三丁酸甘油酯和甘露寡糖对菊黄东方鲀生长性能、体组成及肠道健康指标的影响
2014-12-20翟秋玲张春晓孙云章
翟秋玲 张春晓* 孙云章 王 玲 宋 凯 张 璐
(1.厦门市饲料检测与安全评价重点实验室,集美大学水产学院,厦门 361021;2.通威股份有限公司,成都 610041)
肠道健康是鱼类快速生长和鱼体保持健康的重要保障。其中,肠道菌群的组成和数量是评价肠道健康的重要指标,其在促进宿主的生长和营养物质的消化吸收、机体免疫防御、防止消化道疾病发生、保证肠道功能正常以及促进肠细胞增殖等方面起着重要作用。鱼类肠道正常菌群的维持和优化主要依赖于肠道优势菌群种类和数量的变化[1-5]。另外,肠道黏膜形态结构是鱼类消化吸收的结构基础,肠黏膜上皮绒毛高度和宽度的增加,即吸收面积的增加,能够增强其对营养物质的吸收能力[6]。三丁酸甘油酯(tributyrin,TB),化学式为C15H26O6,为无色近油状液体,几乎没有气味或略有脂肪香味。TB在肠道脂肪酶的作用下,分解成丁酸、丁酸甘油一酯和甘油,主要有效成分为丁酸,其作为短链脂肪酸(SCFA),能够为肠道上皮细胞提供能量,促进其生长发育,改善肠道的形态结构[7-8]。研究表明,饲粮中添加TB可以提高断奶仔猪[9]和肉鸡的生长性能[10],改善肉鸡的肠道形态结构和微生物菌群,提高饲料养分消化率[10]。目前,对TB在畜禽动物上的应用有少量研究报道,而在水产动物上的应用则鲜见报道。甘露寡糖(mannan-oligosaccharides,MOS)是典型的功能性寡糖之一,饲料中添加适量的MOS能够提高水产动物的生长性能、非特异性免疫力,改善肠道形态结构及肠道菌群[11-15]。已有研究发现,丁酸钠和MOS的复合添加可使断奶仔猪的生长性能显著优于单独添加丁酸钠或MOS[16]。然而,尚未见动物饲料中TB和MOS复合添加的研究报道。
菊黄东方鲀(Takifugu flavindus),属鲀形目,鲀科,东方鲀属,俗称河鲀,为我国特有的经济东方鲀类,主要分布于我国黄海和东海,为暖温性鱼类[17-18],具有很高的经济价值。近年来,菊黄东方鲀的养殖产业发展迅速,而在高密度集约化养殖条件下,菊黄东方鲀生长缓慢、病害频发,随之而来的药物滥用严重威胁着水环境和食品安全。因而,绿色饲料添加剂的开发和使用备受关注。本试验以菊黄东方鲀为研究对象,通过在饲料中添加不同水平的TB和MOS,考察其对鱼体生长性能、体组成及肠道健康指标的影响,探讨TB与MOS的作用机制和适宜的添加量,为其在水产饲料中的合理应用提供理论依据。
1 材料与方法
1.1 试验饲料
以鱼粉和豆粕为主要蛋白质源,鱼油、豆油和卵磷脂为脂肪源,并补充矿物质、维生素等配制出基础饲料,其组成及营养水平见表1。以基础饲料作为对照(D1组),分别在基础饲料(干物质基础)中添加50 mg/kg TB(购自上海阿拉丁化学有限公司,纯度为98%;D2组)、100 mg/kg TB(D3组)、150 mg/kg TB(D4组)、50 mg/kg TB+250 mg/kg MOS(购自成都格拉西亚化学科技有限公司,纯度为93%;D5组)、100 mg/kg TB+250 mg/kg MOS(D6组),配制成5种试验饲料。将所有固体原料粉碎,过80目筛,按比例混匀后添加油脂并再次混匀,然后加去离子水揉成面团,经双螺杆制粒机(CD4×1TS型,华南理工大学)加工成 1.5 mm×5.0 mm的颗粒,阴干后保存于-20℃冰箱中备用。
1.2 试验鱼来源与驯化
试验用菊黄东方鲀为福建省漳浦慧丰养殖场当年人工孵化的同一批鱼苗,正式试验前,试验鱼在集美大学海水试验场循环过滤桶(1 200 L)中暂养14 d,并以基础饲料饱食投喂(08:30、16:30各投喂1次),使之逐渐适应试验饲料和养殖环境。
1.3 试验分组与饲养管理
试验鱼驯养结束后,停食24 h,挑选出体格健壮、规格一致[平均体重为(15.99±0.15)g]的试验鱼,随机分为6组,每组设3个重复,每个重复20尾,以重复为单位放养于玻璃缸(120 L)内。试验在海水循环过滤系统中进行,每天饱食投喂2次(08:30、16:30 各投喂 1 次),在摄食 0.5 h 后吸去残饵和粪便,并换水50%。试验期间,水温为(27±2)℃,盐度为(30±1)‰,溶解氧浓度>7.6 mg/L。试验期为8周。
