,J.J.(1982).New Reasons for Realism.in E.Reed & R.Jones(eds),Reasons for Realism:Selected Essays of James J.Gibson.Hillsdale,New Jersey:Lawrence Erlbaum Associates,Publishers.
Gibson,J.J.(1986).The ecological approach to visual perception.Hillsdale,New Jersey:Lawrence Erlbaum Associates.
Lee,D.N.(1980).The Optic Flow Field:The Foundation of Vision.Phil.Trans.R.Soc.Lond.B290(1038).
Lee,D.N.,Reddish,P.E.,& Rand,D.T.(1991).Aerial Docking by Hummingbirds.Naturwissenschaften 78.
Lombardo,T.J.(1987).The reciprocity of perceiver and the environment.Mahwah,NJ:Lawrence Erlbaum Associates,Inc.P3.
Mantel,B.,Stoffregen,Thomas A.,& Bardy,B.G.(2008).Multisensory perception of whether an object is within reach.Journal of Vision 5.
Runeson,S.,Frykholm,G.(1983).Kinematic Specification of Dynamics as an Informational Basis for Person-and-Action Perception:Expectation,Gender Recognition,and Deceptive Intention.Journal of Experimental Psychology:General,112(5).
Shaw,B.K.,McGowan,R.S.,& Turvey,M.T.(1991).An Acoustic Variable Specifying Time-to-Contact.Ecological Psychology,3(3).
Stoffregen,T.A.,Riccio,G.E.(1988).An Ecological Theory of Orientation and the Vestibular System.Psychological Reviewo,95(1).
Stoffregen,T.A.,Bardy,B.G.(2001).On specification and the senses.Behavioral and Brain Sciences,24(2).
Warren,W.H.(1984).Perceiving affordances:visual guidance of stair climbing.Journal of experimental psychology:Human perception and performance,10(5).