1.4 样品采集
饲养试验结束后停食24 h,用丁香酚麻醉,对各组试验鱼进行称重并计数。每缸随机捞取3尾鱼,尾静脉取血,血液放在4℃冰箱内静置12 h后,离心(3 500 r/min,10 min)取血清,置于-80℃冰箱中保存用于分析生化指标;之后,解剖取出内脏,分离肝胰脏,分别称量内脏总重和肝胰脏重;并取出全部肠道和肝胰脏,剔除脂肪,用冷却的生理盐水(质量分数为0.86%)冲洗干净并用滤纸吸干,液氮中保存用于分析肠道消化酶活力及肝胰脏组成。每缸另随机捞取9尾鱼,其中3尾用于肠道菌群培养计数,3尾用于肠道组织切片观察,3尾用于全鱼组成分析。
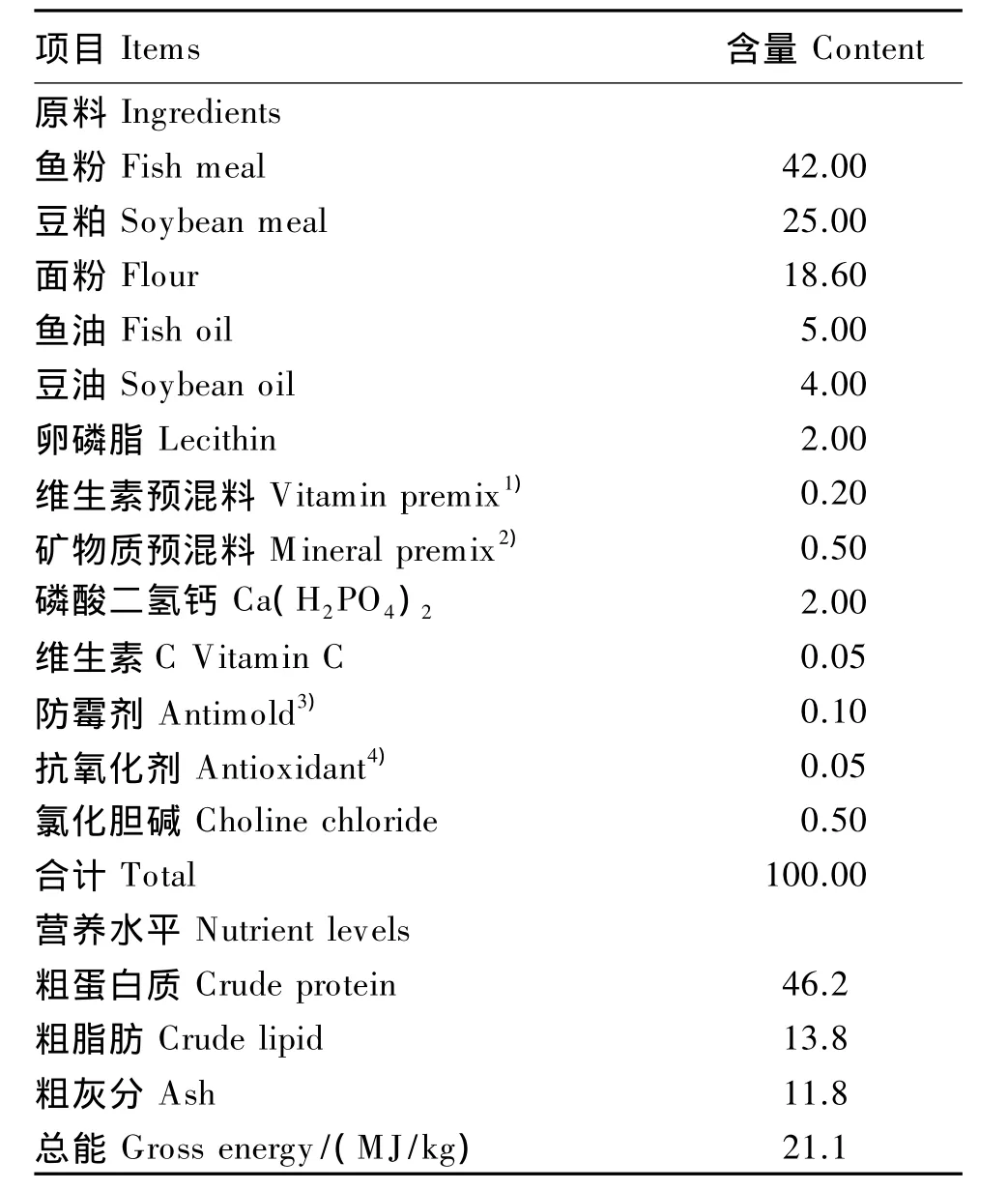
表1 基础饲料组成及营养水平(干物质基础)Table 1 Composition and nutrient levels of the basal diet(DM basis) %
1.5 指标分析与测定
1.5.1 饲料、全鱼和肝胰腺的组成分析
饲料、全鱼和肝胰腺的常规成分测定采用AOAC(2001)[19]的方法。其中,水分含量是在105℃烘箱中烘至恒重后测定,粗蛋白质含量采用凯氏定氮法,使用全自动凯氏定氮仪(FOSS Kjeltec 8400,美国)测定,粗脂肪含量采用索氏抽提法(乙醚为溶剂)测定,粗灰分含量是在马福炉中550℃灼烧12 h后测定。肝糖原含量采用南京建成生物工程研究所生产的试剂盒测定,操作步骤按照说明书进行。
1.5.2 肠道消化酶活力的测定
称量全肠重量,按1∶9(质量体积比)加入0~4℃的生理盐水,在组织匀浆器中匀浆(冰水浴)。用低温离心机2 000 r/min离心10 min后,取出上清液于4℃冰箱中保存备用,24 h内测定完毕。淀粉酶和脂肪酶的活力均采用南京建成生物工程研究所生产的相应试剂盒测定,操作步骤按照说明书进行;组织蛋白质含量采用微量考马斯亮蓝法(试剂盒购自南京建成生物工程研究所)测定;蛋白酶的活力采用福林-酚法[20]测定。淀粉酶活力单位定义:组织中每毫克蛋白质在37℃条件下与底物作用30 min,水解10 mg淀粉定义为1个酶活力单位。脂肪酶活力单位定义:在37℃条件下,每克组织蛋白质在反应体系中与底物反应1 min,每消耗1μmol底物为1个酶活力单位。蛋白酶活力单位定义:在37℃、pH为7.5的条件下,与1%酪蛋白底物作用10 min,每分钟内分解出1μg酪氨酸的酶量为1个酶活力单位。
1.5.3 血清生化指标的测定
血清中碱性磷酸酶(AKP)、谷草转氨酶(GOT)、谷丙转氨酶(GPT)、溶菌酶(LYS)和总超氧化物歧化酶(T-SOD)的活力以及总蛋白(TP)含量均采用南京建成生物工程研究所生产的相应试剂盒测定;血清中甘油三酯(TG)和总胆固醇(TC)含量采用浙江东瓯诊断产品有限公司生产的相应试剂盒测定;血清中葡萄糖(GLU)含量采用上海荣盛生物药业有限公司生产的试剂盒测定。
1.5.4 肠道菌群培养计数
样品采集参考王琨等[21]提供的方法略作改进,具体如下:清水冲洗鱼体表面,拭干,再用酒精棉球擦拭。用灭菌的解剖工具解剖试验鱼,取出肠道,用磷酸盐缓冲溶液(PBS,pH 7.3)冲洗肠道并去除内容物,保留肠壁样品,置于经称重的1.5 mL离心管内,称重,并以1∶4的质量体积比加入无菌PBS,用灭菌匀浆器充分匀浆,制成肠道原液,然后进行梯度稀释。取 10-1、10-2、10-3梯度稀释液各100μL,分别涂布在选择性培养基MRS培养基(购自青岛日水生物技术有限公司)上,用于乳酸菌的分离计数;取10-2、10-3、10-4梯度稀释液各100μL,分别涂布在硫代硫酸盐柠檬酸盐胆盐蔗糖(TCBS)琼脂培养基(购自北京陆桥技术有限责任公司)上,用于弧菌的分离计数;取10-2、10-3、10-4梯度稀释液各100μL,分别涂布在2216培养基(购自青岛海博生物技术有限公司)上,用于需氧异养细菌的分离计数。将上述培养基置于28℃恒温生化培养箱(PYX-250S-B,科力仪器)中培养。选择菌落生长疏密适当、菌落清晰的平板进行计数,并将结果换算成每克样品中菌落数的对数值[lg(CFU/g)][22]进行统计。1.5.5 肠道组织切片观察
用PBS冲洗肠道后,分别取前肠的中间部分和后肠的中间部分各 0.5 cm,Bouin氏液固定12 h,乙醇脱水,二甲苯透明,石蜡包埋,切片(厚度为5μm),苏木精-伊红(HE)染色。切片采用光学显微镜(奥林巴斯BX80,日本)观察、拍照,每张切片选择5个视野。在200倍下测量肠绒毛高度与宽度,在400倍下测量肠上皮细胞高度和微绒毛估计高度,测量方法参考 Escaffre等[23]的方法,并用Motic照相处理软件处理。
1.6 计算公式

式中:W0为初始体重(g);Wt为终末体重(g);t为试验天数(d);WP为摄入的蛋白质总量(g);Wf为摄入的饲料总量(g,干物质基础);Wh为样品鱼肝胰脏重(g);Wv为样品鱼内脏重(g);Wd为样品鱼体重(g)。
1.7 统计与分析
各组数据均采用SPSS 17.0统计软件进行方差分析,若差异显著,则进行Tukey氏多重比较,差异显著水平为P<0.05。所有数据均以平均值±标准误(mean±SE)表示。
2 结果
2.1 TB和MOS对菊黄东方鲀生长性能的影响
养殖试验结束后,各组试验鱼成活率均为100%。由表2可知,菊黄东方鲀的增重率、特定生长率、饲料效率和蛋白质效率随着TB添加量的增加先升高,在添加量为100 mg/kg时达到最大值(分别为 74.72%、1.00%/d、0.53 和 1.14),而后趋于平稳。与单独添加50 mg/kg TB相比,饲料中添加50 mg/kg TB+250 mg/kg MOS显著提高菊黄东方鲀的增重率、特定生长率、饲料效率和蛋白质效率(P<0.05);饲料中添加 100 mg/kg TB+250 mg/kg MOS与单独添加100 mg/kg TB相比,菊黄东方鲀的各生长性能指标均无显著变化(P>0.05)。饲料中单独添加TB有促进肝胰指数和脏体指数增大的趋势,当添加量为150 mg/kg时菊黄东方鲀的肝体指数和脏体指数显著高于对照组(P<0.05),但与2个联合添加组差异不显著(P>0.05)。此外,50 mg/kg TB+250 mg/kg MOS 组的增重率显著高于100 mg/kg TB+250 mg/kg MOS组(P<0.05),其他生长性能指标2组间差异不显著(P>0.05)。
2.2 TB和 MOS对菊黄东方鲀鱼体和肝胰腺组成的影响
由表3可知,饲料中添加TB和MOS对鱼体水分、粗灰分及粗蛋白质以及肝胰腺粗蛋白质、粗灰分含量均无显著影响(P>0.05)。鱼体粗脂肪含量以150 mg/kg TB组最高,显著高于其他各组(P<0.05);同时,联合添加组的鱼体粗脂肪含量要高于对应添加量的单独添加组,且在TB添加量为50 mg/kg时组间差异达到显著水平(P<0.05)。肝胰腺粗脂肪含量以150 mg/kg TB组最高,显著高于除50 mg/kg TB+250 mg/kg MOS组外的其他各组(P<0.05);同时,联合添加组的肝胰腺粗脂肪含量均显著高于对应添加量的单独添加组(P<0.05)。150 mg/kg TB组的肝胰腺水分含量显著低于其他单独添加组(P<0.05),与2个联合添加组差异不显著(P>0.05);同时,联合添加组的肝胰腺含量要低于对应添加量的单独添加组,且在TB添加量为50 mg/kg时组间差异达到显著水平(P<0.05)。肝糖原的含量随着TB添加量的增加而显著下降(P<0.05),联合添加组显著低于对应添加量的单独添加组(P<0.05),以 100 mg/kg TB+250 mg/kg MOS组的肝糖原含量最低。

表2 TB和MOS对菊黄东方鲀生长性能的影响Table 2 Effects of TB and MOS on growth performance of tawny puffer(Takifugu flavindus)
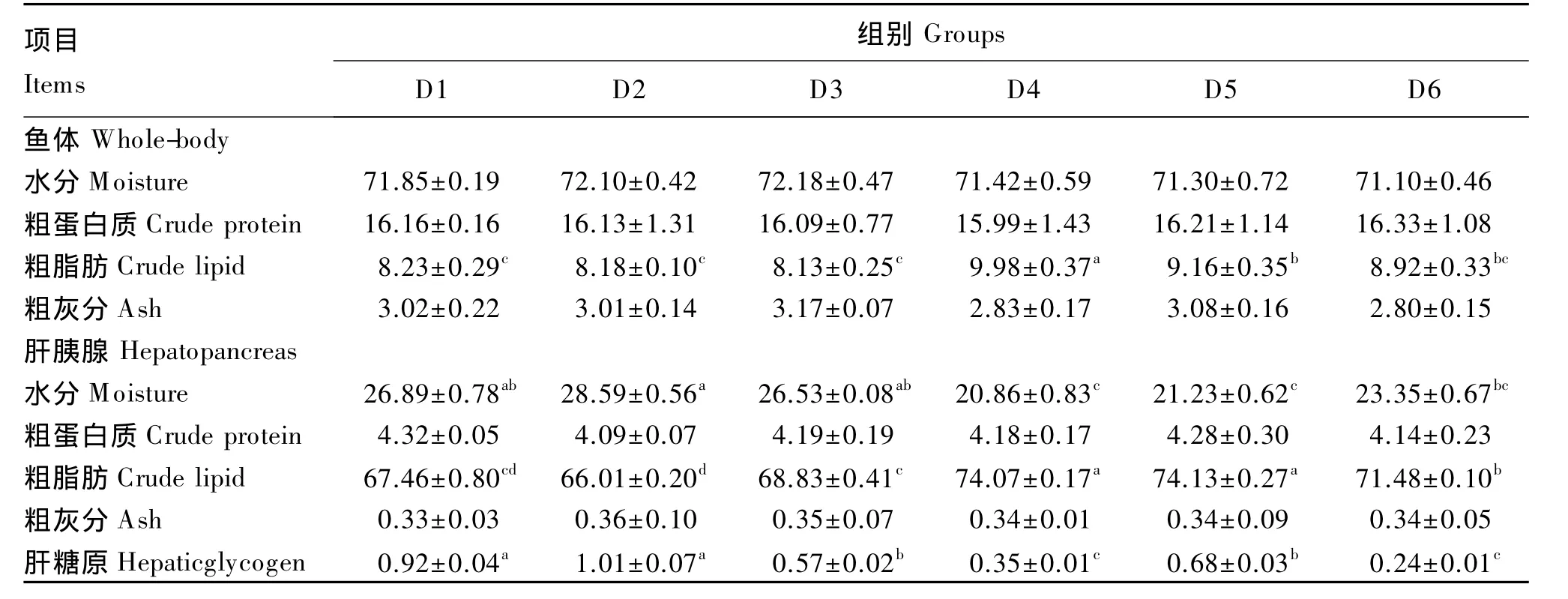
表3 TB和MOS对菊黄东方鲀鱼体和肝胰腺组成的影响(湿重基础)Table 3 Effects of TB and MOS on whole-body and hepatopancreas composition of tawny puffer(Takifugu flavindus)(wet matter basis) %
2.3 TB和MOS对菊黄东方鲀肠道消化酶活力的影响
由表4可知,随着TB添加量的增加,肠道蛋白酶、脂肪酶和淀粉酶活力均先升高后下降,均在100 mg/kg TB组达到最高值。50 mg/kg TB+250 mg/kg MOS组的肠道蛋白酶活力与50 mg/kg TB 组相比显著升高(P<0.05),但肠道脂肪酶和淀粉酶活力2组间差异不显著(P>0.05)。100 mg/kg TB+250 mg/kg MOS 组的肠道脂肪酶活力与100 mg/kg TB组相比显著升高(P<0.05),但其肠道蛋白酶和淀粉酶活力却显著下降(P<0.05)。
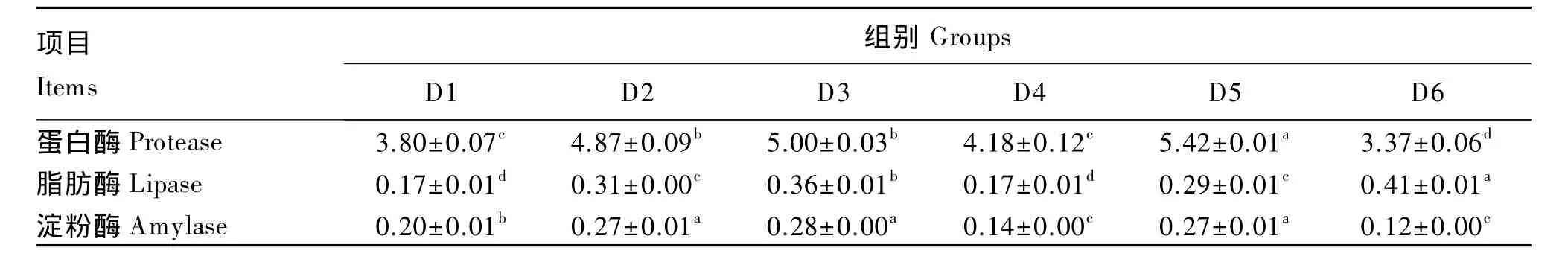
表4 TB和MOS对菊黄东方鲀肠道消化酶活力的影响Table 4 Effects of TB and MOS on intestinal digestive activities of tawny puffer(Takifugu flavindus) U/mg prot
2.4 TB和MOS对菊黄东方鲀血清生化指标的影响
由表5可知,随着TB添加量的增加,菊黄东方鲀血清中总胆固醇含量显著降低(P<0.05),甘油三酯含量有下降的趋势(P>0.05);联合添加组血清总胆固醇和甘油三酯含量均低于对应添加量的单独添加组,但差异不显著(P>0.05)。血清总蛋白和葡萄糖含量随着TB添加量的增加而显著升高(P<0.05);50 mg/kg TB+250 mg/kg MOS 组与50mg/kg TB组相比,血清总蛋白和葡萄糖含量均 显 著 升 高 (P<0.05);100 mg/kg TB+250 mg/kg MOS组与100 mg/kg TB组相比,血清总蛋白含量显著升高(P<0.05),而血清葡萄糖含量则无显著差异(P>0.05)。血清谷草转氨酶和谷丙转氨酶活力随着TB添加量的增加而显著降低(P<0.05),且联合添加组显著低于对应添加量的单独添加组(P<0.05)。血清碱性磷酸酶、溶菌酶以及总超氧化物歧化酶活力在各组间均无显著差异(P>0.05)。

表5 TB和MOS对菊黄东方鲀血清生化指标的影响Table 5 Effects of TB and MOS on serum biochemical indices of tawny puffer(Takifugu flavindus)
2.5 TB和MOS对菊黄东方鲀肠道菌群的影响
由表6可知,随着TB添加量的增加,肠道需氧异养细菌数量先增加后降低,在100 mg/kg TB组显著高于除50 mg/kg TB+250 mg/kg MOS组外的其他各组(P<0.05);同时,50 mg/kg TB+250 mg/kg MOS组与50 mg/kg TB组相比显著增加(P<0.05),100 mg/kg TB+250 mg/kg MOS 组与100 mg/kg TB组相比则显著下降(P<0.05)。50 mg/kg TB+250 mg/kg MOS组的肠道弧菌数量显著高于对照组(P<0.05),但各单独添加组之间无显著差异(P>0.05),联合添加组与对应添加量的单独添加组之间亦无显著差异(P>0.05)。肠道乳酸菌数量先随着TB添加量的增加显著升高(P<0.05),在添加量为 50 mg/kg之后趋于稳定(P>0.05),联合添加组与对应添加量的单独添加组之间无显著差异(P>0.05)。

表6 TB和MOS对菊黄东方鲀肠道菌群的影响Table 6 Effects of TB and MOS on intestinal microflora of tawny puffer(Takifugu flavindus) lg(CFU/g)
2.6 TB和MOS对菊黄东方鲀肠道形态结构的影响
由表7可知,菊黄东方鲀前肠和后肠的绒毛高度和宽度均随着TB添加量的增加而增加,在150 mg/kg TB组达到最大值,其中前肠的绒毛高度显著高于其他单独添加组(P<0.05),前肠的绒毛宽度显著高于对照组(P<0.05),后肠的绒毛高度和宽度显著高于对照组和50 mg/kg TB组(P<0.05)。联合添加组的前肠和后肠的绒毛高度和宽度均高于对应添加量的单独添加组,但二者间差异未达显著水平(P>0.05)。前肠和后肠的上皮细胞高度和微绒毛估计高度各组间均无显著差异(P>0.05)。

表7 TB和MOS对菊黄东方鲀肠道形态结构的影响Table 7 Effects of TB and MOS on intestinal morphology and structure of tawny puffer(Takifugu flavindus) μm
3 讨论
3.1 TB和MOS对菊黄东方鲀生长性能的影响
本试验结果表明,饲料中添加TB对菊黄东方鲀的生长具有促进作用。Hou等[9]和胡杰[24]研究发现,饲粮中添加TB显著提高了断奶仔猪的平均日增重,并降低了料肉比。彭丽莎等[10]在对肉鸡的研究中也获得了相似的结果。此外,本试验结果还表明,与单独添加TB相比,同时添加TB和MOS菊黄东方鲀的生长性能得到进一步提高。研究表明,饲料中单独添加MOS能提高欧洲鲈鱼(Dicentrarchus labrax)[25]、 鲤 鱼 (Cyprinus carpio)[26]、虹鳟(Oncorhynchus mykiss)[27]和红罗非鱼[28]的增重率,降低饲料系数。游金明等[16]研究发现,饲粮中添加0.1%丁酸钠+0.1%MOS组断奶仔猪的生长性能显著优于单独添加丁酸钠或MOS组,而本试验中 50 mg/kg TB+250 mg/kg MOS组菊黄东方鲀表现出最好的生长性能,说明饲料中丁酸与MOS具有协同促生长作用,而二者的实际摩尔比例可能影响受试养殖动物的生长效果。
本试验中,随着TB添加量的增加,肠道蛋白酶、脂肪酶和淀粉酶活力均先升高后下降,其总的变化趋势与饲料效率、增重率和特定生长率相似,其中100 mg/kg TB组试验鱼肠道中3种消化酶的活力均最高,且生长性能最好。这说明适量的TB可能通过影响消化酶活力,促进饲料营养物质的消化吸收,从而改善菊黄东方鲀的生长性能,这与其他学者对丁酸钠(主要有效成分为丁酸)在断奶仔猪上的研究结果[7]相似。TB在肠道内被脂肪酶分解成丁酸、丁酸甘油一酯和甘油[29]。脂肪酸盐可通过刺激胆囊收缩素释放而促进胰腺分泌消化酶类[30],从而提高肠道消化酶活力。而150 mg/kg TB组肠道蛋白酶、脂肪酶和淀粉酶活力均显著低于100 mg/kg TB组,100 mg/kg TB+250 mg/kg MOS组肠道蛋白酶和淀粉酶活力显著低于50 mg/kg TB+250 mg/kg MOS组,这可能是由于饲料TB添加过量或一定量的TB与MOS协同对消化酶活力产生负面影响,其具体机理还有待进一步研究。
3.2 TB和 MOS对菊黄东方鲀鱼体和肝胰腺组成的影响
肝胰腺是营养物质代谢的主要器官,因此肝体指数的大小与动物代谢活力有关[31-32]。本试验发现,饲料中单独添加TB或联合添加TB和MOS后,试验鱼鱼体和肝胰腺中粗脂肪含量增加,而血清中总胆固醇和甘油三酯含量降低,说明TB和MOS的添加促进了脂质在鱼体和肝胰腺中沉积。此外,试验鱼的肝体指数和脏体指数随着TB添加量的增加而升高,与鱼体和肝胰腺中粗脂肪含量的变化趋势一致,而肝胰腺是菊黄东方鲀脂肪沉积的主要器官,也是其最大的内脏器官,因此,菊黄东方鲀的肝体指数和脏体指数也能间接反映肝胰腺的脂肪水平。
本试验中,随着饲料中TB添加量的增加,肝糖原的含量呈下降趋势,而血清中葡萄糖含量则呈升高趋势。这说明TB可促进菊黄东方鲀肝糖原分解为葡萄糖进入血液,为机体新陈代谢供能。而细胞利用葡萄糖的主要限速步骤是葡萄糖的跨膜转运,这一过程是依靠细胞膜上的葡萄糖运载体(glucose transporter,GT)来完成的,而丁酸能够促进转运蛋白基因的表达,这已被Takano等[33]证实,因此丁酸可促进动物体内糖的分解代谢。根据鱼体组成和血清总胆固醇与甘油三酯含量数据,作者认为饲料中添加TB和MOS可能促进菊黄东方鲀对碳水化合物的优先利用而节约脂质。另外,与50 mg/kg TB+250 mg/kg MOS组相比,100 mg/kg TB+250 mg/kg MOS组试验鱼肝胰腺粗脂肪和血清葡萄糖含量显著降低,这可能由于该组试验鱼的肠道淀粉酶活力较低,影响其对碳水化合物的吸收,导致其肝糖原和血清葡萄糖含量降低,从而动员机体脂肪分解以维持肝糖原和血糖水平。
3.3 TB和MOS对菊黄东方鲀血清生化指标的影响
血清转氨酶活力是评价动物肝胰腺健康程度的指标[34]。本试验中,饲料中添加TB和MOS降低了试验鱼血清中谷草转氨酶和谷丙转氨酶活力,说明TB和MOS有利于改善菊黄东方鲀肝胰腺的健康。血清中总蛋白含量在一定程度上反映了机体的蛋白质代谢状况。已有研究表明,饲料中添加MOS可提高草鱼(Ctenopharyngodon idellus)血清中总蛋白含量[35]。在本试验中,饲料中单独添加TB或联合添加TB和MOS均显著提高了试验鱼的血清总蛋白含量,这说明饲料中TB和MOS的添加促进了菊黄东方鲀蛋白质的沉积。另外,本试验中肠道蛋白酶活力与血清总蛋白含量的变化并不一致,这可能是因为从肠道消化吸收的氨基酸并不是合成蛋白质的氨基酸的唯一来源,因此肠道蛋白酶活力是鱼体内蛋白质合成水平的充分不必要条件。
3.4 TB和MOS对菊黄东方鲀肠道菌群和形态结构的影响
动物肠道菌群和形态结构是评价动物肠道健康的重要指标。本试验中,饲料中单独添加TB或联合添加TB和MOS均提高了试验鱼肠道中需氧异养细菌和乳酸菌数量,并降低了弧菌数量。彭丽莎等[10]研究发现,在肉鸡饲粮中添加TB可显著增加其盲肠乳酸菌数量,显著降低大肠杆菌数量。Gálfi等[36]和 Castillo 等[37]在断奶仔猪的饲粮中添加丁酸钠也得到相似的结果。这说明丁酸能够选择性地抑制弧菌等有害菌的增殖,促进乳酸菌等有益菌的增殖[38-39]。MOS在肠道能被乳酸杆菌等有益菌特异性利用而促进有益菌群大量增殖,有益菌竞争性占据上皮黏膜结合位点,有效地阻断了弧菌等有害菌群的定植[25,40-41]。另外,MOS也可以直接与病原菌细胞表面外源凝集素有效结合,从而抑制病原菌与肠道上皮的结合,有利于有益菌的增殖[42]。然而,与单独添加TB相比,联合添加TB和MOS并没有显著影响肠道各菌群的数量,说明二者对菊黄东方鲀肠道菌群的影响不存在交互作用。
本试验结果表明,饲料中单独添加TB或联合添加TB和MOS增加了菊黄东方鲀前肠和后肠的绒毛高度和宽度,有利于肠道吸收面积的增加和营养物质消化利用率的提高[43]。已有研究表明,饲料中添加丁酸钠或 MOS可增加肉仔鸡[10,44-45]、断奶仔猪[46]、异带重牙鲷(Diplodus sargus)[47]及军曹鱼(Rachycent roncanadum)的肠绒毛高度和微绒毛密度,使肠绒毛更加整齐完整[41]。丁酸是肠上皮细胞的快速能量和黏膜的营养因子,可促进肠上皮的增殖,修复受损黏膜上皮[48-49];而MOS可能通过促进肠道乳酸杆菌等有益菌的增殖,改善肠道内环境,从而有利于肠绒毛的发育生长。本试验中,TB和MOS联合添加组前肠和后肠的绒毛高度和宽度与对应添加量的TB单独添加组相比有增加的趋势,说明二者在改善菊黄东方鲀肠道形态结构上可能存在协同促进作用。
4 结 论
饲料中添加50 mg/kg TB+250 mg/kg MOS可改善菊黄东方鲀肠道菌群和形态结构,维护肠道健康,增强肠道消化酶活力,促进碳水化合物的利用而节约脂质,提高饲料效率和生长性能。
[1] 尾崎久雄.鱼类消化生理(下)[M].上海:上海科学技术出版社,1983:675.
[2] CAHILL M M.Bacterial flora of fishes:a review[J].Microbial Ecology,1990,19(1):21-41.
[3] NAYAK S K.Role of gastrointestinal microbiota in fish[J].Aquaculture Research,2010,41(11):1553-1573.
[4] RAWLS JF,SAMUEL B S,GORDON JI.Gnotobiotic zebrafish reveal evolutionarily conserved responses to the gut microbiota[J].Proceedings of the National Academy of Sciences of the United States of America,2004,101(13):4596-4601.
[5] SAXTON K E,RAWLS W J,ROMBERGER J S,et al.Estimating generalized soil-water characteristics from texture[J].Soil Science Society of America Journal,1986,50(4):1031-1036.
[6] 刘爱君,冷向军,李小勤,等.艾佐迈对奥尼罗非鱼生长、肠道结构和非特异性免疫的影响[J].动物营养学报,2009,21(6):1006-1011.
[7] 钟翔,黄小国,陈莎莎,等.丁酸钠对断奶仔猪生长性能和肠道消化酶活性的影响[J].动物营养学报,2009,21(5):719-726.
[8] 王继凤,陈耀星,王子旭,等.丁酸钠对断奶仔猪小肠黏膜形态结构的影响[J].中国兽医科技,2005,35(4):298-301.
[9] HOU Y Q,LIU Y L,HU J,et al.Effects of lactitol and tributyrin on growth performance,small intestinal morphology and enzyme activity in weaned pigs[J].Asian Australasian Journal of Animal Sciences,2006,19(10):1470-1477.
[10] 彭丽莎,孙健栋,史艳云,等.三丁酸甘油酯对肉鸡生长性能、养分表观消化率、屠宰性能、肠道形态及微生物菌群的影响[J].动物营养学报,2014,26(2):466-473.
[11] 刘爱君,冷向军,李小勤,等.甘露寡糖对奥尼罗非鱼(Oreochromis niloticus×O.aureus)生长、肠道结构和非特异性免疫的影响[J].浙江大学学报:农业与生命科学版,2009,35(3):329-336.
[12] 王锐,刘军,刘辉宇,等.半乳甘露寡糖对异育银鲫幼鱼生长和非特异性免疫的影响[J].上海水产大学学报,2008,17(4):502-506.
[13] DIMITROGLOU A,MERRIFIELD D L,SPRING P,et al.Effects of mannan oligosaccharide(MOS)supplementation on growth performance,feed utilisation,intestinal histology and gut microbiota of gilthead sea bream(Sparus aurata)[J].Aquaculture,2010,300(1/2/3/4):182-188.
[14] YILMAZ E,GENC M A,GENC E.Effects of dietary mannan oligosaccharides on growth,body composition,and intestine and liver histology of rainbow trout,Oncorhynchus mykiss[J].Israeli Journal of Aquaculture:Bamidgeh,2007,59(3):182-188.
[15] SANG H M,FOTEDAR R.Prebiotic mannan oligosaccharide diet improves health status of the digestive system of marron,Cherax tenuimanus(Smith 1912)[J].Journal of Applied Aquaculture,2010,22(3):240-250.
[16] 游金明,付建福,王自蕊,等.丁酸钠和甘露寡糖对断奶仔猪生长性能和免疫功能的影响[J].动物营养学报,2010,22(2):346-351.
[17] 范文涛,刘海金,赵文江,等.菊黄东方鲀♀×红鳍东方鲀 杂交后代早期形态特征及生长速度的比较[J].水产学报,2011,35(7):1065-1071.
[18] KIM S S,LEE K J.Dietary protein requirement of juvenile tiger puffer(Takifugu rubripes)[J].Aquaculture,2009,287(1/2):219-222.
[19] AOAC.Official methods of analysis of official analytical chemists international[S].15th ed.Arlington,VA:Association of Official Analytical Chemists,2001.
[20] 中山大学生物系生化微生物教研室.生化技术导论[M].北京:科学出版社,1979:46-56.
[21] 王琨,孙云章,李富东,等.饲料中添加两种寡糖和一种芽孢杆菌对牙鲆肠道菌群的影响[J].大连海洋大学学报,2011,26(4):299-305.
[22] ZHANG J,LIU Y,TIAN L,et al.Effects of dietary mannan oligosaccharide on growth performance,gut morphology and stress tolerance of juvenile Pacific white shrimp,Litopenaeus vannamei[J].Fish & Shellfish Immunology,2012,33(4):1027-1032.
[23] ESCAFFRE A M,KAUSHIK S,MAMBRINI M.Morphometric evaluation of changes in the digestive tract of rainbow trout(Oncorhynchus mykiss)due to fish meal replacement with soy protein concentrate[J].Aquaculture,2007,273(1):127-138.
[24] 胡杰.几种物质对早期断奶仔猪生长性能,肠道结构与功能及血液指标的影响[D].硕士学位论文.武汉:华中农业大学,2004.
[25] TORRECILLAS S,MAKOL A,CABALLERO M J,et al.Immune stimulation and improved infection resistance in European sea bass(Dicentrarchus labrax)fed mannan oligosaccharides[J].Fish & Shellfish Immunology,2007,23(5):969-981.
[26] STAYKOV Y,DENEV S,SPRING P.Influence of dietary mannan oligosaccharides(Bio-Mos)on growth rate and immune function of common carp(Cyprinus carpio L.)[M]//HOWELL B,FLOS R.Lessons from the past to optimise the future.[S.l.]:European Aquaculture Society,2005:431-432.
[27] STAYKOV Y,SPRING P,DENEV S,et al.Effect of a mannan oligosaccharide on the growth performance and immune status of rainbow trout(Oncorhynchus mykiss)[J].Aquaculture International,2007,15(2):153-161.
[28] HANLEY F,BROWN H,CARBERRY J.First observations on the effects of mannan oligosaccharide added to the hatchery diets for warmwater hybrid red tilapia[C]//Nutritional biotechnology in the feed industry:proceedings of Alltech’s 11th annual symposium.Nottinghan:Nottinghan University Press,1995.
[29] PIVA A,PRANDINI A,FIORENTINI L,et al.Tributyrin and lactitol synergistically enhanced the trophic status of the intestinal mucosa and reduced histamine levels in the gut of nursery pigs[J].Journal of Animal Science,2002,80(3):670-680.
[30] 艾晓杰,韩正康.禽胰液分泌的调节[J].国外畜牧科技,2000,27(5):37-39.
[31] ROBAINA L,IZQUIERDO M S,MOYANO F J,et al.Soybean and lupin seed meals as protein sources in diets for gilthead seabream(Sparus aurata):nutritional and histological implications[J].Aquaculture,1995,130(2/3):219-233.
[32] KROGDAHL Å,SUNDBY A,OLLI J J.Atlantic salmon(Salmo salar)and rainbow trout(Oncorhynchus mykiss)digest and metabolize nutrients differently.Effects of water salinity and dietary starch level[J].Aquaculture,2004,229(1/2/3/4):335-360.
[33] TAKANO M,RHOADS D B,ISSELBACHER K J.Sodium butyrate increases glucose transporter expression in LLC-PK1 cells[J].Proceedings of the National Academy of Sciences of the United States of America,1988,85(21):8072-8075.
[34] 伍淳操,王建华.乳酸菌和甘露寡糖对断奶仔猪生长及血清生化指标的影响[J].江苏农业学报,2011,27(1):94-99.
[35] 陈希,吴志新,刘佳佳,等.甘露寡糖对草鱼血清生化特性的影响[J].南昌大学学报:理科版,2011,35(4):393-397.
[36] GÁLFI P,BOKORI J.Feeding trial in pigs with a diet containing sodium n-butyrate[J].Acta Veterinaria Hungarica,1989,38(1/2):3-17.
[37] CASTILLO M,MARTÍN-ORÙE S M,ROCA M,et al.The response of gastrointestinal microbiota to avilamycin,butyrate,and plant extracts in early-weaned pigs[J].Journal of Animal Science,2006,84(10):2725-2734.
[38] CANIBE N,STEIEN S H,OVERLAND M,et al.Effect of K-diformate in starter diets on acidity,microbiota,and the amount of organic acids in the digestive tract of piglets,and on gastric alterations[J].Journal of Animal Science,2001,79(8):2123-2133.
[39] DANIELSON A D,PEO E R,Jr,SHAHANI K M,et al.Anticholesteremic property of Lactobacillus acidophilus yogurt fed to mature boars[J].Journal of An-imal Science,1989,67(4):966-974.
[40] GRISDALE-HELLAND B,HELLAND S J,GATLINⅢ D M.The effects of dietary supplementation with mannanoligosaccharide,fructooligosaccharide or galactooligosaccharide on the growth and feed utilization of Atlantic salmon(Salmo salar)[J].Aquaculture,2008,283(1/2/3/4):163-167.
[41] SALZE G,MCLEAN E,SCHWARZ M H,et al.Dietary mannan oligosaccharide enhances salinity tolerance and gut development of larval cobia[J].Aquaculture,2008,274(1):148-152.
[42] FULLER R,HOUGHTON S B,BROOKER B E.Attachment of Streptococcus faecium to the duodenal epithelium of the chicken and its importance in colonization of the small intestine[J].Applied and Environmental Microbiology,1981,41(6):1433-1441.
[43] CASPARY W F.Physiology and pathophysiology of intestinal absorption[J].The American Journal of Clinical Nutrition,1992,55(Suppl.1):299-308.
[44] 胡忠宏.短链脂肪酸对肉仔鸡小肠功能的影响及有关作用机制[D].博士学位论文.北京:中国农业大学,2006.
[45] HU Z H,GUO Y M.Effects of dietary sodium butyrate supplementation on the intestinal morphological structure,absorptive function and gut flora in chickens[J].Animal Feed Science and Technology,2007,132(3):240-249.
[46] LU JJ,ZOU X T,WANG Y M.Effects of sodium butyrate on the growth performance,intestinal microflora and morphology of weanling pigs[J].Journal of Animal and Feed Sciences,2008,17(4):568-578.
[47] DIMITROGLOU A,DAVIES S,DIVANACH P,et al.The role of mannan oligosaccharide in gut development of white sea bream,Diplodus sargus[C]//Nutritional biotechnology in the feed industry:proceedings of Alltech’s 21st annual symposium.Lexington,KY:[s.n.],2005.
[48] KRIPKE S A,FOX A D,BERMAN JM,et al.Stimulation of intestinal mucosal growth with intracolonic infusion of short-chain fatty acids[J].Journal of Parenteral and Enteral Nutrition,1989,13(2):109-116.
[49] ROMBEAU J L,REILLY K J,ROLANDELLI R H.Short-chain fatty acids and intestinal surgery:rationale and clinical implication[M]//CUMMINGS J H ,ROMBEAU J L,SAKATA T.Physiological and clinical aspects of short-chain fatty acids.Cambridge:Cambridge University Press,1995:401-425.
